Основные компоненты мясных и молочных продуктов (белок, жир, лактоза) представляют собой благоприятную среду для развития бактерий, плесневых грибов и дрожжей. При комнатной температуре (18—25 °C) мясные и молочные продукты нестойки в хранении, так как в них начинают развиваться различные виды микроорганизмов. Одни виды микроорганизмов, например бактерии родов Pseudomonas, Proteus, Escherichia и все плесневые грибы, вызывают органолептическую видимую порчу — на поверхности продукта появляется слизь, возникает неприятный запах и пр. Присутствие или развитие других видов микроорганизмов не приводит к заметным внешним изменениям продукта. Однако эти микроорганизмы могут явиться возбудителями инфекционных заболеваний (например, бактерии рода Salmonella) или пищевых отравлений (например, бактерии Staphylococcus aureus). Микроорганизмы в продукте развиваются при определенных температурных условиях. По отношению к температуре их делят на три группы: термофилы, мезофилы и психрофилы. Термофилы размножаются при температуре от 20 до 80 °C (оптимум 50—55 °C); мезофилы— от 10 до 45°C (оптимум 25—40°C), психрофилы— от +35 до —10 °C (оптимум 10—20 °C). Верхняя температурная граница роста психрофилов совпадает с оптимальной температурой роста мезофилов, а оптимальная температура развития психрофилов — с нижней температурной границей развития мезофилов (рис. 1). Следовательно, для психрофильных и мезофильных микроорганизмов характерен общий диапазон температур размножения (от 35—37 до 10°C). Эти микроорганизмы различаются минимальными температурами роста.
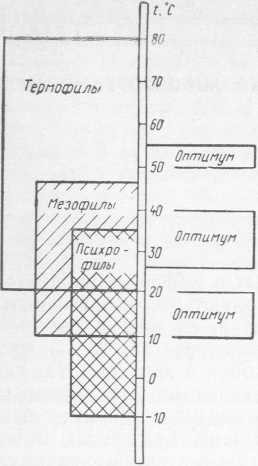
Рис. 1. Температурные границы роста микроорганизмов
При холодильном хранении мясных и молочных продуктов наиболее нежелательно развитие психротрофных микроорганизмов, которые способны размножаться при температуре до —10 °C, а следовательно, и вызывать ухудшение качества продукта. Мезофильные микроорганизмы при температуре развития ниже минимальной могут сохраняться в продукте не размножаясь. Однако происходящие при этом метаболические процессы также отрицательно сказываются на качестве продукта.
Психротрофные микроорганизмы
Микроорганизмы, развивающиеся в пищевых продуктах при холодильном хранении, относятся к факультативным психрофилам. Было предложено называть эти бактерии «психротрофными», т. е. способными питаться и расти на холоду (Eddy, 1960). В 1968 г. по решению Международной молочной федерации принято следующее определение психротрофов и психрофилов. Психротрофы — это микроорганизмы, которые размножаются при 7 °C или ниже, без учета их оптимальной температуры роста. К психрофилам отнесены микроорганизмы, которые также размножаются при 7 °C, но имеют оптимальную температуру роста ниже 20 °C. На 64-й сессии Международной молочной федерации в 1980 г. было сделано уточнение: психротрофными следует считать микроорганизмы, которые вырастают на питательных средах при 6,5 °C в течение 10 дней.
Психротрофные микроорганизмы встречаются среди бактерий, плесневых грибов и дрожжей.
Бактерии. Возбудителями бактериальной порчи мясных и молочных продуктов являются главным образом психротрофные бактерии родов Pseudomonas, Aeromonas. Brochothrix и родов Flavobacterium и Alcaligenes с неясным систематическим положением.
Бактерии рода Pseudomonas представляют собой грамотрицательные аэробные палочки. Клетки одиночные, прямые или изогнутые (но не спиральные) размером 0,5—1X1,5—4 мкм (рис. 2, а). Движутся с помощью моно- или политрихальных полярных жгутиков (рис. 2,6). Метаболизм дыхательный.
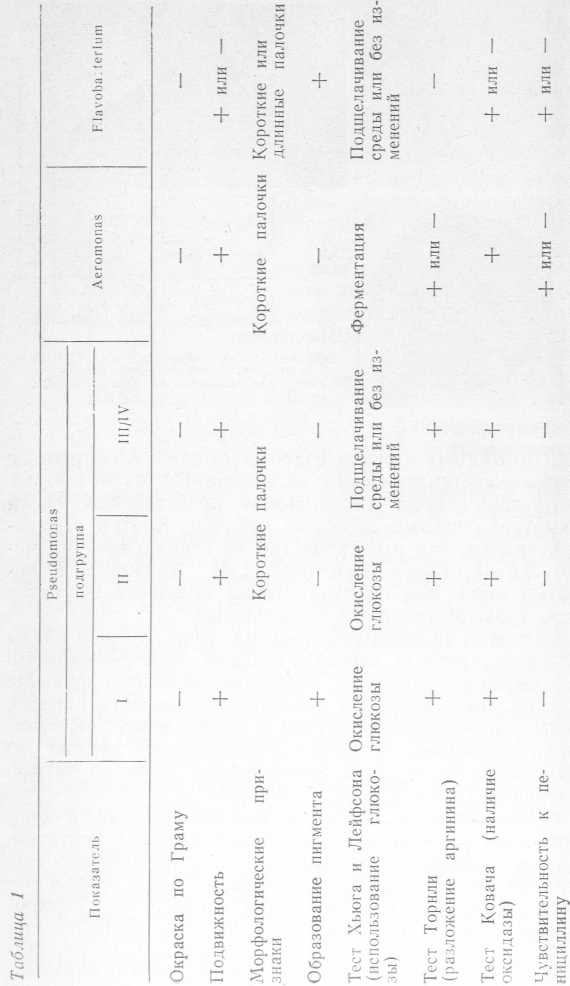
Бактерии на поверхности твердой питательной среды образуют крупные серовато-грязные или зеленовато- желтые полупрозрачные, слизистые, флюоресцирующие колонии (рис. 2,в). По способности изменять окраску среды в результате диффузии в нее пигмента, выделяемого бактериями, а также образовывать кислоту из глюкозы бактерии рода Pseudomonas делят на четыре подгруппы (табл. 1, данные Dempster).
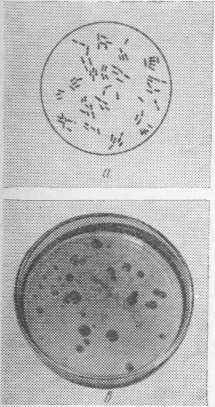
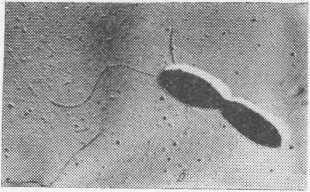
Рис. 2. Бактерии Pseudomonas fluorescens:
а — вид при окраске по Граму; б — вид под электронным микроскопом; в — колонии на мясопептонном агаре.
В I подгруппу включены пигментообразующие и сбраживающие глюкозу с образованием кислоты бактерии, а также бактерии, разжижающие и неразжижающие желатин. В нее входят мезофильные виды, способные расти при 42 °C и не дающие роста при 5 °C, и психротрофные — растущие при 5 °C и не дающие роста при 42 °C. Во II подгруппу включены бактерии, не обладающие способностью к пигментообразоваишо и образующие кислоту из глюкозы, в III подгруппу — бактерии, не обладающие способностью к пигментообразованию и подщелачивающие среду Хыога и Лейфсона, в IV — бактерии, не обладающие способностью к пигментообразованию и не изменяющие среду Хыога и Лейфсона.
К важным диагностическим свойствам психротрофных бактерий по схеме Хендри и Шевана относятся следующие тесты, позволяющие дифференцировать Pseudomonas от других грамотрицательных бактерий: оксидазный (по Ковачу); цитохромоксидазный (по Гейби и Фри); на аргинингидролазу (по Торнли). Согласно этим тестам бактерии Pseudomonas оксидазоположительные, содержат цитохромоксидазу, гидролизуют аргинин с образованием аммиака. Представителями бактерий I подгруппы, разжижающих желатин и наиболее часто встречающихся в мясных и молочных продуктах, являются бактерии Ps. fluorescens и Ps. geniculata. Большинство штаммов этих бактерий обладают протеолитической и липолитической активностью. По способности расщеплять белки протеолитические виды Pseudomonas занимают третье место после мезофильных спорообразующих анаэробов и бактерий Proteus.
Бактерии Ps. fluorescens, развиваясь в мясе и мясных продуктах, могут изменять pH среды от 5,6 до 8,6. Это дает основание отнести их к сильным аммонификаторам. Многие виды Pseudomonas выделяют липазу и активно расщепляют жир, что отрицательно сказывается на качестве жиросодержащих продуктов (жирное мясо, сливочное масло и пр.).
Минимальная температура роста бактерий Pseudomonas ниже О °C. Так, в переохлажденной жидкой питательной среде они размножаются при —5 °C. Продолжительность генерации при этом составляет 168 ч.
Представителями бактерий I подгруппы, не разжижающих желатина, являются Ps. putida, Ps. ovalis, Ps. rugosa, Ps. convexa. Эти бактерии обладают слабой липолитической активностью. Минимальная температура их роста —24—5 °C.
 Примечание. Н реакция положительная; реакция отрицательная
Примечание. Н реакция положительная; реакция отрицательная
К бактериям рода Pseudomonas, не образующим пигмента (II подгруппа), относятся Ps. fragi и Ps. putre- faciens. Эти бактерии способны разжижать желатин и разлагать белки в мясных и молочных продуктах при их холодильном хранении. Несмотря на то что большинство штаммов бактерий рода Pseudomonas II подгруппы липолитически неактивны, Ps. fragi обладает достаточно выраженной липолитической активностью. Этот вид бактерий выделен из жирного мяса, масла, сливок.
Бактерии III и IV подгрупп существенно не влияют на качество мясных и молочных продуктов при холодильном хранении.
Бактерии рода Pseudomonas очень чувствительны к нагреванию. Многие психрофильные штаммы этих бактерий начинают отмирать при 37 °C, а быстрая гибель наблюдается при 70°С и выше.
Рост бактерий рода Pseudomonas зависит от количества свободной воды в продукте, т. е. активности воды (aw). Интенсивное развитие бактерий происходит при aw=0,98; наименьшее значение aw, допускающее рост бактерий, составляет 0,91. Бактерии рода Pseudomonas широко распространены в природе.
Бактерии рода Aeromonas — это грамотрицательные факультативные анаэробы. Клетки от палочковидных с закругленными концами до кокковидных диаметром 1,0—4,4 мкм иногда образуют нити длиной до 8 мкм; встречаются поодиночке, парами или образуют цепочки; движутся с помощью полярных жгутиков, обычно моно- трихальных; некоторые виды неподвижны.
Метаболизм бактерий дыхательный и бродильный. Они расщепляют углеводы до кислоты или до кислоты и газа (СО2 и Н2). На мясопептонном агаре бактерии образуют небольшие беловато-серые, полупрозрачные, гладкие, блестящие колонии.
Большинство видов Aeromonas пигмента не образуют, однако Aeromonas salmonicida выделяют коричневый водорастворимый пигмент при росте на триптиказном соевом агаре. Для бактерий рода Aeromonas характерны положительные реакции на цитохромоксидазу, оксидазу и каталазу (см. табл. 1). Они гидролизуют казеин, крахмал, декстрин и глицерин, разжижают желатин, расщепляют глюкозу, фруктозу, мальтозу и трегалозу. Максимальная температура роста 38—41 °C (некоторые виды не растут при 37 °C), минимальная — 0—5 °C. Оптимальная температура роста для большинства видов 30 °C, хотя некоторые виды хорошо развиваются при 20 °C. Бактерии растут при pH 5,5—9,0, не развиваются в питательном бульоне с содержанием хлорида натрия 7,5%.
Бактерии рода Aeromonas довольно широко распространены в природе, особенно в водоемах с пресной водой, откуда они могут попасть на шкуру животных, а затем в молоко и мясо. Являясь факультативными анаэробами, бактерии вызывают порчу продуктов при их холодильном хранении. Порче подвергаются продукты, хранящиеся в аэробных условиях и различных модифицированных средах (углекислый газ, азот), а также упакованные под вакуумом в газонепроницаемые пленки.
Имеются сведения, что отдельные виды Aeromonas, вызывающие заболевания некоторых животных, являются потенциально патогенными для человека.
Бактерии рода Brochothrix представляют собой грамположительные палочки с закругленными концами размером 1,0x8,0 мкм, неподвижные, слабо ферментируют углеводы с образованием £(+)—молочной кислоты. Для них типично угловое и палисадное расположение клеток. Оптимальная температура роста бактерий примерно 30 °C, при 40 °C не развиваются, минимальная температура около 10 °C. Однако имеются сообщения, что типичный представитель этого рода Brochothrix ther- mosphactum давал рост при 4 °C. Довольно устойчивы к нагреванию: выдерживают температуру 72 °C в течение 15 мин в обезжиренном молоке. Бактерии часто встречаются в молочных продуктах, на молочном оборудовании, а также на упакованном в пленку охлажденном мясе при его хранении.
Бактерии рода Flavobacteriutn представляют собой грамотрицательные факультативно-анаэробные палочки. Они неподвижны или перемещаются посредством перитрихальных жгутиков. Метаболизм дыхательный (см. табл. 1).
На поверхности твердых питательных сред бактерии образуют колонии желтого, оранжевого, коричневого цветов. Цвет колоний наиболее четко проявляется при посеве на желатин или в молокосодержащие среды и зависит от температуры — более выражен при пониженных температурах (15—20°C). В связи с этим при идентификации бактерий предпочтительно инкубацию осуществлять при температуре ниже 30 °C; при более высоких температурах рост может быть ограничен, хотя имеются сообщения, что отдельные виды растут при 37 °C.
Бактерии выделяют пигменты, нерастворимые в питательных средах, природа которых неизвестна. Предполагают, что они представляют собой каротиноиды. Большинство видов Flavobacterium гидролизуют желатин и казеин. Наиболее часто они встречаются в молоке и молочных продуктах.
Бактерии рода Alcaligenes — это грамотрицательные одиночные кокковидные палочки или кокки размером 0,5—1,2x0,5—2,6 мкм. Движутся бактерии с помощью перитрихальных жгутиков, строгие аэробы; оксидазоположительные (тест Ковача); быстро растут в средах с pH 7, не ферментируют углеводы.
Колонии различных видов бактерий на мясопептонном агаре весьма разнообразны — мелкие, плоские, коричневатые, серые, грязно-белые, прозрачные с непрозрачным центром и волнистым краем. Оптимальная температура роста 22—37 °C.
Бактерии рода Alcaligenes могут вызывать порчу молока и молочных продуктов при длительном холодильном хранении, хотя протеолитическая способность у них выражена слабо. При развитии бактерий Alcaligenes viscosus молоко становится слизистым или тянущимся.
Плесневые грибы. Большинство плесневых грибов (плесеней) растут при низких положительных, а отдельные роды — при отрицательных температурах. Это дает основание отнести их к психротрофным микроорганизмам. Плесневые грибы широко распространены в природе. Они встречаются в воде, воздухе, почве, на растениях и кожном покрове животных; могут расти на древесине, бумаге, оштукатуренных и окрашенных стенах, резине, кожаной обуви и многих других объектах. Наиболее благоприятной средой для развития плесневых грибов являются пищевые продукты, в том числе мясные и молочные. Вследствие высокой биохимической активности плесени способны очень быстро окислять и расщеплять углеводы, жиры, белки, вызывать изменения самых различных веществ.
Плесени наносят большой вред народному хозяйству. Однако в настоящее время их с успехом направленно используют для получения различных ценных соединений, таких, как пенициллин, ацетон, бутиловый спирт. Плесени нашли применение и в пищевой промышленности, в частности для придания продукту специфических вкуса и запаха (например, в сыроделии при производстве сыра рокфор и камамбер), а также в качестве катализаторов биохимических процессов при созревании отдельных видов сырокопченых колбас.
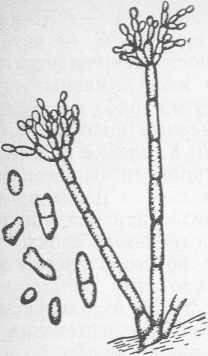
Рис. 3. Cladosporium herbarum
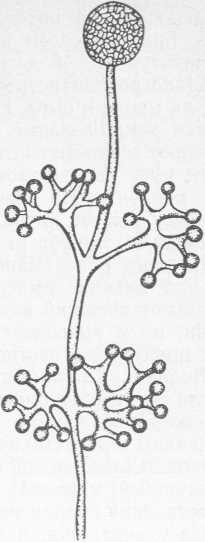
Рис. 4. Thamnidium elegans
Плесени — строгие аэробы. В отличие от бактерий они могут развиваться при высоких осмотическом давлении и кислотности среды, а также недостаточном количестве свободной воды. Минимальное значение активности воды для плесеней составляет 0,7—0,8.
В мясных и молочных продуктах при холодильном хранении наиболее часто развиваются и вызывают их порчу следующие роды плесеней: Cladosporium, Thamnidium, Penicillium и др. Признаки плесеней описаны согласно определителю Джильмена.
Колонии Cladosporium herbarum (рис. 3) плоские, бархатистые, оливково-зеленые или серо-зеленые; поверхность, примыкающая к питательной среде на чашке Петри,— черная или зеленовато-черная. На концах конидиеносцев образуются главным образом одноклеточные конидии различной формы: округлые, лимоновидные или цилиндрические. Видимый рост у Cladosporium sp. наблюдается на питательной среде при —5 °C на 10-й день. Гриб способен расти и при —8 °C (в интервале температур от —8 до —12°C рост прекращается).
Cladosporium встречается в мясе и мясных продуктах, на масле и сыре. При развитии гриба на мясе образуются темно-зеленые, почти черные колонии, которые врастают в поверхностный слой. Удаление их с поверхности мяса посредством среза приводит к потерям ценного продукта. На поверхности сыра и масла он также образует темные пятна, а в масле (при наличии пустот в монолите) может расти и в глубинных слоях.
Плесени рода Cladosporium протеолитически активны при низких температурах, поэтому они не только ухудшают внешний вид продукта при холодильном хранении, но и вызывают биохимические изменения в самом продукте, в частности протеолиз белков.
Колонии Thamnidium elegans (рис. 4) белые или светло-серые с длинными, нитевидными, прямо стоящими спорангиеносцами, на которых невооруженным глазом видны сероватые пушинки. Мицелий развивается как в глубине питательной среды, так и над ее поверхностью (воздушный мицелий). При плодоношении образуются разветвления спорангиеносцев в форме пушистых пучков (реже у верхушки, чаще повсеместно), мутовчато расположенных один против другого. Главный спорангиеносец заканчивается круглым спорангием диаметром 100— 200 мкм с колонкой длиной 60—90 мкм, шириной 50— 70 мкм. На разветвлениях спорангиеносцев имеются спорангиоли различной величины — от очень мелких до крупных диаметром 24 мкм.
Thamnidium elegans развивается при температуре —8-1—10 °C; при —12 °C роста не наблюдается в течение 400 дней выдержки посевов в незамерзшей питательной среде. Наиболее часто эта плесень встречается на мясе и мясных продуктах при их холодильном хранении. Thamnidium elegans обладает выраженными протеолитическими свойствами, в результате чего белки мяса подвергаются распаду, который сопровождается появлением неприятного запаха. Для удаления глубинного мицелия разросшегося плесневого гриба приходится зачищать или срезать поверхностный слой мяса, что приводит к существенным потерям продукта.
Penicillium glaucum (рис. 5) вызывают порчу мясных и молочных продуктов при холодильном хранении.
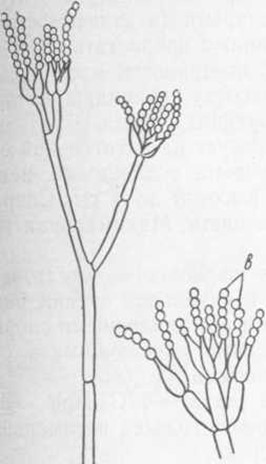
Рис. 5. Penicillium glaucum: a — конидиеносец; б — разветвленный конидиеносец (кисточка), в — цепочки конидий
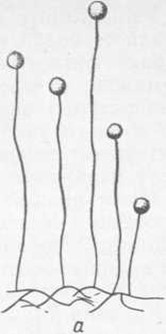
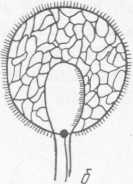
Рис. 6. Mucor mucedo:
а — конидиеносцы с головками; б — головка
Колонии этого вида плесени серовато-зеленого или синевато-зеленого цвета; конидиеносцы прямые, многоклеточные, различной высоты. Концы конидиеносцев разветвлены в виде кисти или вилки, на них образуются цепочки конидий бесцветные или слабо-зеленого цвета.
Колонии Penicillium brevicaule светло-коричневые, неравномерно пушистые; на поздней стадии развития становятся слизистыми с неприятным запахом мышьяковистого водорода. Конидиеносцы чаще неразветвленные, короткие. Конидии на довольно длинных стеригмах, желтовато-бурые, крупные или лимоновидные, сначала гладкие, затем шероховатые и иглистые.
Плесневые грибы рода Penicillium растут при отрицательных температурах: Penicillium sp. при —9 °C, Р. glaucum при —5 °C.
Penicillium с поверхности мяса и сыра удаляют, обмывая ее водой и зачищая, а масла — только зачищая. Однако полностью удалить споры этими способами невозможно, и через некоторое время (в зависимости от температуры) они вновь начинают прорастать. Для более полного удаления спор с поверхности мяса и сыра применяют специальную обработку, принципы которой будут изложены в соответствующих главах.
Mucor mucedo (рис. 6) образует на питательной среде колонии желтовато-серого цвета с длинными, неветвящимися спорангиеносцами высотой до 4 см. Спорангии крупные желтого и серого цвета. Минимальная температура роста 2 °C.
Колонии Mucor racemosus шелковистые, желтовато-белого, желто-бурого цветов, в начальной стадии роста малопушистые, затем с тонкими разветвленными спорангиеносцами, усыпанными мелкими сливающимися спорангиями, кажущимися слизистыми.
Минимальная температура роста —4 °C. При —8 °C роста гриба не наблюдалось более года на незамерзшей среде.
Плесневые грибы рода Mucor хорошо растут на хлебе, пищевых остатках, овощах. Поэтому появление плесеней на мясных и молочных продуктах можно объяснить контактом их с обсемененными продуктами (в основном хлебом). Колонии Rhizopus nigrificans (рис. 7) рыхлые, с белым паутинистым мицелием, позднее серым. Спорангиеносцы высотой около 2—4 мм пучками отходят от одной общей точки, с большими черными головками диаметром 100—350 мкм, споры серые, круглые или овальные, угловатые различной величины. При температуре от —5 до—8°C Rhizopus прекращает рост. При развитии на поверхности мяса, как и Thamnidium, прорастает в поверхностный слой его.
Колонии Geotrichum candidum (рис. 8) развиваются в виде бархатистого белого налета, с возрастом не изменяющегося. Конидии образуются простым делением мицелия и распадаются на отдельные бесцветные, прямоугольные клетки, которые, иногда соединяются в цепочки размером (10—30) X 3,5 мкм.
Вид Geotrichum candidum (молочная плесень) встречается очень часто на молочных продуктах при холодильном хранении. Обсеменение продуктов происходит в процессе их выработки и в основном при контакте с плохо вымытым оборудованием и через воздух. Вследствие способности к росту при низких положительных температурах плесневый гриб может развиваться на твороге, сметане, сыре (при их холодильном хранении). Обладая активными липолитическими свойствами, Geotrichum candidum вызывает быстрое расщепление жира, в результате чего происходит прогоркание продукта. Растет при —4 °C.
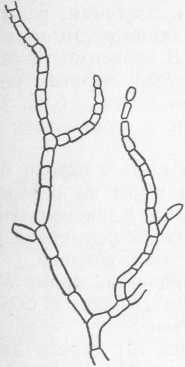
Рис. 8. Geotrichum candidum
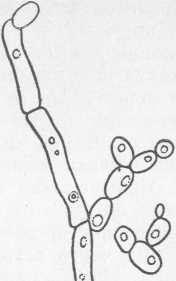

Рис. 9. Monilia
Колонии Monilia (Candida) (рис. 9) в начале развития сходны с колониями дрожжей, так как этот вид является переходным от дрожжей к плесеням; размножается почкованием. На поздних стадиях роста образуется мицелий с воздушными нитями. В зависимости от вида колонии бывают окрашены в белый, черный, розовый цвета. Минимальная температура роста —5 °C. Биохимическая активность проявляется в способности вызывать спиртовое брожение.
Monilia часто встречается на сыре и масле при их хранении. При развитии Monilia nigra на поверхности продуктов образуются черные пятна. Корковый слой сыра в результате выделения плесенью ферментов разрушается и плесень проникает в глубь головки.
На масле при холодильном хранении, кроме Monilia nigra, может развиваться Monilia roseum, с образованием на поверхности розовых пятен.
Колонии Botrytis cinerea (рис. 10) серого цвета, с крупными рыхлыми конидиеносцами. Иногда из мицелия возникают коричневые или черные склероции размером с дробинку или горошинку. Конидиеносцы высотой около 1 мм разветвляются наподобие виноградной грозди. Концы разветвлений имеют вздутия, на которых находятся многочисленные стеригмы, образующие по одной желтовато-бурой, эллиптической или круглой конидии с гладкой оболочкой. Конидии можно легко распознать по значительному размеру: 9—15x6,5—10 мкм.
На пищевых продуктах при холодильном хранении при —5 °C растут виды Botrytis cinerea и Botrytis sp.
Колонии Aspergillus glaucus (рис. 11) желтовато-зеленые. Конидиеносцы рыхлые, в большинстве случаев различимые невооруженным глазом, короткие (1—2 мм). Размножаются грибы как с помощью конидий, так и спор. Споры диаметром 6—10 мкм образуются в овальных сумках (аскусах), которые в свою очередь заключены внутри круглого тела (клейстокарпии), окрашенного в лимонно-желтый цвет. Споры бесцветные, с поперечными бороздами, в каждой сумке по 8 спор.
Колонии Aspergillus niger сначала белоснежные, затем приобретают темно-коричнево-черную окраску, у края более светлую, рыхлобархатистые. Конидиеносцы обычно несептированные, с круглым утолщением на конце, диаметром около 80 мкм. Разветвленные стеригмы (первичные длинные и узкие, вторичные короткие) располагаются вокруг утолщения. Конидии мелкие, круглые, сначала гладкие, затем шероховатые, темно-коричневые.
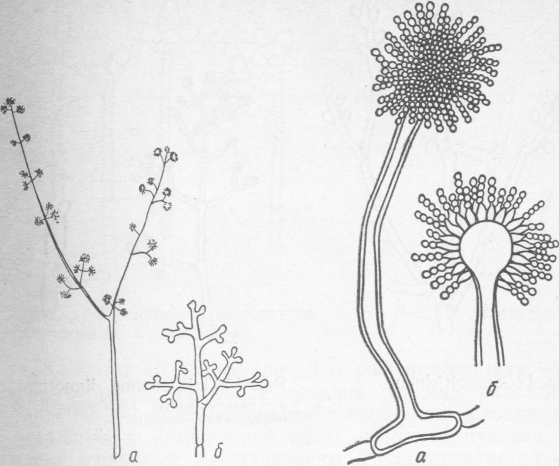
Рис. 10. Botrytis cinerea — конидиеносцы с конидиями; о — верхушечные участки
Плесневые грибы рода Aspergillus редко встречаются на мясе при холодильном хранении, так как минимальная температура их роста выше, чем температура хранения мяса. Чаще эти виды плесени встречаются на молочных продуктах и особенно на сыре в процессе его созревания при 10—12 °C. Aspergillus может развиваться на масле при нарушении температуры его хранения.
Имеются сообщения, что плесени рода Aspergillus образуют токсины, которые могут переходить в продукт (особенно в сыр).
Колонии Sporotrichum (рис. 12) белого цвета. Мицелий стелящийся, ветвистый. Конидиеносцы развиваются в виде коротких ветвей. Конидии одиночные, одноклеточные, шаровидной или яйцевидной формы. Они образуются на очень коротких выростах гиф.
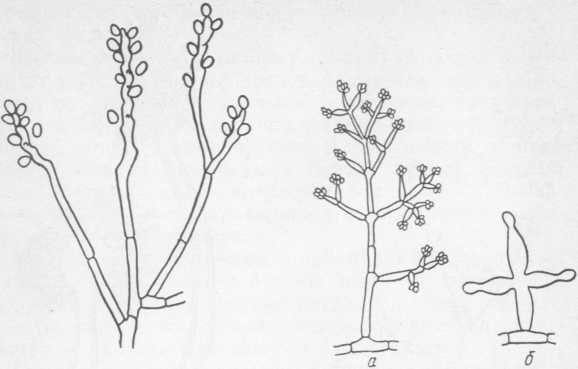
Рис. 12. Sporotrichum Рис. 13. Trichoderma lignorum: a — разветвленный конидиеносец; б — образование конидий
Грибы этого рода, развивающиеся преимущественно на мясе, имеют нижний температурный предел роста при —6,5 °C, а вид Sporotrichum caris развивался при -8 °C.
Колонии Trichoderma lignorum (рис. 13) зеленовато-желтоватые. Конидиеносцы разветвленные. Конидии зеленоватого цвета, шаровидные, размещаются в округлых головках, собранных вместе по 10—20 шт. На чашках Петри с питательной средой дает характерный концентрический рост. Встречаются чаще на охлажденном мясе. Нижний температурный предел роста от 1,1 до 2,2 °C.
Колонии Trichotecium roseum (рис. 14) абрикосовокрасного цвета от покрывающих белый мицелий окрашенных конидий; колонии обнаруживают ясный концентрический рост. Конидиеносцы длинные, тонкие, несептированные (без перегородок) и не разветвленные. На конце конидиеносца образуются крупные, грушевидные, сначала бесцветные, затем светло-красные конидии. Trichotecium roseum растет на пищевых продуктах при холодильном хранении.
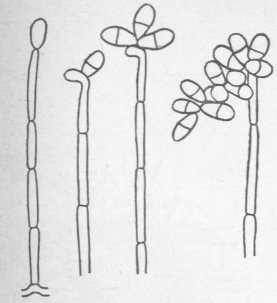
Рис. 14. Trichotecium roseum (конидиеносцы с конидиями)
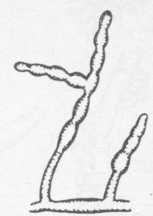
Рис. 15. Alternaria
Колонии Alternaria (рис. 15) желтовато-оливково-зеленого цвета, на более поздних стадиях развития — оливково-коричневые, нижняя поверхность колонии, прилегающая к питательной среде, темно-коричневая, мицелий пушистый. Конидиеносцы в большинстве случаев представляют собой короткие боковые ответвления мицелия, иногда вильчато-разветвленные; конидии крупные, многоклеточные, кругловатые или грушевидные от темно-оливкового до черного цвета.
Колонии Ptillularia pullulans (рис. 16) в начальной стадии роста имеют белый, розоватый, рыжевато-коричневый цвета, слизистую консистенцию. На более поздних стадиях роста колонии приобретают зеленый, а затем черный цвет; консистенция становится жесткой, кожистой; характерно образование по краю колоний корневидной разрастающейся каймы. Конидии образуются непосредственно на мицелиях, веретенообразные или про- долговато-круглые. Этот гриб может размножаться и путем распада мицелия.
Колонии Photna (рис. 17) вначале белые, затем темные. Мицелий паутинистый, стелящийся по поверхности субстрата, с черными пятнами кругловатых или овальных пикнидий, в которых образуются конидии. По достижении зрелости конидии высыпаются через отверстия в пикнидиях. Конидии одноклеточные, бесцветные, веретенообразной или эллиптической формы. Грибы рода Phoma растут при О С°; могут давать рост при ограниченном содержании кислорода в субстрате, поэтому они встречаются внутри монолита масла (черные пятна).
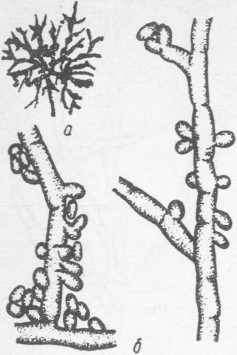
Рис. 16. Pullularia pullulans:a — молодая колония; б — гифы, образование конидий
Рис. 17. Phoma
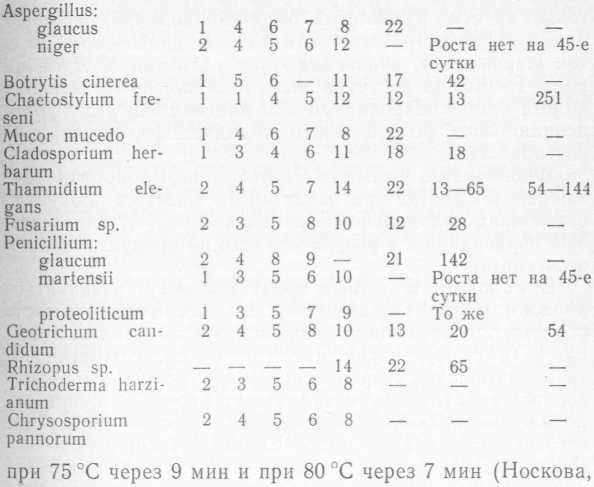
Все плесневые грибы распространяются главным образом через воздух. Наличие их на мясных и молочных продуктах и видовой состав зависят в первую очередь от условий окружающей среды. Даже в одних климатических зонах в зависимости от времени года, степени запыленности воздуха, поступающего в производственные цеха и холодильные камеры, может наблюдаться большое разнообразие видового состава плесневых грибов. При холодильном хранении важно знать срок начала роста колоний плесневого гриба в зависимости от температуры. По наличию роста плесневых грибов на продукте можно судить о соблюдении температурных режимов хранения продукта. Время (сутки) от посева до появления роста у различных видов плесневых грибов в зависимости от температуры представлены в табл. 2 (Г. Л. Носкова, 3. 3. Бочарова, А. А. Буканова и др.). Споры плесневых грибов погибают при тепловой обработке. Так, споры Mucor mucedo погибали при 70— 75 °C после 9-минутной выдержки, при 80 °C после 5— 8-минутной выдержки, но переносили нагревание при 65 °C в течение 15 мин. Споры Cladosporium herbarum также выдерживали нагревание при 65 °C в течение 15 мин, но полностью погибали при 70 °C через 13 мин.
Таблица 2

Дрожжи. Дрожжи представляют собой одноклеточные крупные (10—15 мкм) неподвижные микроорганизмы. Размножаются они главным образом путем почкования и реже — деления (рис. 18). Большинство дрожжей аэробы.
Рис. 18. Дрожжи (различные виды)
На мясе и мясных продуктах при холодильном хранении развиваются дрожжи семейства Cryptococcaceae, родов Candida,, Cryptococcus,, Rhodotorula,, Torulopsis и Trichosporon. Эти дрожжи являются аспорогенными, т. е. не образуют спор. Размножаются они почкованием. На мясе чаще обнаруживаются дрожжи родов Rhodotorula и Torulopsis. При росте на питательных средах, а также на мясе Rhodotorula образуют колонии, окрашенные в красный, розовый или желтый цвет. Форма клеток шаровидная, яйцевидная или удлиненная. Дрожжи рода Torulopsis растут в виде неокрашенных колоний. Форма клеток шаровидная или яйцевидная, реже удлиненная. Этот род дрожжей обладает ферментативной способностью.
Большинство штаммов, Cryptococcus,, Rhodotorula, Torulopsis и Candida при развитии на пищевых продуктах в условиях холодильного хранения образуют внеклеточный полисахарид, в результате чего на продукте появляется слизь.
В молоке и молочных продуктах могут развиваться дрожжи рода Saccharomyces. Из них наиболее часто встречаются следующие спорообразующие виды: Sacch. casei (в сыре),, Sacch. lactis и Sacch. cartilaginosus (в кисломолочных продуктах). Все эти виды дрожжей сбраживают глюкозу и галактозу. Кроме того, при холодильном хранении молока и молочных продуктов встречаются дрожжи родов Candida, Torulopsis и Му- coderma. Дрожжи рода Candida способны образовывать примитивный мицелий (псевдомицелий). Некоторые виды обладают липолитической способностью. Дрожжи рода Mycoderma относят к дрожжеподобным микроорганизмам, которые являются как бы промежуточным звеном между дрожжами, и плесневыми грибами, родов Monilia и Geotrichum. Размножаются почкованием, причем почки не отделяются от материнской клетки, образуя скопления из объединенных между собой клеток. На жидких питательных средах такие скопления клеток образуют пленку.
Дрожжи рода Mycoderma обладают протеолитической способностью; развиваются на поверхности кисломолочных продуктов и мягких сыров. Оптимальная температура развития дрожжей 25—30 °C. Многие дрожжи способны размножаться при температуре —34—5 °C, а отдельные виды — при —84—10 °C.
Активность роста отдельных штаммов дрожжей зависит от температуры. Например, появление видимого роста у штамма Torula sp. отмечено при 2 °C через 7 сут, при 0°С—10 сут; —2°С — 21 сут; —5°С — 38 сут; при —8 °C рост не наблюдался в течение 9 мес (Носкова, 1979). Рост при —5 °C характерен для часто встречающихся дрожжей Monilia nigra («черные дрожжи»), являющихся дрожжевидной формой гриба Auerobasidium pullulans.
Из мясных и молочных продуктов выделены психротрофные дрожжи родов Debaryomyces и Pichia, которые росли при холодильном хранении этих продуктов.
Большинство видов дрожжей, развивающихся на мясных и молочных продуктах, обладают липолитической способностью. В связи с этим размножение дрожжей в жиросодержащих продуктах при холодильном хранении вызывает их порчу: прогоркание, осаливание, появление неприятного запаха.
Актиномицеты. Они относятся к семейству Actinomy- cetaceae и представляют собой бактерии, способные образовывать ветвящиеся гифы, которые могут развиваться в мицелий, т. е. обладают признаками, характерными для плесневых грибов (рис. 19). Это грамположительные, неподвижные, факультативные анаэробы, хотя имеются виды, которые развиваются в анаэробных и аэробных условиях. Диаметр клеток обычно менее 1 мкм, длина гифов и мицелия не превышает нескольких миллиметров. На питательных средах образует плотные жесткие колонии (диаметр 1 —10 мм), прочно врастающие в среду. Поверхность колоний бархатистая, что обусловлено структурой мицелия.
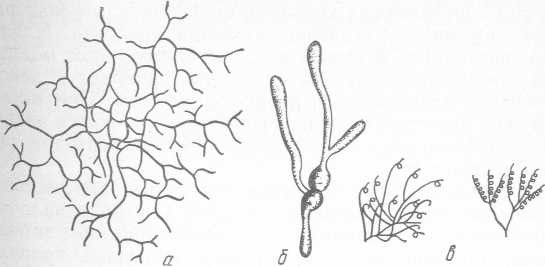
Рис. 19. Актиномицеты:
— общий вид мицелия; б — прорастание спор; в — строение спороносцев
Характерной особенностью актиномицетов при росте на питательных средах является появление землистого запаха, по которому можно судить об их наличии. Actinomyces сбраживают глюкозу, в результате чего образуются уксусная, молочная, муравьиная и янтарная кислоты.
Актиномицеты наиболее часто встречаются в почве, реже — в воде, откуда они попадают на шкуру животного, а затем в мясо и молоко. Большинство видов для роста нуждается в диоксиде углерода, поэтому они могут расти на мясе, упакованном в пленку в среде углекислого газа. Оптимальная температура роста 25—37 °C, но могут расти и при —24—3 °C. Некоторые виды патогенны, например Actinomyces bovis.
Мезофильные микроорганизмы
Мезофильные микроорганизмы, встречающиеся в мясных и молочных продуктах, по их воздействию на продукт можно разделить на полезные и вредные. К полезным относятся молочнокислые бактерии (стрептококки и палочки), на метаболическом действии которых основана технология кисломолочных продуктов, сыра и кислосливочного масла. Эта микрофлора используется также в производстве сырокопченых колбас, ветчины. Однако при определенных условиях молочнокислые бактерии могут вызывать ухудшение качества продукта. Например, развиваясь в сыром или пастеризованном молоке, они повышают его кислотность, что приводит к невозможности дальнейшей переработки молока. При развитии этих бактерий на поверхности упакованного в пленку охлажденного мяса или фарша качество продуктов также ухудшается.
К вредным относятся микроорганизмы, вызывающие порчу продуктов (например, протей, кишечная палочка, маслянокислые бактерии), желудочно-кишечные заболевания (энтеропатогенные штаммы кишечной палочки, сальмонеллы), а также пищевые отравления (стафилококки, клостридии). Несмотря на то что эти бактерии являются типичными мезофилами, некоторые из них могут размножаться в продукте в процессе его хранения при температуре ниже 10 °C (табл. 3, данные Rosset, 1985).
Таблица 3

По наличию и количеству этих бактерий в продукте судят о его безопасности для потребителя, а также санитарно-гигиеническом состоянии предприятий, выпускающих мясные и молочные продукты.
Бактерии. Бактерии родов Streptococcus, Leuconostoc и Lactobacillus очень широко используются в молочной промышленности в качестве заквасок для кисломолочных продуктов, сыров, кислосливочного масла. При изготовлении сырокопченых колбас применяют отдельные виды Lactobacillus. Молочнокислые бактерии подробно описаны многими авторами (Королев, 1974; Королева, 1975; Богданов, 1969; Банникова, 1975 и др.), поэтому ниже будет дана лишь краткая их характеристика.
Клетки бактерий рода Streptococcus (рис. 20) сферические или овальные диаметром около 2 мкм. При росте в жидкой среде клетки располагаются парами пли цепочками.
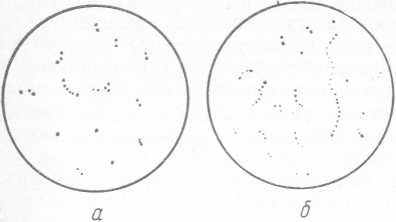
Рис. 20. Молочнокислые стрептококки: а — S. lactis; б — S. cremoris
Колонии этих бактерий на твердой питательной среде мелкие (диаметр 1—2 мм), каплевидные, беловатого цвета; глубинные колонии имеют вид чечевичек; в серологической группе Д (молочнокислые стрептококки фекального происхождения) встречаются штаммы, образующие желтый или красный пигмент. Бактерии грамположительные, неподвижные, факультативные анаэробы, метаболизм бродильный.
Гомоферментативные молочнокислые стрептококки (S. lactis и S. cremoris) образуют из глюкозы молочную кислоту. Для этих видов бактерий рода Streptococcus характерна высокая активность кислотообразования. Гетероферментативные молочнокислые стрептококки (S. diacetilactis и S. acetoinicus) в процессе жизнедеятельности образуют летучие кислоты, диацетил, ацетоин. Температурные пределы роста молочнокислых стрептококков от 10 до 40 °C, оптимальная температура —30 °C.
К роду Streptococcus относятся также молочнокислые бактерии фекального происхождения (стрептококки группы Д по Ленсфильд): S. faecalis, S. faecium, S. bovis. Морфологические свойства их близки свойствам группы молочнокислых стрептококков. Характерные отличительные признаки следующие: рост при 45 °C; развитие в среде с содержанием 6,5% хлорида натрия при pH 9,6.
Эти бактерии постоянно содержатся в кишечнике человека и животных, поэтому для пищевых продуктов они могут иметь значение санитарно-показательных микроорганизмов. При длительном хранении продуктов следует проводить учет этих микроорганизмов, так как стрептококки группы Д более устойчивы к воздействиям внешней среды (высокой и низкой температурам, pH, содержанию солей и пр.), чем бактерии группы кишечной палочки. В этих условиях кишечная палочка погибает быстрее и ее не удается выявить, следовательно, по наличию или отсутствию стрептококков группы Д можно судить о санитарном благополучии продукта. Способность стрептококков группы Д вызывать пищевые отравления достоверно не установлена, хотя сообщения об этом имеются (Седова, 1970).
Клетки бактерий рода Leuconostoc сферической или чечевицеобразной формы, обычно располагающиеся парами или цепочками (рис. 21). На твердых питательных средах они растут в виде мелких (диаметр 1 мм), гладких, серовато-белых колоний. Бактерии неподвижные, грамположительные, требовательные к питательной среде (нуждаются в комплексе факторов роста и аминокислотах), факультативные анаэробы, гетероферментативные, глюкозу сбраживают с образованием молочной кислоты, этанола и диоксида углерода, метаболизм бродильный. Некоторые штаммы бактерий рода Leuconostoc имеют окислительный механизм и вместо этанола продуцируют уксусную кислоту.
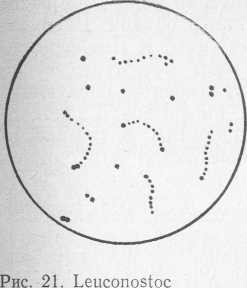
В молоке и молочных продуктах наиболее часто встречаются виды L. lactis, L. cremoris и L. dextrani- cum, которые входят в состав заквасок для кисломолочных продуктов и сыра. Температурные границы роста бактерий рода Leuconostoc 10—39 °C, оптимальная температура 20—30 °C. Выделенные нами из охлажденного молока бактерии L. lactis росли при 5 °C.
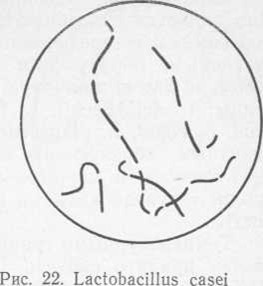
Бактерии рода Lactobacillus представляют собой палочки (от длинных тонких до коротких, напоминающих коккобактерии). Клетки обычно располагаются поодиночке, но в поздней логарифмической фазе роста они часто образуют цепочки (рис. 22). На твердых питательных средах бактерии растут в виде очень мелких «точечных» беловатых колоний. Бактерии рода Lactobacillus грамположительные, в основном, неподвижные (подвижные формы встречаются очень редко), факультативные анаэробы, метаболизм бродильный. Активный рост бактерий наблюдается при анаэробных условиях и при содержании в среде 5—10% диоксида углерода. Оптимум pH составляет 5,5—5,8, однако бактерии могут развиваться при pH 5 и ниже.
Характерной особенностью бактерий рода Lactobacillus является способность расщеплять сахара. Различают гомо- и гетероферментативные типы. К гомоферментативным, образующим из глюкозы преимущественно (85% и более) молочную кислоту, относятся следующие виды: L. delbrueckii, L. lactis, L. bulgaricus, L. helveti- cus, L. casei, L. plantarum, L. acidophilus. К гетерофер- ментным, образующим из глюкозы около 50% молочной кислоты и значительные количества диоксида углерода, уксусной кислоты и этанола,— L. brevis, L. buchneri.
Температурными границами роста молочнокислых палочек принято считать 10—53°C (оптимум 30—40°C). В «Кратком определителе бактерий Берги» (1980) приведена минимальная температура роста, равная 5 °C. Нами из сыра были выделены L. casei и L. brevis, которые росли при 5 и 2 °C. Развитие молочнокислых палочек наблюдалось на мясе, упакованном под вакуумом и хранившемся при 0°С. Бактерии рода Lactobacillus встречаются в молочных продуктах, смывах с молочного оборудования, мясных продуктах, воде, сточных водах.
При холодильном хранении молока и молочных продуктов наибольший интерес представляют нижние температурные границы роста мезофильных молочнокислых бактерий. Способность отдельных видов и штаммов молочнокислых бактерий развиваться при температуре ниже 10 °C позволяет считать, что среди молочнокислых бактерий встречаются психротрофные виды. Возможно, что вследствие широкого использования холода при получении, переработке и хранении молока произошла адаптация отдельных видов молочнокислых бактерий к более низким температурам, в связи с чем изменилась и минимальная температура их роста. Способность молочнокислых бактерий развиваться при температуре ниже 10 °C следует учитывать при выборе режимов и сроков хранения молока, молочных продуктов, а также мяса, упакованного под вакуумом.
Нежелательными и даже опасными для здоровья человека являются мезофильные бактерии семейства Enterobacteriaceae родов Escherichia, Enterobacter, Proteus, Salmonella. Характеристика этих бактерий подробно дана многими авторами, поэтому мы лишь кратко рассмотрим их основные признаки.
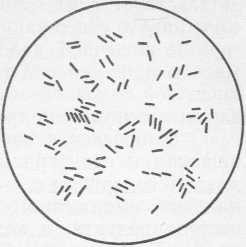
Рис. 23. Escherichia coli
Бактерии рода Escherichia (рис. 23)— это факультативно-анаэробные. грамотрицательные палочки размером 1,1 — 1,5x2,0—6,0 мкм (живые) или 0,4—0,7X1,0—3,0 мкм (высушенные и окрашенные), располагающиеся отдельно или парами. Подвижные бактерии перемещаются посредством перитрихальных жгутиков, однако имеются и неподвижные формы. На питательном агаре развиваются гладкие колонии (S-форма), слабовыпуклые с блестящей влажной поверхностью, ровным краем, сероватые; могут также образовываться колонии с шероховатой поверхностью; встречаются и слизистые формы.
Характерной особенностью бактерий этого рода является их способность сбраживать глюкозу при 37 °C в течение 24 ч с образованием различных кислот (молочная, уксусная, муравьиная) и газа. При росте на фуксинсульфатном агаре (среда Эндо) образуются типичные колонии — красные с металлическим блеском, темно-красные, розовые с темным центром и прозрачные неокрашенные. Типичным представителем бактерий рода Escherichia является Escherichia coli — фекальная кишечная палочка.
Бактерии группы кишечной палочки различаются оптимальной температурой роста. Так, у Е. coli она составляет 43 °C, у других видов — 37 °C. Несмотря на то что бактерии рода Escherichia типичные мезофилы, имеются сообщения о их развитии при температуре ниже 10 °C. Например, в молоке они размножаются при 3—5 °C. Из мясных продуктов были выделены штаммы бактерий группы кишечной палочки, которые хорошо росли при 1,5 °C (Носкова, 1972).
Бактерии группы кишечной палочки неустойчивы к высоким температурам (погибают при 60 °C и выдержке 15 мин). Они сравнительно устойчивы (3—6 мес) в за-мороженной при —18 °C среде.
Основная среда обитания бактерий рода Escherichia — кишечник человека и животных, в связи с чем присутствие этих бактерий в пищевых продуктах свидетельствует о их фекальном загрязнении. При плохом санитарном содержании животных на ферме бактерии группы кишечной палочки попадают в молоко или на мясо в процессе его выработки. Обсеменение продукта кишечной палочкой обусловлено несоблюдением элементарных санитарных требований.
О безопасности продукта в микробиологическом отношении и санитарном благополучии производства судят по содержанию в нем бактерий группы кишечной палочки, выраженного либо титром (наименьшее количество продукта, в котором обнаружена кишечная палочка), либо количеством колоний, выросших на твердых питательных средах.
Молоко и почти все молочные продукты, готовые быстрозамороженные! продукты, мороженое нормируются по содержанию в них бактерий группы кишечной палочки.
В семейство Enterobacteriaceae входят также бактерии группы кишечной палочки, обитающие во внешней среде и относящиеся к роду Enterobacter. Представителем этого рода является Enterobacter aerogenes (ранее определялся как Coli. aerogenes). Основные отличительные признаки бактерий рода Escherichia от Enterobacter приведены в табл. 4.

Образование:
индола ± —
ацетилметилкарбинола ± —
сероводорода — ±
уреазы — ±
Реакция с метиловым красным + —
Использование цитрата — +
Примечание: + --------- положительный результат; - ---------------- отрицательный результат; ± — возможен положительный и отрицательный результат.
Бактерии рода Proteus — это факультативно-анаэробные грамотрицательные палочки размером 0,4—0,6Х X 1,0—3,0 мкм, располагающиеся парами или цепочками (рис. 24); очень подвижные (движутся посредством перитрихальных жгутиков), причем подвижность хорошо выражена при 20 °C и может полностью отсутствовать при повышении температуры до 37 °C.
На мясопептонном агаре (МПА) бактерии рода Proteus дают полупрозрачный вуалеобразный рост, особенно характерный на скошенном МПА (тест Щукевича), или роение.
Характерным признаком бактерий Proteus является высокая протеолитическая активность.
Бактерии рода Proteus (типичный представитель вид Proteus vulgaris), широко распространенные во внешней среде, также обнаруживаются в кишечнике человека. Основные биохимические признаки Proteus vulgaris следующие: сбраживает глюкозу, мальтозу, сахарозу с образованием кислоты и газа; не сбраживает лактозу и манниты; образует сероводород и индол, разлагает мочевину.
Температурные границы роста этих бактерий 10— 43 °C, однако имеются данные (Носкова, 1972) о их развитии при 5 и даже 0 °C. Наши исследования показали, что бактерии Proteus хорошо развиваются в свежем мясном фарше при 10 °C; продолжительность генерации в этом случае составляет 8,2 ч. При 5 °C на мясе не наблюдалось роста в течение 6 сут.
Бактерии Proteus погибают в результате нагревания при 63 °C в течение 30 мин. Однако они устойчивы к замораживанию. Так, бактерии не погибали в мясном фарше в течение 4 мес при —18 °C.
Бактерии Proteus чаще встречаются в мясных продуктах, чем в молочных. Являясь активными протеолитами, они могут вызвать быструю порчу мяса (гнилостное разложение) с появлением неприятного запаха. В пищевые продукты бактерии Proteus попадают при нарушении санитарного режима производства. Имеются сообщения, что бактерии Proteus могут вызывать пищевые токсикоинфекции. Заболевание возникает при употреблении сильно обсемененных продуктов без внешних признаков порчи.

Бактерии рода Salmonella — это факультативно-анаэробные грамотрицательные подвижные палочки размером 2—4X0,5 мкм (рис. 25). Движение осуществляется с помощью перитрихальных жгутиков. На МПА растут в виде круглых гладких полупрозрачных колоний. Для большинства типов сальмонелл при росте на плотной питательной среде характерно образование слизистого вала (феномен валообразования) вокруг колоний. У многих типов сальмонелл колонии похожи на капельки слизи. Растут бактерии на синтетических солевых средах и могут использовать цитрат как источник углерода. Многие штаммы газообразующие, сбраживают глюкозу при 37 °C, не сбраживают лактозы, сахарозы, не образуют индола, не разлагают мочевины, не образуют ацетилметилкарбинола, дают каталазоположительную и оксидазоотрицательную реакции, не разжижают желатина.
Сальмонеллы хорошо растут при pH 7,2—7,4; при более низкой активной кислотности среды (pH 5,0 и менее) они погибают.
Бактерии рода Salmonella различаются антигенной структурой. Известно около 1000 серотипов этих бактерий.
В пищевых продуктах наиболее часто встречается S. typhimurium. Температурные границы роста сальмонелл 5,2—56,5 °C. Минимальная температура роста 7 серотипов сальмонелл составляет 5,5—6,8 °C.
Активное развитие S. typhimurium наблюдалось на тушках птицы при 15 °C. С уменьшением температуры активность роста бактерий снижалась: при 10; 8,8; 7,7 и 6,6 °C время до появления роста увеличивалось соответственно до 1, 2, 3 и 4 сут (Носкова, 1972).
Сальмонеллы являются основным возбудителем пищевых отравлений. При этом более 50% всех пищевых заболеваний возникает вследствие потребления обсемененных мяса и мясных продуктов. Отравление человека продуктами, обсемененными сальмонеллами, наблюдается, если в 1 г содержится 105—106 бактерий. При таком содержании бактерий внешний вид, запах, вкус продукта не изменяются. В продукт бактерии рода Salmonella попадают при недостаточно тщательном ветеринарно-санитарном контроле производства. Так, туши животных, обсемененные сальмонеллами, если они вовремя не были отбракованы, могут явиться источником заражения туш, благополучных в санитарном отношении. В готовом продукте сальмонеллы обнаруживаются в случае недостаточной тепловой обработки.
При обработке продуктов (варка мяса, пастеризация молока) при температурах, применяемых в промышленности, сальмонеллы погибают. Мясо, обсемененное сальмонеллами, для полной гарантии его обезвреживания необходимо подвергать тепловой обработке с таким расчетом, чтобы температура в середине (центре) куска в течение 10 мин держалась на уровне 80 °C.
Для того чтобы исключить развитие сальмонелл при холодильном хранении пищевых продуктов, температура хранения должна быть ниже минимальной температуры их роста.
Сальмонеллы довольно устойчивы к воздействию низких температур. Имеются сообщения, что они сохранялись в почве и кале животных весь зимний период. Сальмонеллы, содержащиеся в пищевых продуктах, хорошо переносят замораживание. Так, в замороженном мясном фарше количество бактерий S. typhimurium через 3 мес хранения при —20 °C составляло около 50 % от начального. В готовых быстрозамороженных тефтелях, специально инокулированных сальмонеллами, последние обнаруживались через 1,5 года хранения при — 18°С.
Высокую устойчивость сальмонелл к воздействию различных факторов необходимо учитывать при разработке новых видов упаковок для пищевых продуктов, дополнительных к холоду средств, которые, подавляя развитие психротрофиой микрофлоры, могут привести к относительному увеличению количества сальмонелл.
Бактерии рода Staphylococcus — это грамположительные кокки диаметром 0,5—1,5 мкм (рис. 26). В результате характерного деления клеток (более чем в одной плоскости) образуются гроздевидные скопления. На твердых питательных средах бактерии растут в виде золотистых, лимонно-желтых и белых колоний. Стафилококки относятся к факультативным анаэробам, однако они лучше развиваются в аэробных условиях. Бактерии сбраживают глюкозу с образованием молочной кислоты в аэробных условиях и уксусной кислоты в анаэробных. Оптимальная температура их роста 37 °C; максимальная 45 °C; минимальная около 7 °C. Стафилококки довольно устойчивы к низким (отрицательным) температурам.
Рис. 26. Staphylococcus
Бактерии рода Staphylococcus гидролизуют различные белковые и жиросодержащие субстраты. Большинство штаммов солеустойчивы (развиваются при содержании в среде 7,5—12% хлорида натрия), растут как в кислых, так и щелочных средах (pH 4,2—9,3, оптимум pH 7,0—7,5). По отношению стафилококков к температуре, выше максимальной температуры их роста, существуют разноречивые мнения. Имеются данные (Минор, Март, 1980), что популяция от 106 до 107 клеток может быть полностью уничтожена за 4—24 мин при 54—60 °C; в то же время сообщается, что для гибели стафилококков необходимы нагревание среды, в которой они находятся, до 80 °C и последующая выдержка при этой температуре в течение 10—40 мин. Степень воздействия высоких температур на стафилококки зависит от реакции питательной среды и возраста культуры.
Типичным представителем рода Staphylococcus является S. aureus. По данным Минора и Марта, замораживание клеток S. aureus в триптиказо-соевом бульоне и последующее хранение при —30 °C в течение 24 ч незначительно влияло на их выживаемость. По нашим данным, выживаемость S. aureus штамм 209 при замораживании в жидком азоте составляла 95%, на воздухе при —30 °C—98%; при —18 °C —90,7% и при —10 °C— 78%. При хранении культур стафилококков в мясном фарше в течение 4 мес при. —18 °C бактерии полностью не погибали.
Бактерии S. aureus способны образовывать токсин, вызывающий пищевые отравления. По данным В. Hobbs (1974), отравление человека наступает, когда в 1 г продукта содержится 500 тыс. и более активно растущих токсигенных стафилококков. При этом токсин накапливается в продукте. Такое отравление определяют как токсикоинфекцию. Токсигенные стафилококки, содержащиеся в 1 г продукта в количестве 500 тыс., могут развиваться в желудке человека, выделяя токсин. В этом случае отравление определяют как интоксикацию.
S. aureus являются коагулазоположительными микроорганизмами. До недавнего времени многие исследователи считали, что способность образовывать энтеротоксин характерна лишь для коагулазоположительных штаммов. В настоящее время установлено1, что некоторые коагулазоотрицательные стафилококки, выделенные из пищи при вспышках стафилококковых отравлений, продуцировали энтеротоксин (Минор, Март, 1980).
Стафилококки очень широко распространены в природе. Обычным местом обитания энтеропатогенных стафилококков являются слизистые оболочки теплокровных животных, носоглотка человека, кожный покров, гнойничковые раны на руках. Из этих источников бактерии могут попадать в продукт при его выработке, если нарушены санитарно-гигиенические требования производства. Стафилококки встречаются в мясных и молочных продуктах, в мороженом, о чем более подробно будет изложено в соответствующих главах.
Клетки бактерий рода Bacillus палочковидные, прямые, размером 0,3—2,2X1,2—7,0 мкм (рис. 27). Большинство бактерий этого рода подвижны. В процессе развития они образуют термоустойчивые эндоспоры (не более одной в клетке). Бактерии грамположительные, строгие аэробы или факультативные анаэробы.
Микроорганизмы рода Bacillus широко распространены в природе (в воде, воздухе, почве). Пищевые продукты являются для них хорошей питательной средой. Они могут вызывать порчу продукта, особенно подвергнутого тепловой обработке, в процессе его последующего длительного охлаждения. Наиболее часто в мясных и молочных продуктах встречаются следующие виды: В. subtilis, В. mycoides, В. mesentericus, В. megatherium, В. cereus. Все эти бактерии, кроме В. cereus, сапрофиты. Характерной особенностью бактерий рода Bacillus является способность активно разлагать белок, поэтому в процессе их развития происходит гнилостная порча мясных продуктов и пептонизация молока. Максимальная температура развития бактерий рода Bacillus 45 °C, оптимальная 25—37 °C, минимальная 10 °C.
Рис. 28. Cl. perfringens
Бактерии рода Bacillus хорошо развиваются в нейтральной или щелочной среде (pH 7—9,5). В кислой среде процессы жизнедеятельности микроорганизмов замедляются. Например, В. mesentericus при pH 5 не развиваются.
В. cereus имеют широкие температурные границы роста (6—55°C). Они хорошо развиваются при pH от 4 до 12,5, а также в средах с содержанием соли до 10%. В связи с большой устойчивостью к различным факторам внешней среды споры В. cereus могут встречаться в пищевых продуктах, прошедших тепловую обработку (пастеризованное молоко, колбасы, готовые кулинарные изделия), и прорастать при благоприятных для них условиях. Поэтому продукты, подвергнутые тепловой обработке, необходимо быстро охлаждать до температуры не выше 5 °C, исключающей развитие этих бактерий.
В. cereus могут вызывать пищевые интоксикации, в случае если в 1 г продукта содержится 106—107 клеток.
Бактерии рода Clostridium (рис. 28)—это крупные, споровые, подвижные (иногда неподвижные) грамположительные палочки размером 0,8—1,5X4,0—8,0 мкм; анаэробные, сульфитредуцирующие.
Типичным представителем этого рода является С. perfringens. Оптимальная температура роста 37—45 °C, минимальная 6,5 °C. Кислая реакция среды замедляет развитие бактерий. В пищевых продуктах размножается при 18—20 °C, выделяя токсины, которые вызывают пищевые отравления. Образующиеся токсины не изменяют органолептических свойств пищевых продуктов (исключение составляет молоко). Пищевые отравления наступают при потреблении продукта, в 1 г которого содержится 106 бактерий. Наиболее нежелательно наличие бактерий в продукте, который после тепловой обработки охлаждают продолжительное время в диапазоне температур от 45 до 18 °C. Охлаждение продукта следует проводить в таком режиме, чтобы как можно быстрее его температура стала ниже минимальной температуры развития С. perfringens.
Плесневые грибы и дрожжи. Оптимальная температура роста плесневых грибов и дрожжей составляет 20—25 °C. Следовательно, они могут хорошо развиваться в области температур, характерных для роста мезофильных микроорганизмов. При этих температурах активизируются их рост и метаболические процессы, в частности способность к протеолизу и липолизу у плесневых грибов, и к сбраживанию сахаров и липолизу у дрожжей. У грибов, способных продуцировать токсины, этот процесс также активизируется. Например, в мясе, содержащем токсигенные штаммы плесневых грибов, при 15, 20 и 30 °C образовывались афлотоксины. Особенно это свойственно для Aspergillus при развитии на молочных продуктах (сыр, масло).
Адаптация микроорганизмов к низким температурам
Механизм адаптации психротрофных микроорганизмов к низким температурам до настоящего времени полностью не раскрыт. Главной специфической чертой психротрофных микроорганизмов является их способность активно расти при температурах ниже 10 °C. Более высокую скорость размножения при этих температурах имеют облигатные психрофилы по сравнению с психротрофами (Лях, 1976; Чистяков, 1959).
При понижении температуры у психротрофных бактерий наблюдается интенсивный перенос питательных веществ внутрь клетки, увеличивается образование экстрацеллюлярных энзимов, с помощью которых может происходить более активное расщепление субстрата и поступление его составных частей в клетку.
Некоторые исследователи считают, что психротроф- ные микроорганизмы, приспосабливаясь к росту при низких температурах, синтезируют увеличенное количество ферментов.
Способность микроорганизмов расти при низких температурах объясняют строением клеточной мембраны и ее липидным составом. Бактериальная мембрана — это система, состоящая из цитоплазматической, охватывающей всю клетку, и внутриклеточных мембран, которые являются выростами цитоплазматической мембраны. Клетки грамотрицательных бактерий имеют еще одну особенность в строении — внешнюю мембрану, представляющую собой гликолипротеидную часть клеточной стенки. В процессе роста культуры психрофильных бактерий, выделенных из вод субарктической зоны, образуется дополнительный слой клеточной стенки. Толщина клеточной стенки при этом изменяется, появляются микрокапсулы и слизь. Имеются сообщения, что в липидах мембран клеток психротрофных микроорганизмов содержится больше ненасыщенных жирных кислот, чем в липидах мембран клеток мезофилов (Иннис, Ингрем, 1981; Schmidt-Lorenz, 1972).
Нашими исследованиями установлено, что с понижением температуры выращивания Ps. fluorescens штамм 146 происходит изменение жирнокислотного состава липидов мембраны клетки. При этом увеличивается общее количество ненасыщенных жирных кислот, в частности, происходит накопление отдельных жирных кислот с четным числом углеродных атомов (табл. 5).
Таблица 5
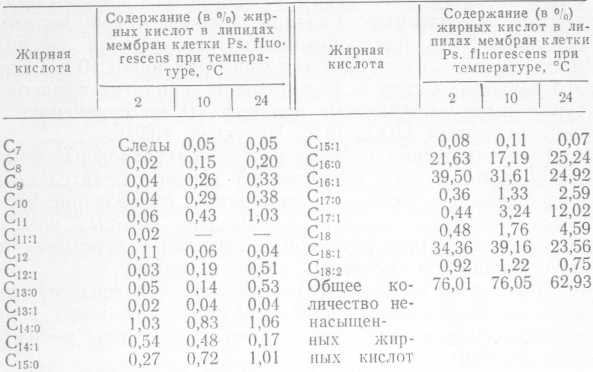
Эти изменения влияют На физическое состояние мембраны, в частности увеличивается степень разжиженности липидного матрикса. Перестройка в липидном компоненте мембраны, по-видимому, вызывает изменение в направленности действия ферментов и позволяет бактериальной клетке перейти на новый метаболический уровень.
Предполагают, что при адаптации психротрофных бактерий к низким температурам происходит изменение процесса дыхания микроорганизмов.
Для дыхания психротрофных бактерий, которые являются аэробами, необходимо наличие кислорода. Они обладают особым аппаратом — дыхательной цепью, в которой значительная часть свободной энергии клетки переводится в биологически доступную форму. Компонентами дыхательной цепи являются дыхательные ферменты, сосредоточенные в цитоплазматической мембране. Они катализируют посредством цитохромов процесс переноса электронов от окисляемого субстрата к восстанавливаемому кислороду (Фробишер, 1965).
Зависимость активности эндогенного дыхания от температуры у психротрофных бактерий Ps. fluorescens была изучена в лаборатории микробиологических исследований ВНИКТИхолодпрома Г. А. Баландиной и автором совместно с сотрудниками Института биохимии АН СССР Д. Н. Островским и А. С. Капрельянцем. Установлено, что активность эндогенного дыхания психротрофных бактерий Ps. fluorescens штамм 146 в логарифмической фазе роста при 24 и 10 °C была одинаковой и составляла 0,040 мкмоль О2/мин на 1 мг сухой массы.
Температура роста, °C
24 10 52
0,04 0,04 0,085 0,07
Средняя активность эндогенного дыхания мкмоль О2/мин на 1 мг сухой массы
При понижении температуры выращивания бактерий До 5 и 2 °C активность их дыхания увеличивалась и была равна соответственно 0,085 и 0,070 мкмоль О2/мин на 1 мг сухой массы.
Полученные данные позволяют сделать вывод, что активность дыхания психротрофных бактерий Ps. fluorescens возрастает с понижением температуры их культивирования при неизменном содержании цитохромов в 1 мг клеточного белка.
Изменение жирнокислотного состава липидов мембраны и повышение активности дыхания клетки позволяют ей адаптироваться к понижению температуры выращивания.
Влияние замораживания на микроорганизмы
В настоящее время широко используется способ хранения мяса, мясных и молочных продуктов в замороженном виде, поэтому важно знать, какие изменения происходят с микроорганизмами при замораживании и хранении продуктов. Наибольшее распространение получили следующие температуры замораживания и хранения: -12-4—18-4—23 °C. В последнее десятилетие наметилась тенденция к замораживанию пищевых продуктов при очень низких температурах —30-4—50 °C, а также в среде диоксида углерода (—78 °C) или жидкого азота (—196°C).
При выборе обоснованных санитарно-гигиенических требований к процессам замораживания и размораживания продуктов, а также последующей выдержки их в размороженном состоянии необходимо учитывать степень устойчивости микроорганизмов к замораживанию.
Исследованиями многих авторов установлено, что устойчивость микроорганизмов к замораживанию зависит от ряда факторов: температуры и скорости замораживания; состава среды, в которой происходит замораживание; кратности замораживания; вида микроорганизма и его физиологического состояния (Смит, 1963; Носкова, 1960, 1979).
Гибель микроорганизмов при замораживании в основном обусловлена температурой и скоростью замораживания (табл. 6, данные О. Смита, 1963).
Таблица 6
|
Изменение температуры |
Скорость изменения температуры |
Температура, °C |
|
Сверхбыстрое |
2 с и менее |
От 0 до —190 |
|
Быстрое |
От 2 с до 2 мин |
От 0 до —79 |
|
Медленное |
10 мин и более |
От 0 до —79 |
|
Очень медленное |
1 ч и более |
-20 |
При замораживании среды в ней появляются кристаллы льда, размер которых зависит от скорости изменения температуры. Так, при медленном замораживании, начиная с температуры —2-.—3 °C, постепенно вне клеток образуются крупные кристаллы иглообразной формы с четкими гранями. Кристаллы прокалывают клеточную оболочку бактерии или сдавливают ее, целостность бактериальной клетки нарушается.
При быстром замораживании образуются очень мелкие вне- и внутриклеточные кристаллы, которые в значительно меньшей степени повреждают бактериальные клетки.
Считают, что при сверхбыстром замораживании вода внутри клеток и в межклеточном пространстве превращается в лед, минуя стадию кристаллизации (явление витрификации). При этом степень повреждения бактериальной клетки снижается. Данные, полученные с помощью рентгеноструктурного анализа и метода электронной микроскопии, свидетельствуют о наличии мельчайших кристаллов в биологических объектах после сверхбыстрого замораживания (Никитин, Звягин, 1971).
Влияние замораживания было изучено еще в 1884 г. Пикте, который установил, что бактерии различных видов, замороженные в жидком воздухе и выдержанные затем при температуре —70 °C в течение 3 сут и при — 120 °C в течение 2,5 сут, оставались жизнеспособными после размораживания.
Дальнейшие исследования влияния низких температур на различные виды патогенных бактерий также показали, что они остаются жизнеспособными после выдержки в течение 20 ч при температурах —182-4—190 °C.
Гибель бактериальных клеток при замораживании происходит не только в результате механического разрушения ее оболочки, но и вследствие повышенной концентрации солей в среде. Например, при замораживании бактериальных клеток в питательной среде, содержащей различные соли, сначала замерзает вода, концентрация солей в растворе повышается и достигает максимальных значений. В этих условиях происходит денатурация белков и липопротеидов бактериальной клетки, приводящая к ее гибели.
По мнению ряда авторов (Никитин, 1971), степень выживаемости бактерий при замораживании зависит от состава среды, в которой замораживаются бактерии, и наличия в ней защитных веществ. Так, при замораживании культуры сальмонеллы в 0,85%-ном растворе NaCl при —73 °C погибало 87—98% бактериальных клеток, в то время как при замораживании в растворе желатозы только 2,5—34%.
В качестве защитных веществ при замораживании бактериальных суспензий, в случае необходимости сохранения большего числа жизнеспособных клеток, используют глицерин, сахарозу, желатин, сывороточные белки, углеводы, обезжиренное молоко. Например, замораживание бактерий Е. coli при —20 °C в воде и выдерживание их при этой температуре в течение 5 сут показало, что жизнеспособными оставались менее 1 % бактерий. При замораживании этих же бактерий в молоке при тех же условиях процент выживших клеток значительно повышался, а при разбавлении молока водой снова снижался.
Существенное значение для сохранения жизнеспособности микроорганизмов имеет кратность замораживания и размораживания. Установлено, что каждое повторное замораживание и размораживание суспензии бактериальных клеток ускоряет их гибель. Так, при однократном замораживании бактерий Е. coli при —78 °C выживало 84% клеток, при десятикратном — 10 и пятнадцатикратном— 6% (Смит, 1963).
Устойчивость микроорганизмов к замораживанию зависит от возраста бактериальной клетки. Наибольшее количество бактерий погибает, если процесс замораживания совпадает с логарифмической фазой их роста.
Для восстановления жизненных функций бактерий после замораживания большое значение имеет состав питательной среды и условия культивирования бактерий в размороженном состоянии.
Влияние замораживания на психротрофные бактерии изучалось в лаборатории микробиологии ВНИКТИхолодпрома на двух видах (Ps. fluorescens штамм 146 и Ps. fragi штамм 237), выделенных из охлажденного мяса. Бактерии замораживали в мясном фарше и синтетической среде (Arpai, 1965) следующего состава: К2НРО4 — 2 г; NH4H2PO4 —2 г; (NH4)2SO4 —2 г; MgSO4-7H2O— 0,2 г; цитрат натрия 0,5 г; СаС12 — 0,01 г; FeSO4-7H2O — 0,005 г, вода дистиллированная 1000 мл; pH 7,2. Замораживание осуществляли на воздухе при — 18 °C и в жидком азоте при —196 °C, а размораживание на воздухе при 20 °C в течение 1,5 ч (медленное размораживание) и в воде при 30 °C в течение 15 мин (быстрое размораживание). Начальное количество бактерий в 1 мл среды перед замораживанием составляло 107 (табл. 7).
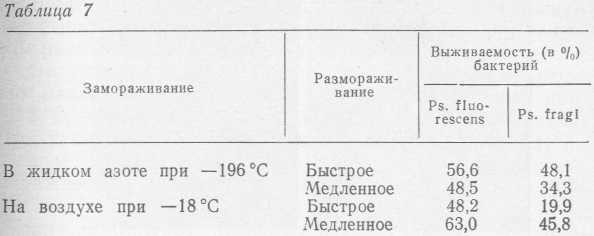
Аналогичные результаты были получены при замораживании бактерий в мясном фарше.
В целом, в результате замораживания как на воздухе, так и в жидком азоте, погибает от 37 до 80% психротрофных бактерий. Аналогичные данные, характеризующие выживаемость бактерий, были получены Г. А. Баландиной при электронно-микроскопическом исследовании морфологических изменений в клетке бактерий Ps. fluorescens штамм 146. Так, при быстром размораживании культуры (5 мин при 30°C), замороженной при —196 °C, клетки теряли свою типичную палочковидную форму. На поверхности бактерий встречались осмиофильные конгломераты, по своей электронно-оптической плотности близкие к плотности цитоплазмы (рис. 29, а). При медленном размораживании (60 мин при 20 °C) наблюдалась сильная дезагрегация всех составных частей клетки; клеточная стенка бактерий была разорвана и содержимое клетки переходило во внешнюю среду (рис. 29,6).
При быстром размораживании культуры, замороженной при —18 °C, клетки принимали неправильную форму, выглядели размытыми и были похожи на клетки, которые подвергали быстрому замораживанию и медленному размораживанию (рис. 29, в).
При медленном размораживании культуры, замороженной при —18 °C, отмечалось отхождение клеточной стенки от протопласта. Иногда в поле зрения микроскопа встречались клетки с разрушенной клеточной стенкой (рис. 29,г).
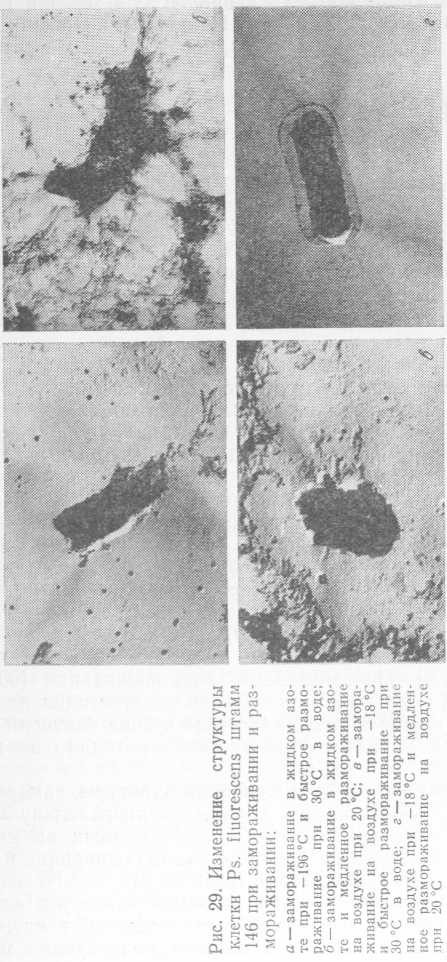
Таким образом, наиболее существенные деструктивные изменения клеток Ps. fluorescens штамм 146, приводящие к гибели бактерий, были отмечены при резком перепаде температур, т. е. при медленном размораживании быстрозамороженных бактериальных клеток и быстром размораживании медленнозамороженных клеток.
Полученные результаты позволяют сделать вывод, что мясные и молочные продукты, замороженные при — 18 °C, следует подвергать быстрому размораживанию. При таком режиме большинство бактерий погибает, и, следовательно, степень порчи продукта в процессе размораживания и последующего кратковременного хранения в условиях реализации уменьшается.
При длительном хранении психротрофных бактерий в замороженном состоянии в питательной среде происходит их постепенное отмирание (рис. 30). Так, наибольшее количество бактерий погибает в первые месяцы хранения, однако и после 12 мес хранения полной гибели не происходит. Следовательно, можно сделать вывод, что замораживание не является стерилизующим фактором.
Бактерии, сохранившиеся после замораживания и длительного хранения в замороженном состоянии, способны размножаться после размораживания (рис. 31).
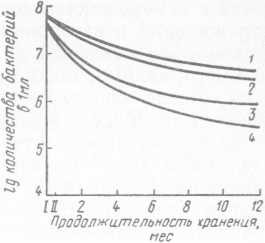
Рис. 30. Динамика отмирания бактерий Ps. fluorescens штамм 146 (а) и Ps. fragi штамм 237 (б) в процессе замораживания и хранения при —18 °C:
Ь 3 — замораживание соответственно в воздушной среде и жидком азоте, быстрое размораживание; 2, 4 — замораживание соответственно в воздушной среде и жидком азоте, медленное размораживание; 1 — RO замораживания; II — после замораживания
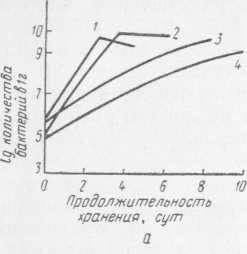
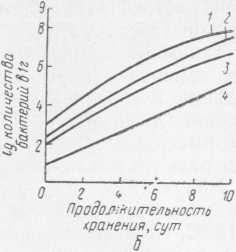
Рис. 31. Динамика роста бактерий: а — Ps. fragi штамм 237; 1, 3 — рост незамороженной культуры соответственно при 24 и О °C (контроль); 2, 4 — рост культуры, замороженной в жидком азоте, после размораживания соответственно при 24 и О °C; б — Ps. fluorescens штамм 146; 1 — рост незамороженной культуры при О °C (контроль); 2, 3, 4 — рост размороженной культуры при О °C после хранения при —18 °C соответственно в течение 1, 2, 5 мес
Как видно из рис. 31, замораживание и хранение не влияют на продолжительность генерации бактерий Ps. fragi штамм 237 и Ps. fluorescens штамм 146 (она была одинаковой как у незамороженной культуры, так и у бактерий после замораживания и длительного хранения при —18°C). Более того, отмечены отдельные случаи, когда бактерии Ps. fluorescens через 1 мес хранения в замороженном состоянии при —18 °C после размораживания и выдержки при 24 °C развивались более активно, чем бактерии, не подвергавшиеся замораживанию. Это, видимо, можно объяснить тем, что под действием холода происходит как бы искусственный отбор более устойчивых и жизнеспособных клеток.
Замораживание бактерий при —196 и —18 °C и последующее хранение при —18 °C в. течение 12 мес не влияет на культурально-биохимические свойства психро- трофных бактерий Ps. fluorescens. В частности, одно из практически важных свойств этих бактерий при холодильном хранении мяса и молока — протеолитическая активность — не изменялось при замораживании.
Психротрофные бактерии, перенесшие замораживание, развиваются на размороженных пищевых продуктах и могут ухудшать их качество. Это следует иметь в виду при размораживании мяса, мясных продуктов, молока, творога.
Наиболее часто в мясных и молочных продуктах из мезофильных бактерий встречаются стафилококки, энтерококки, протей. Эти микроорганизмы имеют санитарно-гигиеническое значение.
Влияние замораживания на стафилококки изучали на токсигенном штамме 209. Чистую 24-часовую культуру вносили в стерильное молоко и замораживали в жидком азоте при температуре —196 °C и на воздухе при температуре —30, — 18 и —10 °C (табл. 8). Стафилококки характеризовались высокой выживаемостью. С повышением температуры замораживания от —196 до —10 °C количество погибших клеток увеличивалось. При хранении замороженных в молоке бактерий в течение 4 мес постепенное отмирание их происходило при температурах —196, —30 и —18 °C и резкое — при -10°С.
Таблица 8

Имеются сообщения, что стафилококки не погибали в течение 6 мес при —12 и —18 °C. При этом сохранялся и токсин, выделенный бактериями до замораживания.
Стафилококки, замороженные в молоке, при последующей выдержке при 37 и 10 °C размножались примерно так же, как и не подвергшиеся замораживанию (рис. 32, а). Количество стафилококков при выдержке размороженной культуры в течение 3—5 сут при 10 °C достигало 5хЮ5 в 1 мл продукта. Длительное хранение замороженной культуры стафилококков существенно не влияло на рост бактерий после размораживания — продолжительность генерации была примерно такой же, как у незамороженных культур бактерий.
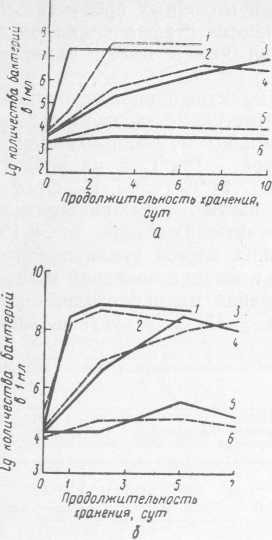
Рис. 32. Динамика развития S. aureus (штамм 209 (а), Str. faecalis штамм 489 (б), Proteus vulgaris (в) после замораживания в жидком азоте и последующего хранения в размороженном состоянии при различных температурах: 1, 2 — 37 °C; 3, 4 — 10 °C; 5, 6 — 5 °C
При замораживании и хранении культур S. aureus в течение 4 мес сохранялись их токсигенные свойства, что подтверждалось положительной реакцией плазмокоагуляции и гемолитической активностью. При оптимальной температуре роста этих бактерий гемолиз эритроцитов кролика был явно выражен в разведениях 1 : 50, 1 : 100 и 1 : 200. При 10 °C как у незамороженных, так и у культур, подвергавшихся замораживанию, токсигенные свойства были выражены очень слабо, о чем свидетельствуют нечеткие реакции плазмокоагуляции и гемолиза. Хотя при 10 °C накопление токсина в продукте замедлялось, однако наличие стафилококков в продуктах, хранящихся при 10 °C, небезопасно для здоровья человека.
Развитие стафилококков не вызывает каких-либо визуальных изменений, по которым можно было бы судить о непригодности продукта к употреблению. В связи с особой устойчивостью стафилококков к замораживанию следует осуществлять строгий санитарно-гигиенический контроль при производстве продуктов, вырабатываемых с использованием холода и поступающих к потребителю в готовом виде — мороженое, готовые быстрозамороженные блюда (подвергающиеся лишь разогреванию перед употреблением).
Довольно часто в пищевых продуктах и особенно замороженных встречаются фекальные стрептококки. Влияние замораживания на фекальные стрептококки изучали на культуре Str. faecalis штамм 489, внесенной в стерильное молоко. Степень выживаемости этих бактерий при —18 и —196 °C и последующем хранении при — 18 °C в течение 6 мес представлены в табл. 9.


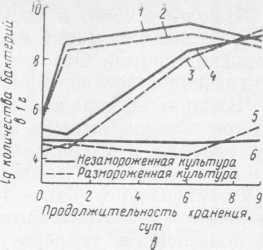
влияло на развитие бактерий после размораживания (см. рис. 32,6). Бактерии размножались, минуя лаг-фазу; продолжительность генерации в логарифмической фазе роста при оптимальной температуре развития 37 °C составляла 1,8 ч и при минимальной 10 °C — 8,7 ч. Биохимические свойства, характерные для Str. faecalis (восстановление лакмусового молока при 45 °C, способность к росту в мясопептонном бульоне (МПБ) с pH 9,6 и в МПБ с содержанием 6,5% NaCl, ферментация углеводов после замораживания и хранения в замороженном состоянии), не изменялись. Что касается термоустойчивости Str. faecalis, то бактерии, подвергнутые нагреванию при 63 °C в течение 30 мин непосредственно после замораживания, погибали. Однако после 7-суточной выдержки размороженных культур в молоке при оптимальной температуре (37 °C) устойчивость их к нагреванию восстанавливалась. Эти бактерии, благодаря их большой устойчивости к замораживанию, могут быть использованы как санитарно-гигиенический показатель условий производства быстрозамороженных готовых блюд и мороженого, тем более что они довольно часто обнаруживаются в этих продуктах.
Влияние замораживания на бактерии рода Proteus изучали на штамме Proteus vulgaris, наиболее часто встречающемся в мясе и мясных продуктах. Бактерии вносили в стерильный мясной фарш, который затем замораживали. Данные о выживаемости бактерий протея при —18 и —196 °C и последующем хранении при —18 °C в течение 4 мес представлены в табл. 10.
Замораживание не влияло на характер развития бактерий Proteus vulgaris после размораживания (см. рис. 32, в).
После 4 мес хранения клетки, оставшиеся жизнеспособными, размножались при 37 и 10 °C, так же, как и бактериальные клетки, не подвергавшиеся замораживанию, о чем свидетельствует продолжительность генерации этих бактерий: 2 ч при 37 °C и 8,2 ч при 10 °C у незамороженной культуры бактерий и 1,9 ч и 7,8 ч соответственно после замораживания и последующей выдержки в размороженном виде при указанных выше температурах. При 5 °C лаг-фаза у испытуемого штамма Proteus vulgaris длилась почти 6 сут, после чего наблюдалось незначительное развитие.
Замораживание не влияло и на биохимические свойства бактерий протея, в частности на протеолитические. Поэтому мясо и мясные продукты, обсемененные микроорганизмами при размораживании и особенно при вынужденном последующем хранении при температуре выше минимальной температуры развития протея, могут быстро портиться.
Таблица 11

Молочнокислые бактерии — стрептококки и палочки—довольно хорошо переносят замораживание. Данные о выживаемости отдельных видов бактерий в процессе хранения при —25 °C представлены в табл. И (Банникова, 1975). Через 6 мес физиолого-биохимические свойства выживших культур не изменялись. Способ замораживания чистых культур и их сочетаний с целью длительного сохранения используется в нашей стране и за рубежом.
Сопоставляя степень выживаемости мезофильных бактерий кокковой и палочковидной формы, можно видеть, что первые более устойчивы при замораживании и холодильном хранении. Это обусловлено специфическим строением клеточной оболочки кокковых форм бактерий.
