Психротрофы - микроорганизмы, которые при температуре 6,5° С и ниже дают рост в благоприятных для их развития средах при инкубации в течение до 10 сут. Из них выделяют психрофилы, оптимальная температура для роста которых - не выше 15° С, максимальная - не выше 20° С. Психрофилы обитают в специфических экологических нишах, где температура не поднимается выше 20° С, например, в глубине морей и океанов; они не играют никакой роли в сыроделии. Собственно психротрофы имеют максимальную температуру для роста выше 20° С, оптимальную - выше 15° С.
Психротрофы широко распространены в природе: в воде, почве, сточных водах. Широкое внедрение холодильного хранения делает психротрофы одной из основных групп микрофлоры сырого молока, что, наряду с уникальными свойствами многих психротрофов, определяет их огромное значение для сыроделия.
На молочные фермы психротрофы попадают главным образом с водой. На большинстве ферм имеется оборудование для охлаждения и хранения охлажденного молока. В охлажденном молоке психротрофы размножаются без особой конкуренции со стороны других сапрофитов и при нарушениях правил гигиены распространяются по всей ферме. По Моисеевой, доля психротрофов в микрофлоре свежевыдоенного молока составила при высоком уровне гигиены на фермах - 10%, при низком - до 75%. Содержание психротрофов в молоке с фермы, где оно не охлаждалось и через 1-2 ч после дойки направлялось на молочный завод, равнялось в среднем 47-103 (7-87 тыс.) КОЕ/мл или 19,6% от общего количества микроорганизмов. В то же время, в молоке, которое на ферме хранили при 9-10° С и дважды в сутки отправляли на завод, их количество составило 1,85-10* (от 1,6 до 2,3106) КОЕ/мл или 45,7% от общего количества бактерий. По неопубликованным данным Перфильева (1998), содержание психротрофов в 31 партии молока, поступившего в 1993-1994 гг. в экспериментальный цех ВНИИМС, составило в среднем 4,8-106 (от 4-103 до 1,2-107 КОЕ/мл). При этом 65,4% образцов содержали менее 103 КОЕ/мл психротрофов. Высокая обсеме- ненность некоторых партий молока психротрофами объясняется низким качеством мойки и дезинфекции оборудования для хранения и транспортировки охлажденного молока, адгезией клеток психротрофов к поверхностям из нержавеющей стали и образованием на них колоний, достаточно высокими температурами хранения молока по сравнению с применяемыми за рубежом. Так, если время генерации Pseudomonas fluorescens в молоке при 4° С равняется 10,5 ч, то при 10° С - 5,5 ч. Эти данные свидетельствуют о том, что главной причиной высокого загрязнения молока психротрофами является размножение в недостаточно охлажденном молоке. По Кузину с соавт., 75% закупленного учебноопытным заводом Вологодского молочного института молока содержало более 106 КОЕ/мл психротрофов. В молоке, поступающем на сыродельный завод ВНИИМС, содержание психротрофов достигало 80% от общего содержания бактерий. Психротрофы обладают низкой редуцирующей активностью и поэтому не полностью выявляются редуктазной пробой. Вызывает удивление большое увеличение содержания психротрофов в молоке во время транспортировки: при среднем увеличении содержания микроорганизмов в молоке за время транспортировки в 3,2 раза, численность психротрофов увеличилась в 21,7 раза. Возможно, это связано с увеличением содержания растворенного в молоке кислорода или с разобщением скоплений психротрофов, в результате чего количество клеток психротрофов, составляющих образующую колонию единицу, уменьшается, а следовательно, увеличивается количество колониеобразующих единиц при постоянном содержании клеток.
Состав психротрофной микрофлоры сырого молока (% от общего числа психротрофов): Pseudomonas 37,1; Acinetobacter - 24,7; Flavo- bacterium - 13,7; Escherichia - 10,7; Micrococcus - 6,0; Enterobacter - 5,0; Aeromonas - 1,8; Klebsiella - 1,0 [1756]. Близкий состав психротрофов молока установлен венгерскими исследователями, но они нашли дополнительно представителей родов Alcaligenes и Serratia.
Психротрофы можно разделить на такие, у которых способность расти при низких температурах является родовым или видовым признаком (Pseudomonas, Acinetobacter, Flavobacterium и др.), и такие, у которых это свойство присуще только части штаммов, например, энтеробактерии. До 75% психротрофов сырого молока принадлежат к Pseudomonas к Acinetobacter. В Японии установлено, что 90,2% культур психротрофов, выделенных из сырого и пастеризованного молока, являются псевдомонадами, в Аргентине доля псевдомонад среди психротрофов сырого молока превысила 50%. Pseudomonas, Acinetobacter, Flavobacterium, составляющие основную часть психротрофов сырого молока, являются грамотрицательными палочками размером, соответственно (0,5—1,0)х(1,5—5,0); (0,9-1,6)х(1,5-2,5) и (0,5-1,0)х(1,0-3,0) мкм. Представители рода Acinetobacter в стационарной фазе развития приобретают сферическую форму. Флавобактерии неподвижны, ацине- тобактеры двигаются рывками, псевдомонады осуществляют плавающее движение с помощью одного или нескольких жгутиков. Строгие аэробы, метаболизм дыхательный с использованием кислорода в качестве конечного акцептора электронов, псевдомонады могут расти в анаэробных условиях с использованием в качестве акцептора электронов нитратов. Ацинетобактеры в питательных средах не образуют пигментов, псевдомонады образуют флюоресцирующие диффундирующие и недиффундирующие пигменты, флавобактерии при росте на питательных средах образуют пигменты от желтого до оранжевого цвета.
Из псевдомонад в молоке чаще всего обнаруживают Р. fluorescens, и, кроме него, Р. putida, Р. fragi, Р. aeruginosa. Они непатогенны для человека, кроме Р. aeruginosa, который является условно- патогенным или даже патогенным микроорганизмом. Р. aeruginosa (синегнойная палочка) образует диффундирующий нефлюоресцирующий пигмент сине-зеленого цвета, Р. fluorescens пигменты этого типа не образует. Р. aeruginosa синтезирует экзо-, эндоэнтеротоксины и может вызывать заболевания по типу пищевых и водных ток- сикоинфекций, но случаи таких заболеваний через сыры в литературе не описаны (сычужные сыры неблагоприятны для роста строгих аэробов). Доля желудочно-кишечных заболеваний, вызываемых синегнойной палочкой, в общем количестве заболеваний ЖКТ, вызванных условно- патогенными микроорганизмами, равняется 2,04%. Р. aeruginosa при хороших гигиенических условиях получения молока редко обнаруживается, однако при неудовлетворительной мойке и дезинфекции оборудования она обнаруживалась в 56% проб в количестве от 10 до 106КОЕ/мл.
Способностью медленно расти при низких температурах обладают L monocytogenes и Y. enterocolitica, но доля этих видов в общем количестве психротрофов сырого молока незначительна.
Оптимальные температуры для роста Р. aeruginosa - 37° С, для остальных видов - 22-26° С; Р. aeruginosa не растет при температуре 4° С и ниже, растет при 41° С; Р. putida и Р. fluorescens растут при 4° С, но не 41° С; Р. fragi не растет при температуре выше 36° С. Оптимальный pH для роста Р. aeruginosa в молоке равен 6,25-6,50; рост этого вида в средах задерживали 5% соли.
В сыроделии условия для преимущественного роста психротрофов создаются при хранении молока и сыра при 2-8° С и созревании молока при 8-12° С. Сведения о скорости развития психротрофов в молоке при этих температурах противоречивы. По одним данным, за 4 сут их количество в молоке увеличилось при 2° С в 3-4 раза, при 4° С - в 6-8 раз, по другим - в 41 или в 100 раз. По данным венгерских ученых, количество психротрофов в молоке за 48 ч хранения при температурах 2, 5 и 8° С возросло в 2; 3,8 и 6,6 раза соответственно.
В табл. 6.25 показано развитие психротрофов в молоке в зависимости от степени его исходного загрязнения микроорганизмами, температуры и продолжительности хранения. В молоке со сравнительно невысокой исходной бактериальной обсемененностью количество психротрофов увеличилось за двое суток при 4-5° С примерно в 13 раз, при 7-8° С - в 500 раз и при 10-12° С - в 800 раз. В молоке с высокой исходной бактериальной загрязненностью скорость размножения психротрофов уменьшилась в 3-7 раз. Тесной корреляции между исходным и конечным содержанием психротрофов в молоке после хранения или созревания нет.
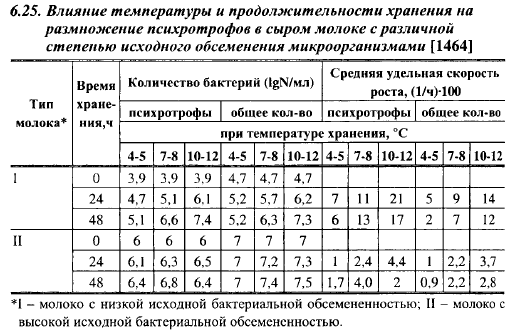
Различия в интенсивности развития психротрофов в молоке, устанавливаемые разными авторами, могут быть также обусловлены неодинаковым составом психротрофной микрофлоры в изучаемых образцах сырого молока. При температурах 4-5° С и ниже в молоке активно размножается только Р. fluorescens, что, очевидно, и делает этот вид наиболее распространенным в сыром молоке, хранившемся при достаточно низких температурах. Чем выше исходная доля этого вида среди психротрофов, тем быстрее будет идти накопление их биомассы.
Как среда для роста психротрофов образцы молока по физико-химическим свойствам могут сильно различаться. Особенно большое значение имеет содержание в молоке растворенного кислорода, поскольку большинство психротрофов являются строгими аэробами.
Снижение интенсивности размножения психротрофов в молоке низкого бактериального качества можно объяснить ингибиторным действием на рост психротрофов остальной микрофлоры. Имеются многочисленные данные об ингибировании роста психротрофов в молоке молочнокислыми бактериями. Кузин изучал совместный рост Р. fluorescens и молочнокислых бактерий в молоке при 9-11° С. Посевные дозы псевдомонад (0,01-1,0)-106 КОЕ/мл, молочнокислых бактерий - (0,01—10)-106 КОЕ/мл. Степень антагонизма количественно он характеризовал антагонистическим индексом (А):
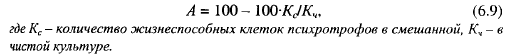
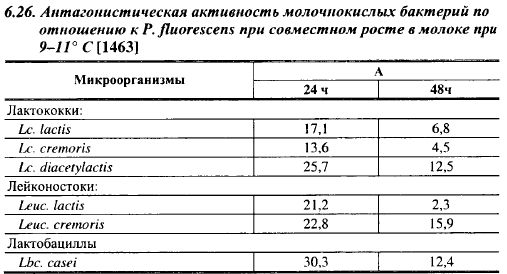
В табл. 6.26 приведены результаты его исследований. Из 112 проверенных штаммов молочнокислых бактерий только 4 не обладали антагонизмом к Р. fluorescens, что свидетельствует о том, что способность подавлять рост этого вида в молоке при 9-11° С является общим свойством молочнокислых бактерий, хотя выраженность антагонистических свойств неодинакова у различных родов, видов и штаммов. Наиболее сильными антагонистами из лактококков оказался подвид Lc. diacetylactis, из лейконостоков - Leuc. cremoris, среди лактобацилл - Lbc. casei. В пределах вида антагонистическая активность была наименее изменчива у Leuc. cremoris и Lbc. casei, штаммы которых по антагонистической активности разнились не более чем на 25%. Антагонистические индексы были в 1,4-9,2 раза выше при совместном росте в первые 24 часа, чем по прошествии 48 ч. Так, если после 24 ч совместного роста с Lbc. casei количество жизнеспособных клеток псевдомонад было ниже на 30,3%, то после 48 ч разница уменьшилась до 12,4%. Важно, что антагонистическое действие на псевдомонады проявили даже виды и штаммы молочнокислых бактерий, которые не размножались при 9-11° С в течение 1-2 сут, в частности Lbc. casei. Американские ученые наблюдали значительное ингибирование роста психротрофов при 5,5° С при внесении в среду 2,5Т08 /мл болгарской палочки, которая не размножается при этой температуре; степень ингибирования коррелировала с количеством образуемой лактобациллами перекиси. Это явление можно объяснить или внесением с инокулятом молочнокислых бактерий специфических антибиотических веществ, адсорбированных на поверхности клеток, или разобщенностью процессов размножения и синтеза антибиотических веществ, подобно разобщенности кислотообразования и размножения у лактококков (гл. 3). Каким бы ни был механизм ингибирования роста психротрофов покоящимися клетками лактобацилл, это явление позволяет использовать молочнокислые бактерии для ограничения роста психротрофов во время хранения или созревания молока без опасения нежелательных для производства сыра изменений физико-химических свойств молока. Добавление в сырое молоко иммобилизованных в частицах Са - альгинатного геля клеток L. lactis и Lbc. helveticus в количестве 2,7-107 и 13-107 клеток/мл, соответственно, снижало в нем содержание психротрофов после 2-суточного хранения при 7° С на 50% при снижении pH только на 0,1-0,22 ед.
Положительные результаты получены при использовании специальных заквасок для созревания молока с целью ингибирования роста психротрофов. Закваски с таким функциональным назначением может поставлять Угличская экспериментальная биофабрика.
Пастеризация молока при 12° С в течение 15 с снижала содержание в нем псевдомонад с 10б клеток/мл до 10 клеток/мл; выдержка этого молока после пастеризации при 7° С в течение 7 сут привела к увеличению содержания псевдомонад до 104 клеток/мл. Таким образом, пастеризация молока по принятому в сыроделии режиму не полностью уничтожает псевдомонады при достаточно высоком их исходном содержании в молоке. В то же время, Mikawa, изучив очень большое количество психротрофных бактерий, выделенных из сырого и пастеризованного молока, пришел к выводу об их уничтожении во время пастеризации молока при 63° С в течение 30 мин. Анаэробные условия в сырах препятствуют активному развитию психротрофов молока, большинство которых являются строгими аэробами, но существует прямая корреляция между содержанием психротрофов в твердых сырах с низкими температурами II нагревания после прессования и количеством психротрофов в смеси для выработки сыра (г = 0,92).
В опытах Перфильева (неопубликованные материалы) психротрофы достаточно активно размножались во время созревания Голландского сыра. Независимо от исходного содержания в смеси их содержание в период максимума составило (2,2—2,7)-108 КОЕ/r. Однако этот максимум наступил в сырах, выработанных при исходном содержании в смеси единиц, тысяч и десятков тысяч КОЕ/мл психротрофов соответственно через 35, 26 и 15 сут созревания. Напрашивается вывод, что в сырах имеется ограниченное количество источников энергии, которые могут быть использованы только психротрофными клетками (в противном случае они были бы использованы к 15-35-суточному возрасту мезофильной микрофлорой сыра). Видовой состав психротрофов сыра Перфильевым не был исследован. По-видимому, в этом случае размножались штаммы факультативных анаэробов, обладающих психротрофными свойствами.
Несколько другие результаты получены литовскими исследователями. По их данным, увеличение содержания психротрофов в сырах с низкой температурой II нагревания наблюдалось только на начальном этапе созревания, после чего оно стало снижаться. Не установлено статистически достоверной связи между содержанием психротрофов в период максимума и в конце созревания. Масштабы размножения психротрофов зависели от активности молочнокислого брожения.
Различия между результатами этих исследований можно объяснить неодинаковым составом психротрофов в сырах, в частности долей в их составе энтеробактерий. Имеется сообщение, что до 25% психротрофов сыра могут быть представлены энтеробактериями.
Вред сыроделию приносят протеолитические и липолитические энзимы, образуемые психротрофами в молоке и сохраняющие активность после пастеризации. Во время роста в молоке протеолитически активные виды и штаммы психротрофов переводят часть белка в растворимое состояние, что снижает выход сыра. Для определения величины снижения выхода сыра из-за размножения психротрофов предложена формула:
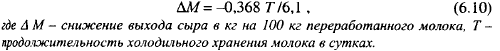
Эта формула дает ориентировочную цифру, так как она не учитывает ряда факторов, влияющих на интенсивность роста и продуцирования протеолитических энзимов психротрофами, в частности температуры хранения.
Протеолитические и липолитические энзимы психротрофов, сохраняющие активность после пастеризации молока, вызывают прогорклый, горький, салистый, фруктовый вкус и некоторые другие привкусы; выраженность этих пороков зависит от количества образовавшихся энзимов, температуры и продолжительности созревания сыров. Фруктовый вкус и запах, например, возникают в результате взаимодействия жирных кислот - продуктов гидролиза молочного жира липазами - и спиртов, образуемых лейконостоками, дрожжами и некоторыми другими микроорганизмами. При высоком содержании в молоке псих- ротрофы ухудшают сычужную свертываемость молока, делают сычужный сгусток менее плотным, плохо отдающим сыворотку, время сычужного свертывания в зависимости от вида психротрофов может снижаться или возрастать. Выдержка молока при низких температурах вызывает изменения мицелл казеина, что также серьезно ухудшает его сычужную свертываемость (гл. 2). Разделить влияние психротрофов и химических изменений молока под воздействием низких температур на его сыропригодность трудно.
В молоке с высоким содержанием психротрофов скорость кислотообразования лактококками повышается, что обусловлено образованием низкомолекулярных продуктов протеолиза. Большинство исследованных штаммов психротрофов стимулировало рост List, monocytogenes.
Протеолитической и липолитической активностями обладали 94% выделенных из молока псевдомонад, в т. ч. 23,4% сильной и 29,4% слабой протеолитической активностью. Среди психротрофов других родов обнаружены слабые протеолиты и липолиты. Wessels et al. нашли, что из 75 психротрофных культур энтеробактерий 7 обладали протеолитической активностью в молоке при 7° С, 10 - при 25° С; липолитической активностью - 9 культур.
Образование внеклеточных иротеиназ и липаз псевдомонадами происходит в конце логарифмической и начале стационарной фаз роста, когда плотность их популяции превысит 105 КОЕ/мл. Достаточно высокие концентрации внеклеточных протеиназ и липаз в молоке обнаруживаются при наличии 106— 107 КОЕ/мл псевдомонад. При длительном культивировании начинается автолиз клеток псевдомонад и в среду выделяются внутриклеточные протеиназы. Очищенные амино- пептидазы из Р. fluorescens расщепляют синтетические «горькие» пептиды (не все) и сохраняют активность в сыре в течение 2 мес.
В культуральных средах Р. fluorescens и Р. aeruginosa максимальное количество протеиназ и липаз образуется при 20 и 30° С соответственно. С понижением температуры содержание этих энзимов в среде уменьшается, а в пересчете на единицу сухой биомассы увеличивается; при температуре 2° С синтез протеиназ и липаз прекращается или происходит с очень низкой скоростью. При 4° С синтез протеиназ и липаз Р. fluorescens на ед. клеточной массы был на 25% выше, чем при 10° С. Липолитическая активность псевдомонад наиболее сильно проявляется при 5° С.
Оптимальный pH для синтеза протеиназ и липаз псевдомонадами лежит в интервале 6,8-7,4; в отсутствие аэрации они начинают синтезировать внеклеточные протеиназы и липазы при более низкой концентрации клеток в среде.
Уникальным свойством протеиназ и липаз психротрофов является необычайно высокая термоустойчивость. Они не инактивируются пастеризацией молока при 77° С в течение 17 с и 140° С в течение 5 с; Дсю°с протеиназы Р. fluorescens равнялось 110 мин. Д90°с протеиназ психротрофов было в 10 раз выше, чем Д-значение спор В. stearothermophilus. Однако 20-минутная выдержка при 55 и 60° С снижает активность протеиназ психротрофов на 47 и 44%, увеличение продолжительности выдержки при этих температурах повышает степень инактивации. При 45° С они инактивируются за 30 мин. Если при 60°С протеиназы психротрофов инактивируются, то при температурах выше 65° С они становятся термоустойчивыми. Липаза Р. fluorescens полностью или частично инактивировалась при 65° С, протеина- за оставалась активной. По Law, липазы псевдомонад полностью или частично сохраняют активность после выдержки при 63° С в течение 30 мин. Инактивацию протеиназ при относительно низких температурах объясняют их автолизом, устойчивость к высоким температурам - конформационными изменениями белка. Возможно, это явление обусловлено разрушением ингибиторов протеиназ при повышенных температурах. Интересно, что в ультрафильтрате молока без казеина липаза Р. fluorescens инактивируется при нагревании до 60 и 65° С, а в ультрафильтрате с казеином сохраняет свою активность. Протеиназы, образуемые различными штаммами и видами псевдомонад, могут обладать различной термоустойчивостью.
Поскольку достаточные количества протеиназ и липаз образуются психротрофами в молоке, когда их количество превысит 106 КОЕ/мл, а пастеризацией молока эти энзимы не инактивируются, можно ожидать, что сыры, выработанные из молока с таким уровнем развития психротрофов, будут низкого качества. В сыре Чеддер, выработанном из молока с наличием более 107 КОЕ/мл психротрофов, количество свободных жирных кислот было в 3-10 раз выше, чем в сырах из молока с низким содержанием психротрофов; в 4-месячных опытных сырах появился прогорклый вкус.
Голландский сыр из молока с 106 КОЕ/мл психротрофов в зрелом возрасте получил оценку на 1 балл ниже, чем контрольный сыр, а опытный сыр из молока с 5106 КОЕ/мл - ниже на 3 балла. Небольшая разница в оценке обусловлена коротким периодом созревания этого сыра (45 сут), во время которого результаты деятельности энзимов психротрофов не успели проявиться в полной мере. Зрелые сыры хранили при 1 и 8° С в течение 3-х мес: после хранения при 1° С качество опытных сыров было ниже контрольных на 1-2 балла, при 8° С - на 3-6 баллов.
Сыры Камамбер, выработанные из молока, хранившегося 9 дней при 5° С, имели горький и прогорклый вкус, не имели аммиачного запаха, содержали больше свободных жирных кислот, чем сыры, выработанные из этого же молока после 2-суточного хранения при 5° С.
Псевдомонады, добавленные в молоко после пастеризации, во время выработки и созревания твердого сыра с низкой температурой II нагревания не размножались и быстро вымирали. Это объясняет отсутствие токсикоинфекций через сыры, вызываемых Р. aeruginosa. Таким образом, основной контроль психротрофов должен проводиться на стадии созревания и резервировании молока. Однако, психротрофные энтеробактерии, составляющие небольшую долю психротрофной микрофлоры сырого молока, размножаются в сырах и могут образовывать протеиназы и липазы непосредственно в сыре. В сырах они вызывают те же пороки, какие вызывают соответствующие энзимы псевдомонад. В твердых сырах энтеробактерии составляют до 21% общего числа психротрофов.
Kielwein показал, что Р. fluorescens является причиной гнилостного запаха Эмментальского сыра, однако нужно отметить ненормальный состав микрофлоры изученных им дефектных сыров.
Псевдомонады были ответственны за появление красно-коричневых пятен и гнилостного запаха на поверхности сыра Моцарелла. Источником загрязнения этого сыра псевдомонадами было не молоко, а вода, применяемая в производстве. Псевдомонады могут вызывать пороки корки и других сыров, в т. ч. твердых сыров с высокими температурами II нагревания. Псевдомонады хорошо размножаются в сыре Коттедж и являются главными виновниками его микробиологической порчи.
Хорошие результаты в предотвращении порчи Домашнего сыра псевдомонадами дает применение биогарда - препарата, приготовленного на основе бактериоцинов молочнокислых бактерий. В опытном сыре с 0,4% биогарда при 7,2° С микрофлора не размножалась в течение 30 дней, большинство контрольных сыров потеряли потребительские качества через 21 день, когда содержание в них псевдомонад достигло 108 КОЕ/г.
Меры предотвращения снижения качества сыров в результате размножения в молоке психротрофов включают:
- улучшение гигиены молока, включая особенно тщательную мойку и дезинфекцию оборудования для хранения охлажденного молока, молокопроводов после каждого производственного цикла;
- хранение молока при температурах ниже 4° С, при которых синтез протеиназ и липаз психротрофами не происходит или идет с очень низкой скоростью;
- термизацию или пастеризацию молока перед хранением и созреванием;
- добавление в молоко, направляемое на созревание или хранение, заквасок, обладающих специфическим антагонизмом по отношению к психротрофам;
- предотвращение вработки воздуха в молоко путем контроля работы насосов, исправности прокладок;
- насыщение молока, находящегося на хранении или созревании, СО2 или азотом для создания анаэробных условий.
