В пиве может развиваться только ограниченное число видов микроорганизмов, среди которых отсутствуют патогенные для человека и спорообразующие штаммы. Современная классификация микроорганизмов пива может быть представлена следующим образом [136]:
грамположительные бактерии — каталазоотрицательные палочки — Lactobacillus; каталазоотрицательные кокки или клетки, собранные в пакеты,— Pediococ- cus;
грамотрицательные бактерии — уксуснокислые бактерии— Acetomonas и Acetobacter; бактерия Obesum- bacterium proteus; бактерия Zymomonas anaerobia; бактерии группы кишечной палочки — Klebsiella и Escherichia;
дикие дрожжи.
Грамположительные бактерии. Встречающиеся в пиве грамположительные бактерии, относящиеся к родам Lactobacillus и Pediococcus, часто объединяются под названием молочнокислых бактерий.
Как указывает Шимвэлл [208], термин «истинные молочнокислые бактерии» относится только к определенной группе бактерий, которые не только образуют молочную кислоту в качестве основного продукта превращения углеводов, но и обладают другими характерными свойствами. Все они грамположительны, неподвижны, не образуют спор, каталазоотрицательны и требуют в различной степени для роста сложную смесь продуктов распада белка и факторов роста. Автор [208] предлагает разделить группу молочнокислых бактерий пива на два рода: Lactobacillus и Streptococcus (вместо названия Pediococcus). В литературе часто встречаются оба названия.
В качестве источников азота молочнокислые бактерии требуют целого набора аминокислот, сильно сбраживают сахара, на белки действуют слабо, жиры почти не атакуют. Дыхательная способность их ограниченна, поэтому рост их усиливается в анаэробных или микро- аэрофильных условиях. Некоторые штаммы обладают способностью усваивать СО2, поэтому в атмосфере этого газа отмечается особенно сильный рост. Оптимум pH для роста — примерно 5,5, но некоторые штаммы могут расти при pH 3,5.
Бактерии рода Pediococcus и некоторые штаммы Lactobacillus являются гомоферментативными бактериями, т. е. образуют из сахара в основном молочную кислоту. Другие штаммы Lactobacillus, например наиболее часто встречающийся в пиве вид L. pastorianus, являются гетероферментативными бактериями, образующими из сахара наряду с молочной кислотой уксусную кислоту, глицерин, этанол и СОг.
В пиве встречаются следующие виды Lactobacillus: L. pastorianus, L. pastorianus var. brownii, L. diastati- cus, L. lindneri, L. malefermentans, L. fridigus и L. parvus. L. lindneri и L. fridigus имеют более низкий оптимум температуры (19° С) для роста, чем L. pastorianus, и встречаются почти исключительно при низовом брожении. L. malefermentans и L. parvus отличаются от L. pastorianus по набору сахаров, которые поддерживают рост. L. diastaticus особенно устойчив к этанолу и хмелевым смолам и характеризуется также способностью гидролизовать декстрины и крахмал.
Гетероферментативные штаммы обычно имеют вид длинных палочковидных клеток, однако часто наряду с длинными могут встречаться и короткие палочки, единичные или соединенные в пары. Размеры клеток 1,ОХ (5-4-120) мкм.
Хмелевые смолы и изомеризованные продукты в общем ингибируют рост молочнокислых бактерий, однако виды, обычно встречающиеся в пиве, в значительной мере утратили свою чувствительность к этим ингибиторам, очевидно, приспособившись к условиям, существующим в пиве. Было показано [136], что устойчивость к гумулону и его производным быстро создается у ряда штаммов и достаточно 2—5 пассажей, чтобы резистентность к этим веществам в концентрации 100 мг/мл повысилась в 8—20 раз.
Многие молочнокислые бактерии способны образовывать в пиве диацетил. Запах диацетила чувствуется уже в концентрации 0,0025 мкг/мл (раньше образование больших количеств диацетила называли «сарцин- ной болезнью» пива; считалось, что это заболевание вызывается определенной сарциной — Pediococcus по современной номенклатуре).
Как гомоферментативные, так и гетероферментатив- ные молочнокислые бактерии являются ауксотрофными по ряду специфических аминокислот и часто не способны расти при отсутствии хотя бы одной из них. К числу таких необходимых для роста аминокислот относятся аргинин, аспарагиновая и глютаминовая кислоты, изолейцин, лейцин, фенилаланин, пролин, серин, треонин, триптофан и валин. Кроме того, некоторые штаммы требуют для роста аланин, глицин, гистидин, лизин, метионин и тирозин, т. е. почти все аминокислоты необходимы для роста молочнокислых бактерий. Различные молочнокислые бактерии требуют определенных витаминов и факторов роста, таких, как р-аминобен- зойная и фолиевая кислоты, а также некоторые пурины и пиримидины.
Количество образующейся молочной кислоты зависит от концентрации углеводов, ассимилируемых молочнокислыми бактериями. Биохимические пути превращения углеводов у молочнокислых бактерий разнообразны. Гомоферментативные штаммы, такие, как Pediococcus, используют гликолитический путь, приводящий к образованию пировиноградной кислоты, которая, действуя как акцептор водорода, превращается в молочную кислоту. Этот процесс катализируется дегидрогеназой молочной кислоты. Полагают, что помимо этого пути гомоферментативные штаммы используют гексозомонофосфатный и, возможно, фосфокетолазный путь в том случае, когда развиваются на пентозах (рис. 1).
Гетероферментативные штаммы не имеют ферментов альдолазы и гексозоизомеразы, необходимых для осуществления гликолитического механизма, поэтому превращение гексоз и пентоз этими штаммами происходит По .фосфокетбл'азйому пути. Ключевой реакцией е этом' механизме, является расщепление пентозо-5-фос- фата до ацетилфосфата й глицеринового альдегида, катализируемое фосфокетолазой. Фруктоза является лучшим, чем глюкоза, ’ субстратом для роста гетеро-
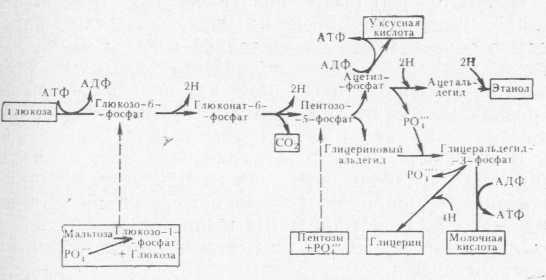
Рис. 1. Фосфокетолазный путь превращения углеводов.
ферментативных штаммов и в том случае, когда присутствуют оба сахара, действует как акцептор водорода и восстанавливается до маннита.
Мальтоза также легко используется бактериями, причем имеются штаммы, которые, потребляя мальтозу, не могут использовать глюкозу [254]. Следовательно, превращение мальтозы идет не по пути ее гидролитического расщепления на две молекулы глюкозы. Установлено, что у этих штаммов отсутствует фермент мальтаза и превращение мальтозы катализируется ферментом мальтофосфорилазой, под влиянием которого из половины молекулы мальтозы образуется р-глю- козо-1-фосфат, а вторая половина молекулы выделяется в среду в виде глюкозы:
Мальтоза + (РО4)3— (3-Глюкозе-1-фосфат + Глюкоза.
Этот путь образования фосфорилированной молекулы глюкозы имеет определенную биологическую целесообразность, поскольку в нем не происходит потребления макроэргических соединений типа АТФ. Возможно, что образование в результате реакции из мальтозы глюкозы способствует симбиотическому развитию других видов и штаммов бактерий, предпочитающих глюкозу.
Пентозы также могут служить субстратом для роста молочнокислых палочек, но после некоторого периода адаптации.
В работе [54] было показано, что три штамма Lactobacillus, выделенные из пива, обладали способностью образовывать на пептонной среде с различными углеводами небольшое количество кислоты только лишь на арабинозе, в то время как на дрожжевой воде с различными сахарами и спиртами эти бактерии образовывали кислоту на многих веществах [18].
Встречающиеся в пиве лактобациллы проявляют широкую вариабельность свойств [129].
Наиболее широко и часто встречающийся в пиве вид — это L. pastorianus. Шимвэлл [208] отмечает, что до настоящего времени биологическая стойкость пива является синонимом, его резистентности к развитию именно этого микроорганизма. L. pastorianus открыт Пастером и впервые выделен в чистую культуру и описан Ван-Лаэром в 1892 г.
Морфология микроорганизма зависит от среды и условий культивирования. Так, высокий процент спирта в пиве приводит к образованию более длинных палочек, при выращивании на сусле или сусло-агаре микроорганизм иногда растет в виде довольно коротких палочек. На поверхности твердых сред образует неправильной формы ризоидные колонии (рис. 2), не растет или почти не растет на средах животного происхождения. Максимальная температура для роста 38° С. В неохме- ленном сусле при pH 5,4 погибает через 5 мин при температуре 54° С, в пиве при pH 4,2 — через 5 мин при температуре 49° С.
Повышенное содержание хмеля задерживает рост L. pastorianus. Спирт проявляет сильное бактерицидное действие только в концентрации около 8%.
В пиве встречаются следующие виды кокковидных молочнокислых бактерий, относящихся к роду Pedio- coccus: Р. damnosus, Р. damnosus var. viscosus, P. dam- nosus var. diastaticus, P. damnosus var. limnosus, t. c. все эти виды являются вариантами одного основного вида — Р. damnosus (синонимы: Streptococcus damnosus, Р. perniciosus, Р. cerevisiae, Р. sarcinoformis). Пе- диококки встречаются главным образом в пиве низового брожения и очень редко — в пиве верхового брожения. Оптимум температуры для роста 21—25° С, однако пе- диококки способны развиваться и при более низкой температуре.
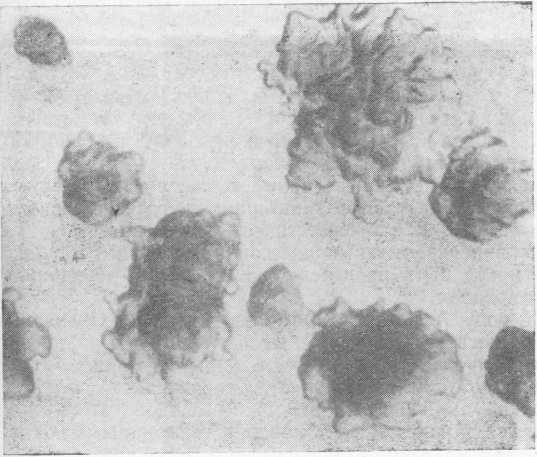
Рис. 2. Микроколонии L. pastorianus на сусло-агаре.
Кокки могут находиться в виде одиночных клеток, а также в парах, тетрадах, коротких цепочках или в виде скоплений. Разные штаммы различаются по способности превращать глюкозу, фруктозу, мальтозу и сахарозу: так, Р. damnosus var. diastaticus способен использовать декстрины и крахмал, но в отличие от других штаммов не образует диацетила. Образование значительного количества диацетила является особенно характерным признаком педиококков. Пиво при сильной инфицированности этими микроорганизмами приобретает медовый аромат («сарцинное заболевание») .
Варианты viscosus и limnosus вызывают ослизнение пива при развитии на глюкозе, фруктозе, сахарозе или мальтозе.
Все штаммы молочнокислых бактерий образуют в пиве шелковистую муть, повышают его кислотность и ухудшают вкус и аромат за счет выделения продуктов метаболизма. Помимо некоторых вариантов Р. damno- sus штаммы L. pastorianus var. brownii могут также вызвать ослизнение пива в результате образования экстрацеллюлярной слизи, которая содержит комплексный гетерополимер, состоящий из глюкозы, маннозы, белка и нуклеиновой кислоты. Полагают, что слизеобразующие штаммы случайно возникают из штаммов, лишенных этой способности, и, наоборот, из слизеобразующих штаммов в результате мутации могут появляться штаммы, не способные к образованию слизи.
Грамотрицательные бактерии. Уксуснокислые бактерии. В зависимости от интенсивности и глубины окислительных процессов уксуснокислые бактерии могут быть разделены на два рода — Acetomonas и Acetobacter.
К роду Acetomonas (прежнее название Gluconobac- ter) относятся уксуснокислые бактерии с ограниченной окислительной способностью: они окисляют этанол только до уксусной кислоты. Отличительной морфологической особенностью подвижных видов этого рода является полярное жгутикование. Бактерии имеют вид коротких палочек, единичных, в парах или цепочках, некоторые виды подвижны. Размеры клеток (0,4— 0,8) Х( 1,0—2,0) мкм.
Виды Acetomonas могут быть отнесены к глюкофилам: они хорошо растут на глюкозе, но плохо или совсем не растут на лактате, довольно требовательны к составу питательной среды для роста [209].
К роду Acetobacter принадлежат уксуснокислые бактерии с более сильной окислительной способностью: они окисляют этиловый спирт до СО2 и Н2О и могут быть отнесены к лактофилам, так как хорошо растут на лактате. Эти виды неприхотливы к составу питательной среды, и их покоящиеся культуры обладают способностью использовать глютаминовую кислоту. Морфологически они аналогичны Acetomonas, но большей частью подвижны и имеют перитрихиальное жгутикование.
Некоторые штаммы Acetobacter для превращения глюкозы используют гексозомонофосфатный путь и цикл трикарбоксильных кислот. У других штаммов перед началом этого пути может происходить окисление глюкозы до глюконата, после чего глюконокиназа образует глюконат-6-фосфат. У третьих штаммов способность к фосфорилированию полностью утеряна и глюкоза окисляется через глюконат до 2- или 5-оксоглю- коната или даже до дикетоглюконата или у-пирона.
У видов Acetomonas гексозомонофосфатный путь может функционировать, но отсутствуют некоторые ферменты цикла трикарбоксильных кислот (например, дегидрогеназа изолимонной кислоты), поэтому глюкоза обычно окисляется только до глюконовой кислоты.
Потребности в источнике азота у бактерий обоих родов уксуснокислых бактерий различны: штаммы Acetobacter могут хорошо развиваться на аммонийном азоте, штаммы Acetomonas требуют присутствия некоторых аминокислот (глютаминовой, пролина или аспарагиновой) или соответствующих кетокислот, участвующих в реакциях трансаминирования.
Уксуснокислые бактерии малочувствительны к низкому значению pH и к фитонцидному действию хмелевых смол и их изомеризованных продуктов. Эти свойства, а также неприхотливость в отношении используемых питательных веществ приводят к широкому, повсеместному распространению уксуснокислых бактерий в пивоварении. Именно эти бактерии наряду с Lactobacillus являются наиболее частой причиной изменения кислотности и вкуса пива и появления в нем мути, а иногда и пленки на поверхности пива в бутылке.
Сообщалось [136] о выделении некоторыми штаммами (например, Acetobacter rancens) токсина, убивающего дрожжевые клетки. В то же время при низкой концентрации уксуснокислых бактерий, инфицирующих пиво, отмечалось их постепенное отмирание и отсутствие роста в пиве.
Следовательно, для проявления отрицательного влияния этих бактерий на прозрачность, вкус и кислотность пива большое значение имеет величина вносимой инфекции (степень инфицированности). Уксуснокислые бактерии, так же как и молочнокислые, характеризуются большим разнообразием свойств, поэтому определение видовой принадлежности часто затруднено.
В 1912 г. в Англии был выделен штамм, вызывающий повышение вязкости пива,— Acetobacter viscosum. В последующем были обнаружены A. capsulatum и A. turbidans, дающие сильную муть в пиве в присутствии небольших количеств кислорода. Особенностью A. turbidans является то, что при температуре 13° С, когда растет он медленнее, чем при комнатной температуре, возникает более интенсивное помутнение готового продукта.
Широко распространены в задаточных дрожжах и пиве виды A. aceti, A. acetosum, A. suboxydans. A. mobile выделен из бутылочного пива, где он развивается очень быстро, вызывая помутнение и образование тонкой пленки на поверхности, позже — порошковидного осадка. Может расти в условиях ограниченного содержания кислорода и подобно A. turbidans должен считаться факультативным аэробом; в пиве образует уксусную кислоту.
Вид A. acidum-polymyxa характеризуется сильным образованием слизи при росте в сусле с pH 4,5, т. е. при пониженной кислотности, или в сусле с добавлением этилового спирта.
Уолкер и Кулка [240] поставили многочисленные эксперименты для выявления особенностей развития в пиве различных видов ускуснокислых бактерий. Они установили, что при развитии видов A. aceti, A. capsulatum, A. acidum-polymyxa, A. mobile и A. turbidans тип, вкус и аромат пива полностью портятся, оно сильно мутнеет. Некоторые виды (A. ascendes, A. acetigenus, A. viscosum), давая сильное помутнение, вкус пива не портят.
По мнению этих исследователей, наиболее опасными для пива уксуснокислыми бактериями являются виды A. acidum-polymyxa, A. mobile, A. turbidans и A. viscosum, так как они способны развиваться в пиве с низким содержанием воздуха так же легко, как и в пиве, имеющем избыток воздуха. Затем идут A. aceti и A. capsulatum.
Поскольку виды A. acetosum, A. capsulatum, A. suboxydans и A. viscosum не используют уксусную кислоту, они должны быть отнесены к роду Acetomonas.
При пастеризации погибают все штаммы уксуснокислых бактерий. Так, A. turbidans погибает при нагревании пива в течение 5 мин при 54° С или 10 мин при 52° С. Этот штамм наиболее устойчив к действию спирта (переносит концентрацию 10% спирта), для остальных штаммов содержание 6% спирта является предельным.
Бактерия Obesumbacterium proteus [210]. Этот микроорганизм (раньше его относили к роду Flavobacterium) обладает способностью развиваться конкурентно с активно почкующимися дрожжами.
О. proteus имеет вид коротких толстых палочек, неподвижных, часто соединенных в цепочки. Клетки могут варьировать по своей форме. Размеры клеток (0,8—1,2) X (1,5—4,0) мкм.
Бактерия является факультативным анаэробом с оптимумом pH 6,0, рост часто отсутствует, если pH 4,5 и ниже. Использует для своего развития различные моно- и дисахариды, крахмал не гидролизует, требует наличия многих аминокислот (аспарагиновая, глютаминовая, цистин, треонин, лейцин, аргинин, гистидин и глицин). Некоторые штаммы могут расти в присутствии только глютаминовой кислоты и аргинина. Необходимы для роста урацил и аденин. Не требуют для развития витаминов группы В.
Скорость развития бактерий зависит от интенсивности почкования культурных дрожжей. Рост бактерий происходит в первые 24 ч после начала брожения и идет особенно интенсивно в том случае, когда отмечаются длительная лаг-фаза и медленное разбражива- ние сусла дрожжами. В результате интенсивного развития микроорганизма пиво приобретает запах пастернака (запах обусловлен образованием дпметилсуль- фида).
Быстрое размножение дрожжей и падение при этом pH сбраживаемого пива подавляют развитие О. proteus. Если задаточные дрожжи сильно инфицированы О. proteus, pH пива остается высоким и уменьшения популяции бактерий так сильно, как обычно отмечается, не происходит. В этом случае бактерии могут сохраняться в разлитом пиве.
Бактерия Zymomonas anaerobia (прежнее название рода Achromobacter). Этот микроорганизм отличается сильной подвижностью. Клетки средней длины, размером (1,0—1,5) X (2,0—3,0) мкм, имеют 1—5 полярных жгутиков. Строгий анаэроб, растущий при широком диапазоне pH (3,5—7,5). Оптимум температуры 30° С, но может расти и при более низкой температуре. Использует глюкозу и фруктозу, иногда сахарозу, но не может потреблять мальтозу. Это, а также то, что для
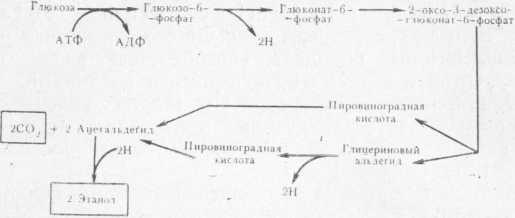
Рис. 3. Путь превращения углеводов микроорганизмами (по Энтнеру — Дудорову).
обильного роста организма требуется широкий набор аминокислот, липоевая кислота и биотин, обусловливают сравнительно редкое появление в пиве Z. anaerobia. Если же условия благоприятствуют развитию этой бактерии, она может испортить пиво в течение нескольких часов.
Бактерии группы кишечной палочки. Эти бактерии в сусле и пиве представлены рядом видов факультативных анаэробов, относящихся к родам Klebsiella (прежнее название Aerobacter) и Escherichia. Эти роды включают очень много видов, широко распространенных в почве, на растительных остатках и в воде, имеют вид коротких палочек, иногда почти шарообразной формы, по Граму окрашиваются отрицательно, спор не образуют. Оптимум pH для роста 5,0—7,5, при pH 4,0 и ниже рост прекращается.
Подобно бактерии Obesumbacterium proteus бактерии группы кишечной палочки очень быстро растут в сусле и образуют большое число различных метаболитов, влияющих на вкус и аромат пива, которое становится сладковатым с фруктовым привкусом или приобретает запах вареной капусты [136]. В результате жизнедеятельности может образоваться и сероводород. Эти бактерии обладают разнообразной биохимической активностью. Например, превращение глюкозы может идти гликолитическим и гексозомонофосфатным путями, а также с использованием механизма Энтнера—Дудорова (рис. 3).
Пировиноградная кислота, образующаяся в этих реакциях, в кислых условиях может давать молочную и уксусную кислоты, этанол, СО2 и водород.
Бактерии рода Klebsiella обладают способностью конденсировать две молекулы пировиноградной кислоты, образуя ацетоин и СО2. Ацетоин, являясь акцептором водорода, дает 2,3-бутандиол.
Клетки бактерий Klebsiella короткие, единичные, неподвижные, размер их (0,5—0,8) X (1,0—2,0) мкм. Бактерии могут размножаться как в сладком, так и в охмеленном сусле даже при температуре 40° С, увеличивая за 7 ч число клеток в миллионы раз, могут сохраняться и в готовом пиве.
Наличие бактерий группы кишечной палочки является показателем санитарного состояния производства и соблюдения персоналом правил личной гигиены [31], поскольку эти бактерии обычно живут в кишечнике здоровых людей и животных, являясь типичными представителями фекальных бактерий.
Бактерии группы кишечной палочки могут попасть в пиво в результате использования недостаточно чистой воды, из воздуха, от загрязненных рук и одежды рабочих, с возвратной тарой и т. д. Наличие бактерий этой группы характеризуется двумя взаимозаменяемыми показателями:
- коли-титром — наименьшим объемом жидкости, в котором обнаруживается хотя бы одна бактерия. Для воды коли-титр по ГОСТу должен быть не ниже 300. Для пива коли-титр не регламентируется, но он не должен значительно отличаться от коли-титра водопроводной воды;
- коли-индексом—количеством бактерий в 1 л жидкости.
Дикие дрожжи. В пивоваренных материалах часто обнаруживаются клетки различных видов диких дрожжей. Вайлес [249] выделил 27 штаммов дрожжей, в том числе из хмеля 12, из задаточных дрожжей 11. Оказалось, что порчу пива вызывают 11 штаммов, выделенных из различных источников.
Наиболее часто встречаются дикие виды Saccharo- myces cerevisiae, Pichia membranaefaciens, Torulopsis sp. и Candida sp., реже — Hanseniaspora, Kloeckera, Rhodotorula.
Бреди [94] исследовал дикие дрожжи, инфицирующие культурные дрожжи, на ряде пивоваренных заводов. Всюду были обнаружены Pichia membranaefaciens, Candida mycoderma, Torulopsis colliculosa и T. incons- picula [18], реже встречались дрожжи родов Candida, Debaryomyces, Hansenula и Rhodotorula. Разводку задаточных дрожжей высевали на специальную среду, содержащую лизин в качестве единственного источника азота. На этой среде виды Saccharomyces не росли, остальные виды образовывали колонии. На 10 млн. клеток культурных дрожжей приходилось 1—30 клеток диких дрожжей, в одном случае — 410 клеток.
Фин и Гусева [75] из 35 образцов пива выделили в чистую культуру 38 штаммов дрожжей, половина из которых была представлена культурными дрожжами. Распределение выделенных штаммов диких дрожжей представлено в табл. 8.
Таблица 8 Таксономия диких дрожжей, выделенных из пива
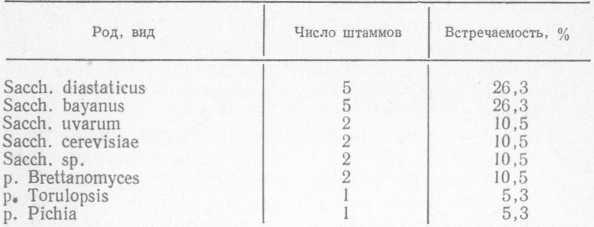
Из данных табл. 8 видно, что большинство выделенных дрожжей принадлежит к роду Saccharomyces. Порча пива, обусловленная дикими дрожжами, может проявляться в помутнении, образовании пленки на поверхности и в изменении вкуса и аромата. Sacch. diastaticus, способные сбраживать декстрины, обладают глюкоамилазной активностью и расщепляют а-1,4-связи в нераз- ветвленных молекулах мальтодекстринов. Хроматографический анализ показал, что при инфицировании S. diastaticus в пиве увеличивается содержание высших спиртов и эфиров (в мг/л): этилацетата 38, пропилового спирта 40, изоамилацетата 7,6 (контрольное пиво содержало соответственно 9, 11, 5 и 1,5 мг/л) [158].
Было найдено, что S. cerevisiae var. turbidans могут вызвать помутнение бочкового пива даже в том случае, если на 1,6-108 клеток культурных задаточных дрожжей приходится всего одна клетка их.
Дрожжи Pichia и Candida могут окислять этанол.
Помутнение пива, вызываемое дикими дрожжами, обусловлено тем, что эти дрожжи медленнее оседают и плохо отделяются при фильтрации; некоторые дикие дрожжи не погибают даже при пастеризации.
Полагают, что имеются существенные различия между культурными и дикими дрожжами в образовании эфиров, высших спиртов, дикетонов и других продуктов, имеющих большое значение для формирования вкуса и запаха пива [149]. Однако в этом направлении требуются дальнейшие исследования.
