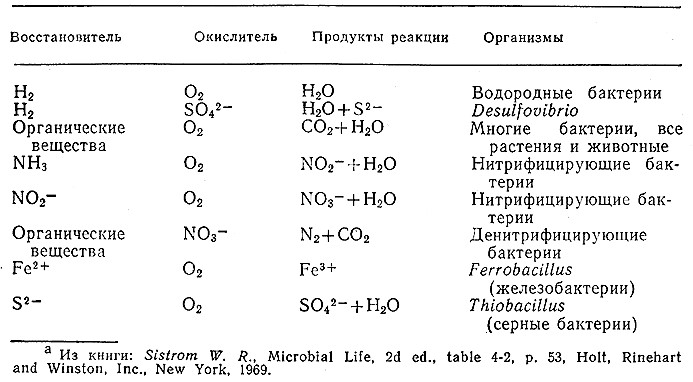
Дыханием называется сопровождающийся выделением энергии процесс, в котором органические или восстановленные не» органические соединения окисляются в результате взаимодей· ствия с неорганическими веществами. Как показано в табл. 5.4, в различных бактериях процесс дыхания осуществляется с участием различных восстановителей и окислителей. Если окислителем является не кислород, то процесс называют анаэробным дыханием, а термин аэробное дыхание применяют в тех типичных для эукариот и многих бактерий случаях, когда роль окислителя выполняет O2. Напомним, что литотрофы используют неорганические восстановители. Примеры литотрофов также приведены в табл. 5.4. По имеющимся в настоящее время данным все литотрофы являются аутотрофами, так как источником углерода для них служит CO2.
Таблица 5.4. Восстановители и окислители в процессах дыхания бактерийa
B большинстве форм дыхания органические вещества окисляются под действием кислорода. Далее в настоящем разделе мы будем рассматривать только такую форму дыхания и, употребляя термин «дыхание», будем подразумевать только аэробный процесс. Удобно подразделять весь процесс дыхания на две фазы, в первой из которых органические вещества окисляются до CO2, а атомы водорода (электроны) захватываются NAD. B следующей фазе атомы водорода участвуют в ряде последовательных химических превращений, в результате которых из ADP регенерируется ATP, а сами атомы водорода соединя- ются с кислородом, образуя воду. Ниже мы рассмотрим эти две фазы более детально.
5.4.1. Цикл трикарбоновых кислот
Сначала в дополнение к разд. 5.3, посвященному метаболизму углеводов, отметим, что все описанные там пути метаболизма, ведущие к пирувату, могут реализоваться и в процессе дыхания, а свойственные собственно дыханию реакции начинаются со стадии образования пирувата. В то же время в метаболизме дыхания пируват не восстанавливается до какого- либо конечного продукта с помощью образовавшихся в процес- серасщепления глюкозы атомов водорода; последние используются для других целей, которые мы вкратце здесь также рассмотрим. Более того, в процессе превращения пирувата в производное уксусной кислоты (ацетил-СоА) генерируется дополнительное количество универсального восстановителя NADH:
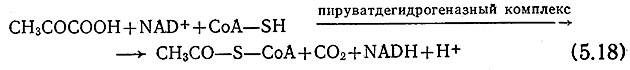
Ацетил-СоА (см. рис. 2.9) является также ключевым промежуточным веществом в катаболизме аминокислот и жирных кислот; следовательно, три класса органических соединений могут окисляться до ацетил-СоА.
Первая фаза этого окисления осуществляется в замкнутой последовательности реакций, называемой циклом трикарбоновых кислот (TKK), циклом Кребса или циклом лимонной кислоты (рис. 5.7). Обратите внимание на то, что два оставшихся атома углерода пирувата (один теряется ранее в виде СО2 при образовании ацетил-СоА) включаются в цикл путем образования карбоновой кислоты с 6 углеродными атомами из ацетил-СоА и С4-кислоты. B дальнейших стадиях цикла TKK, однако, два атома углерода выделяются в виде СО2, и таким образом в этой первой фазе дыхания потребляются все углеродные атомы исходного пирувата.
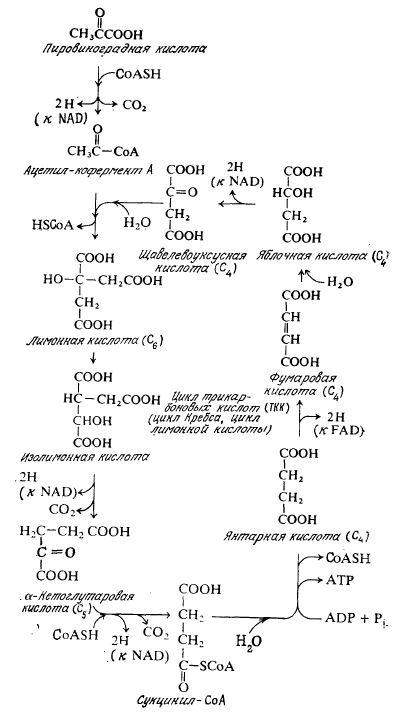
Рис. 5.7. Цикл трикарбоновых кислот
Общая стехиометрия цикла TKK выглядит следующим образом:
C3H4O3+ADP+Pi+2Н2O+FAD+4NAD+ → 3CO2+ATP+FADH2+4 (NADH+H+) (5.19)
Может показаться, что цикл TKK (в таком виде, как он изображен на рис. 5.7) выполняет только каталитическую функцию, поскольку здесь не указано никакого иного источника углерода, кроме субстрата. Ha самом же деле, как показано на рис. 5.1, цикл TKK очень важен и в качестве источнйка предшественников для последующих биосинтетических реакций. Следовательно, некоторые промежуточные соединенияНИС. 5.7. Цикл трикарбоноых кислот.зтого цикла постоянно отбираются для целей биосинтеза и их количества должны возобновляться. Эта цель достигается пу· тем синтеза щавелевоуксусной кислоты из пирувата или другой кислоты с тремя углеродными атомами. B некоторых микро· организмах недостаток промежуточных веществ в цикле TKK возмещается за счет глиоксилатного цикла, в котором в результате конденсации двух молекул ацетата образуется молекула сукцината.
5.4.2. Дыхательная цепь
Оставив на время метаболизм соединений углерода, давайте проследим за реакциями, в которых атомы водорода окисляются до воды, — именно этот процесс в основном обеспечивает аэробные клетки энергией. B каждом единичном цикле TKK высвобождаются четыре пары атомов водорода, три из которых переносятся к NAD, а одна, образующаяся при дегидрировании янтарной кислоты, как показано на рис. 5.7, переносится к фла- винадениндинуклеотиду (FAD) (см. рис. 2.9). Высвобождающиеся в цикле ТКК восстанавливающие эквиваленты частично используются в биосинтетических реакциях, а частично — в регенерации ATP.
B последующем обсуждении регенерации ATP в процессе дыхания мы будем уделять особое внимание такой ситуации, когда в реакциях дыхательной цепи участвуют все атомы водорода, образующиеся при расщеплении глюкозы. Именно таким путем создается большая часть энергетических запасов клетки в виде ATP. Ha рис. 5.8 в сокращенном виде приведена последовательность реакций в дыхательной цепи, а также их связь с процессом полного расщепления пирувата через ацетил- CoA в цикле TKK (Кребса). Ha этой схеме символами FP1 и FP2 обозначены два различных флавопротеина, которые представляют собой ферменты, содержащие FAD и участвующие в транспорте электронов. Электроны от NADH (NADH = FP1) передаются кофермеяту Q (обозначенному на рисунке символом Q); в этом процессе передача каждой пары электронов сопровождается фосфорялированием однОй молекулы ADP до ATP. Электроны, высвобождающиеся в цикле TKK в ходе дегидрирования сукцината, передаются при посредстве FAD (в FP2) непосредственно коферменту Q.
Начиная с этого момента судьба всех электронов одинакова: все они взаимодействуют с рядом цитохромов — гем-содер- жащих белков, обозначенных на рис. 5.8 символами b, с, α и α3- B ходе этого взаимодействия каждая пара электронов генерирует две молекулы АТР. Процесс регенерации ATP в дыхательной цепи называется окислительным фосфорилированием.
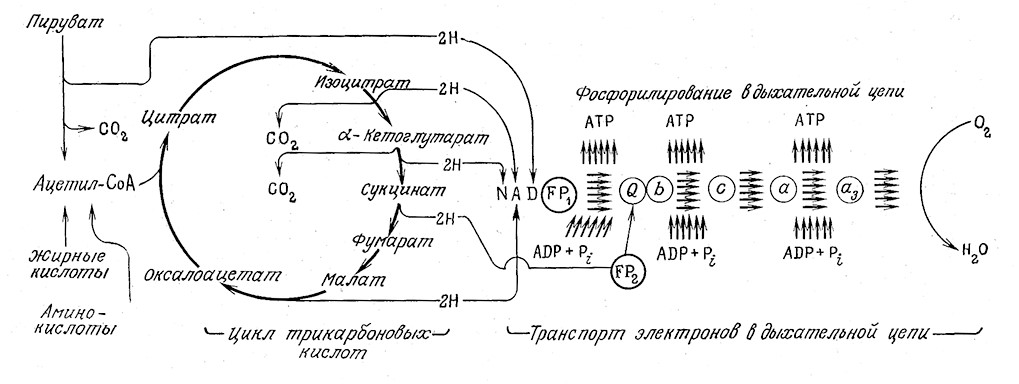
РИС.5.8.ФосфорилированиеАОР в дыхательной цепи с участием высокоэнергетических электронов, высвобождающихся при окислении углеводов, жирных кислот и аминокислот (FP1 — NADH; FP2 — сукцинатдегидрогеназа; Q — кофермент Q; b, с, α и α3—соответствующие цитохромы).
B конце концов атомы водорода вступают в реакцию с растворенным кислородом, в результате чего в качестве второго конечного продукта окисления образуется вода. Если проследить за изменениями свободной энергии на отдельных стадиях дыхательной цепи, нетрудно заметить, что ATP регенерируется во всех случаях, когда уменьшение свободной энергии превышает величину 7,3 ккал/моль, необходимую для фосфорилирования ADP.
Таким образом, все происходящие в дыхательной цепи реакции в конечном счете описываются следующими уравнениями:
NADH+H+ + 1/2O2+3ADP+3Pi → NAD++4H2O+3ATP (5.20)
и
FADH2+1/2O2+2ADP+2Pi →.FAD+3H2O+2ATP (5.21)
Потенциальные возможности дыхания в отношении обеспечения клетки энергией намного превосходят возможности гликолиза, поскольку изменение стандартной свободной энергии ΔG°' в реакции
Глюкоза+6О2 → 6СO2+6Н2O (5.22)
составляет —686 ккал/моль. Можно проверить, насколько эффективно используется этот богатый источник энергии в живых аэробных организмах. Для сравнения с реакцией (5.22) допустим, что глюкоза полностью окисляется до CO2 и H2O по метаболическим путям ЭМП, TKK и в дыхательной цепи. При этом не следует забывать, однако, что путь ЭМП и цикл TKK являются амфиболическими и что, следовательно, часть промежуточных соединений постоянно отвлекается, включаясь в биосинтетические пути метаболизма. Поэтому на самом деле глюкоза не всегда претерпевает именно эти превращения, и приведенные ниже расчеты позволяют оценить только высший предел выхода ATP из глюкозы в аэробной клетке.
Путем сложения уравнения (5.12) (путь ЭМП) с умноженным на два уравнением (5.19) (цикл TK.K), умноженным на десять уравнением (5.20) [окисление всех NADH + H+, образующихся по пути ЭМП (дважды), и реакций циклаТКК (четыре раза)] и умноженным на два уравнением (5.21) [окисление FADH2] получим
C6H12O6+38ADP+38Pi+6O2 → 6СO2+38АТР+44Н2O (5.23)
Поскольку изменение стандартной свободной энергии при гидролизе ATP равно —7,3 ккал/моль, то изменение свободной энергии в реакции (5.23) составит приблизительно
ΔG°'≈(38 моль ATP/моль глюкозы) (7,3 ккал/моль ATP) = —277 ккал/моль глюкозы (5.24)
Эта величина в 19 раз превышает энергию, усваиваемую клеткой в ходе гликолиза. Как и в гликолизе, эффективность процесса дыхания очень высока:
![]()
Если ввести поправки на нестандартные концентрации в клетке, то мы получим поразительно высокую степень эффективности, превышающую 70%. Большая часть не использованной в процессе регенерации ATP энергии рассеивается в виде теплоты, которая должна тем или иным способом отводиться, чтобы температура не вышла за рамки допустимых для клетки физиологических пределов.
Поскольку в практической термодинамике химические машины обычно не изучаются, здесь может оказаться полезной аналогия с классическим примером сжатия газа в цилиндре с поршнем. Если газ сжимают быстро, то большое количество энергии теряется в виде теплоты и поэтому не может быть использовано при последующем расширении. Аналогичным процессом является совершенно неэффективное сжигание глюкозы на воздухе. K идеальному обратимому процессу приближается сжатие газа путем очень медленного движения поршня, когда выделяется минимальное количество тепла. Точно так же и процессы в живой клетке приближаются к обратимым и максимально эффективным с точки зрения извлечения энергии благодаря тому, что окисление глюкозы осуществляется в несколько стадий, каждая из которых характеризуется относительно небольшим изменением свободной энергии.
Описанные выше реакции в дыхательной цепи типичны для обычной эукариотической клетки. B случае прокариот транспорт электрона к атому кислорода может сопровождаться фосфорилированием менее трех молекул ADP. Кроме того, цитохромный путь может несколько видоизменяться в зависимости от вида прокариоты, а на эффективность фосфорилирования могут влиять условия роста клеток. Для оценки эффективности усвоения энергии в дыхательной цепи используется особый параметр, отношение P/O, равное числу актов фосфорилирования в расчете на один усвоенный атом кислорода.
Позднее в этой главе мы узнаем, каким образом степень окисления субстрата и биосинтетические потребности клетки совместно налагают определенные ограничения на стехиометрию амфиболических путей. Прежде, однако, мы завершим наш обзор путей метаболизма как источников энергии, вкратце рассмотрев фотосинтез — процесс преобразования энергии, без которого была бы невозможна жизнь на земле.
