Существует много веществ биологического происхожденияt представляющих интерес с научной или практической точки зрения, которые трудно отнести к одному из рассмотренных в предыдущих разделах классов соединений. Эти вещества имеют смешанное строени и сформированы из остатков липидов, сахаров и аминокислот в различных сочетаниях. Как указано в табл. 2.11, соединения смешанного строения выполняют рял. важных биологических функций. В следующем разделе мы более детально обсудим химию и строение наружных оболочек различных клеток; здесь встречаются смешанные соединения, а также специфические полисахариды и белки.
Таблица 2.11. Некоторые примеры биохимически важных соединений смешанного строения,
их основных элементов структуры и биологических функций

2.5.1. Клеточные стенки; пептидогликаны и липополисахариды
В разделе, посвященном липидам, мы уже говорили о том, что клеточные мембраны играют жизненно важную роль в регуляции транспорта веществ в клетку и из клетки. В этом отношении не менее важны и другие структурные элементы наружных поверхностей клеток микроорганизмов и тканей. С точки зрения условий существования микроорганизмы, напри ер бактерии, живут в гораздо более изменчивом и менее контролируемом окружении, чем клетки тканей животных, например, клетки печени, поэтому микроорганизмы должны обладать гораздо большей жесткостью, устойчивостью к физическим нагрузкам и резким изменениям осмотического давления. Отсюда следует, что по структуре наружные оболочки бактерий до.1жны сильно отличаться от клеток животных.
В процессах биохимической технологии оболочки клеток представляют интерес в нескольких аспектах. Во-первых, свойства наружных поверхностей клеток определяют их способность к адгезии друг с другом, а также со стенками реакторов, трубопроводов и сепараторов. Как мы увидим в последующих главах, такие явления необходимо учитывать при получении иммобилизованных клеточных катализаторов. Они влияют также на работу микробных реакторов непрерывного действия и на процессы отделения клеток от жидкости. Химические и механические характеристики оболочек определяют также устойчивость клетки к воздействию физических, ферментативных и химических факторов, что существенно в процессах выделения компонентов клетки.
Как уже упоминалось выше при обсуждении реакции Грама, структуры оболочек грамположительных и грамотрицательных клеток сильно отличаются друг от друга (рис. 2.24). У тех и у других оболочки состоят из нескольких слоев, но их положение, толщина и состав далеко не идентичны. На этом рисунке не показаны различные белки, находящиеся внутри клеточной мембраны или закрепленные на ее поверхности; их мы рассмотрим ниже, в гл. 5, в ходе изучения транспорта веществ через мембраны.
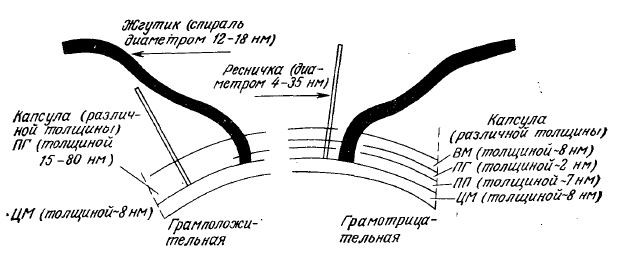
РИС. 2.24. Схематическое изображение структуры стенок грамположительных и грамотрицательных бактерий.
Обозначения: ЦМ – цитоплазматическая мембрана; ПГ – пептидогликан;
ПП – периплазматическое пространство; ВМ – внешняя мембрана.
Грамположительные бактерии имеют одну мембрану, а грамотрицательные клетки – две аналогичные мембраны. Между наружной и цитоплазматической мембранами в rрамотрицательных бактериях находится область, называемая периплазматически и пространством или периплазмой. В периплазматическим пространстве, включающем от 20 до 40 процентов общей массы клетки, находится ряд ферментов, а также белков, связывающих сахара и аминокислоты.
Как в грамположительных, так и в грамотрицательных бактериях непосредственно к внешней поверхности цитоплазматнческой мембраны примыкает пептидогликановый слой. Пептидогликаны построены из остатков дисахарида [состоящего из N-ацетилмурамовой кислоты (NAM) и N-ацетилглюкозамина (NAG), связанных β-1,4-гликозидной связью], мостикового пентапептида, состоящего только из остатков глицина, и тетрапептида. Строение последнего зависит от вида микроорганизма; в Staphylococcus aureus, например, этот тетрапептид содержит остатки L-аланина, D-глутамина, L-лизина и D-аланина (рис. 2.25, а; обратите внимание на наличие редких D-аминокислот). Все перечисленные элементы структуры пептидогликанов связаны множеством поперечных связей, образуя как бы одну гигантскую макромолекулу, окружающую всю клетку (рис. 2.25, 6).
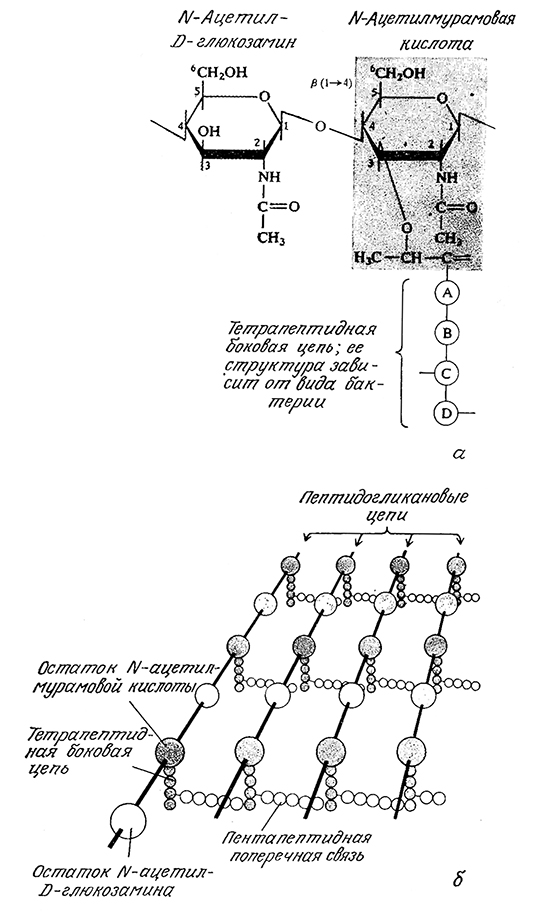
РИС. 2.25. а — строение основных структурных элементов пептидогликанов;
б — схематическое изображение пептидогликана клеточной стенки грамположительной бактерии
Staphylococcus aureus, содержащего большое количество поперечных связей.
Фермент лизоцим, структура которого приведена на рис. 2.17, является эффективным антибактериальным агентом. В основе его бактерицидного действия лежит гидролиз гликозидных β-1,4-связей между остатками NAM и NAG пептидогликана, приводящий к разрушению и удалению пептидогликановой оболочки бактерий и таким путем к разрыву (лизису) клетки в нативных гипотонических растворах. В лабораторных условиях в изотонической среде удается получить живые клетки, не содержащие пептидогликановых оболочек. После обработки лизоцимом в таких условиях образуются клетки, которые теряют свойственные им очертания и принимают сферическую форму; их называют сферопластами. Если известно, что клеточная стенка удалена полностью, то такие клетки называют протопластами.
Важными компонентами внешней части наружных мембран грамотрицательных бактерий являются липополисахариды. Некоторые липополисахариды называют эндотоксинамu, поскольку они в высшей степени токсичны для животных. Токсичность липополисахаридов – одна из причин того, почему заражение крови Е. coli может быть чрезвычайно опасным. По этой же причине при очистке белков, синтезируемых генетически трансформированной Е. coli, необходимо тщательное удаление эндотоксинов.
Молекулы липополисахаридов внешней мембраны имеют три участка: а), липидный компонент А, состоящий из шести ненасыщенных жирных кислот, углеводородные цепи которых проходят в мембрану; они связаны с остатком диглюкозамина; б) центральную) ояигосахаридную область, построенную из остатков десяти моносахаридов; некоторые из них относятся к числу редких Сахаров; в) боковую О-цепь, состоящую из множества повторяющихся тетрасахаридных остатков. Центральная область и О-цепь направлены от клетки в среду. Таким образом, именно наружные боковые О-цепи взаимодействуют с иммунной системой зараженного животного. Путем мутаций бактерии могут достаточно быстро менять структуру О-цепей, что является частью системы их антииммунной защиты.
На этом, однако, рассказ о структуре клеточных стенок бактерий не кончается. У многих видов микроорганизмов внешняя мембрана окружена капсулой или слизистым слоем, по химической природе представляющим собой полисахарид. Капсула одного из штаммов пневмококков (бактерий, вызывающих пневмонию) построена из чередующихся остатков глюкозы и глюкуроновой кислоты. Не имеющие такой полисахаридной капсулы мутанты не обладают патогенными свойствами. Полисахаридная капсула некоторых бактерий может растворяться в среде. Производство внеклеточных полисахаридов является важным промышленным процессом, который, однако, существенно осложняется неньютоновским характером течения жидкой среды (гл. 8). Слизистые слои, кроме того, принимают участие при флокуляции бактерий, являющейся важным этапом процессов обработки сточных вод методом активного ила.
Цитоплазматическая мембрана дрожжевых клеток состоит из липидов, белков и полисахаридов, содержащих остатки маннозы. Если двигаться дальше к внешней оболочке клетки, то за цитоплазматической мембраной будет расположено пери- плазматическое пространство, окруженное в свою очередь клеточной стенкой. В пекарских дрожжах, Saccharomyces cerevi- siae, клеточная стенка содержит от 6 до 8% белков (в том числе несколько ферментов), а также приблизительно по 30% (по массе) глюкана (полисахарида, построенного из остатков d- глюкозы, соединенных β-1,6-связями, а также поперечными β-1,3-связями) и маннана (полиманнозы с а-1,6-связями и а-1,2-боковыми цепями). При обработке 5. cerevisiae ферментом, гидролизующим глюкан, например, 1,3-глюканазой, можно получать соответствующие протопласты. Такую обработку обычно проводят при введении рекомбинантных молекул ДНК в дрожжевые клетки.
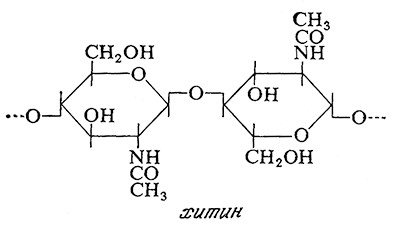
Клеточные стенки дрожжей и многих плесеней содержат хитин, макромолекула которого построена из остатков N-ацетилглюкозамина, соединенных гликозидными β-1,4-связями.
Животные клетки, существующие обычно в строго контролируемом изотоническом окружении, не имеют клеточных стенок. Помимо фосфолипидов и белков в их плазматических мембранах содержится от 2 до 10% углеводов. Последние обнаружены на внешней поверхности всех клеток млекопитающих, изученных до настоящего времени; они связаны с липидами и белками в виде гликолипидов и гликопротеинов соответственно.
2.5.2. Антитела и другие гликопротеины
Белки, содержащие ковалентно связанные остатки моносахаридов или коротких олигосахаридных цепей, называют гликопротеинами. Гликопротеины самых различных типов обнаружены в эукариотах и окружающей их среде. К числу гликопротеинов относится ряд ферментов, например, глюкозооксидаза, продуцируемая Aspergillus niger. Упоминавшийся выше коллаген– биологический опорный элемент – также представляет собой гликозилированный белок. Гликопротеинами являются и некоторые интерфероны, мощные противовирусные агенты. По сути дела, большая часть белков эукариот, контактирующих с окружением клетки или выделяемых ею в среду, представляет собой гликопротеины. Некоторые гликопротеины уже стали или, по всей вероятности, скоро станут ценными промышленными продуктами. Биосинтез гликопротеинов и их участие в транспорте веществ мы рассмотрим в гл. 5 и 6.
К гликопротеинам относятся и антитела – важнейшее оружие иммунологической защиты позвоночных (рис. 2.26). С помощью обсуждаемых в гл. 6 методов слияния клеток можно получать в значительных количествах гомогенные антитела в организмах животных или в биологических реакторах, что позволит в будущем широко применять антитела для диагностических целей, введения лекарственных препаратов и разделения биологически важных веществ. По этой причине нам кажется целесообразным вкратце обсудить здесь происхождение, строение и функции антител.
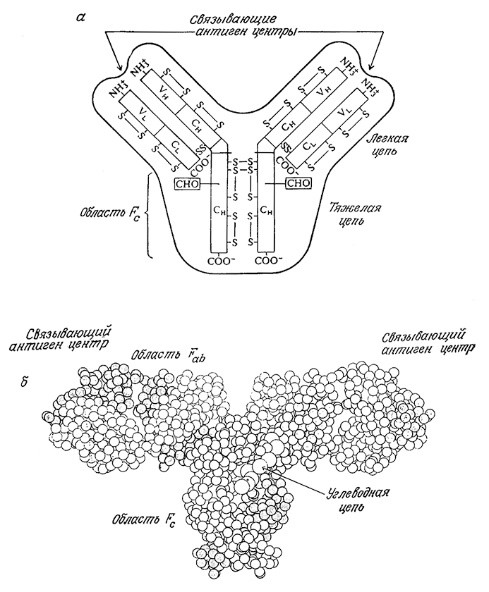
РИС. 2.26. а – схематичное изображение структуры иммуноглобулина.
Символами С и V обозначены постоянные и вариабельные части цепей соответственно,
а индексами Н и L – тяжелые и легкие цепи соответственно. Показаны дисульфидные связи,
а также место присоединения углеводного остатка (СНО) к тяжелым цепям. Область ствола (Fc)
и центр, связывающий антиген, расположены на карбоксильном и аминном концах тяжелых цепей соответственно.
Специфичность связывания антигена и сродство к нему определяются вариабельными частями
как тяжелых, так и легких цепей; б – молекулярная структура антитела IgG, иммуноглобулина G.
В состав иммунной системы позвоночных входят В-клетки, принадлежащие к одному из двух типов обнаруженных в организме лимфоцитов; в присутствии чужеродного вещества, вируса или клетки (антигена) В-клетки дифференцируются и образуют плазматические клетки, которые выделяют антитела. Последние специфично связывают антигены (и близкие по строению вещества); образующийся комплекс антиген-антитело осаждается и выводится из организма.
Антитела представляют собой особый класс белков, называемых иммуноглобулинами-, свойственные иммуноглобулинам общие детали структуры изображены на рис. 2.26. Молекула иммуноглобулина состоит из двух идентичных, более длинных «тяжелых» цепей, связанных друг с другом дисульфидными связями, а нековалентными связями с двумя другими, также идентичными, но более короткими «легкими» цепями. В наиболее широко распространенном классе иммуноглобулинов, обозначаемом IgG, легкие и тяжелые цепи имеют молекулярные массы 23 000 и 53 000 соответственно. С-Концевые участки как легких, так и тяжелых цепей, практически не отличающиеся по структуре в иммуноглобулинах одного типа, называют постоянными участками. Область Fc, представляющая собой ствол молекулы антитела, построена из С-концевых последовательностей постоянных участков тяжелых цепей.
Дифференциация антител в пределах одного типа осуществляется прежде всего в N-концевых участках цепей, называемых вариабельными. Связывающий антиген активный центр антитела (паратоп) формируется из вариабельных участкоз легких и тяжелых цепей. Как и в случае каталитической активности ферментов, связывающие антиген центры в различных антителах могут существенно различаться по своей специфичности и сродству к антигенам.
