B большинстве типов дыхания атомы водорода постоянно переносятся от топлива к кислороду; этезг процесссопровожда- ется выделениемэнергии. Фотосинтез представляет собой, по сути дела, обратный процесс, в котором осуществляется улавливание световой энергии, используемой для превращения диоксида углерода в глюкозу и ее полимеры. Фотосинтез является первичным поставщиком энергии для всей биосферы. B фотосинтезе усваивается энергия единственного значимого источника энергии в Солнечной системе — самого солнца. Другая жизненно важная функция фотосинтеза заключается в его участии в круговороте углерода и кислорода; именно благодаря фотосинтезу осуществляется восстановление углерода, окисленного в ходе дыхания:
Фотосинтез: 6СO2+6Н2O+свет → C6H12O6+6O2 (5.26)
5.5.1. Аккумулирование световой энергии
В. прокариотах (цианобактериях, называемых также сине- зелеными водорослями; зеленых серобактериях; пурпурных серобактериях) фотосинтезосуществляетсяв многослойных мембранах, а в эукариотах (водорослях, растениях) центром фотосинтеза являются особые органоиды, называемые хлоропласта- ми (рис. 1.5). И те и другие системы содержат хлорофилл — сложное органическое соединение, сильно поглощающее свет в видимой области спектра (рис. 5.9). Энергия одного фотона Ep зависит от его частоты (или длины волны) следующим образом:
![]()
где h — константа Планка; v — частота фотона; λ — длина волны фотона; с — скорость света в среде. Два основных пика в спектре поглощения хлорофилла (рис. 5.9) соответствуют энергиям фотонов 43,5 ккал/моль (650 нм) и приблизительно 67 ккал/моль (430 нм). Обе полосы поглощения, очевидно, соответствуют свободной энергии, значительно превосходящей необходимый для фосфорилирования ADP минимум.
Известны две различные системы аккумулирования световой энергии и сопровождающих этот процесс химических реакций, называемые фотосистемами I и II. Фотосистемы I и II активируются светом с длиной волны более и менее 680 нм соответственно. Обе системы обнаружены во всех фотосинтезирующих организмах, выделяющих кислород. Напротив, фотосинтезирующие бактерии, не выделяющие кислород, содержат только фотосистему I.
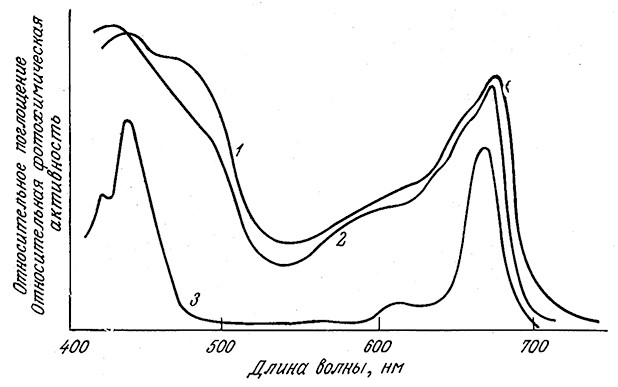
РИС. 5.9. Максимумы в спектре поглощения хлорофилла а (нижняя кривая)
связаны с максимумами поглощения в спектре зеленых листьев (верхняя кривая)
и со спектром фотосинтетической активности листьев (средняя кривая).
Соответствующие пигменты листьев способствуют повышению эффективности
процесса утилизации света в диапазоне 500 – 600 нм по сравнению с чистым хлорофиллом.
5.5.2. Транспорт электронов и фотофосфорилирование
B фотосистемах I и II поглощение двух световых квантов (фотонов) хлорофиллом и другими пигментами приводит к возбуждению электрона в реакционном центре (например, P700 в случае фотосистемы I; см. рис. 5.10). Обладающий достаточно высокой энергией электрон включается в цепь электронного транспорта. Затем электрон «падает» на низший энергетический уровень (этот процесс изображен на рис. 5.10 как повышение значения стандартного электродного потенциала Eoi). B фотосистеме I электрон в конечном счете используется для восстановления NADP+, а возбужденные электроны фотосистемы II передаются фотосистеме I, причем в процессе их транспорта каждая пара возбужденных электронов участвует в фосфорилировании одной молекулы ADP. Таким образом, общая стехиометрия реакций, индуцируемых фотонами в фотосинтезирующих эукариотах, выглядит следующим образом:
H2O+4hv+NADP++ADP+Pi → NADPH+H++1/2O2+(ATP+H2O) (5.28)
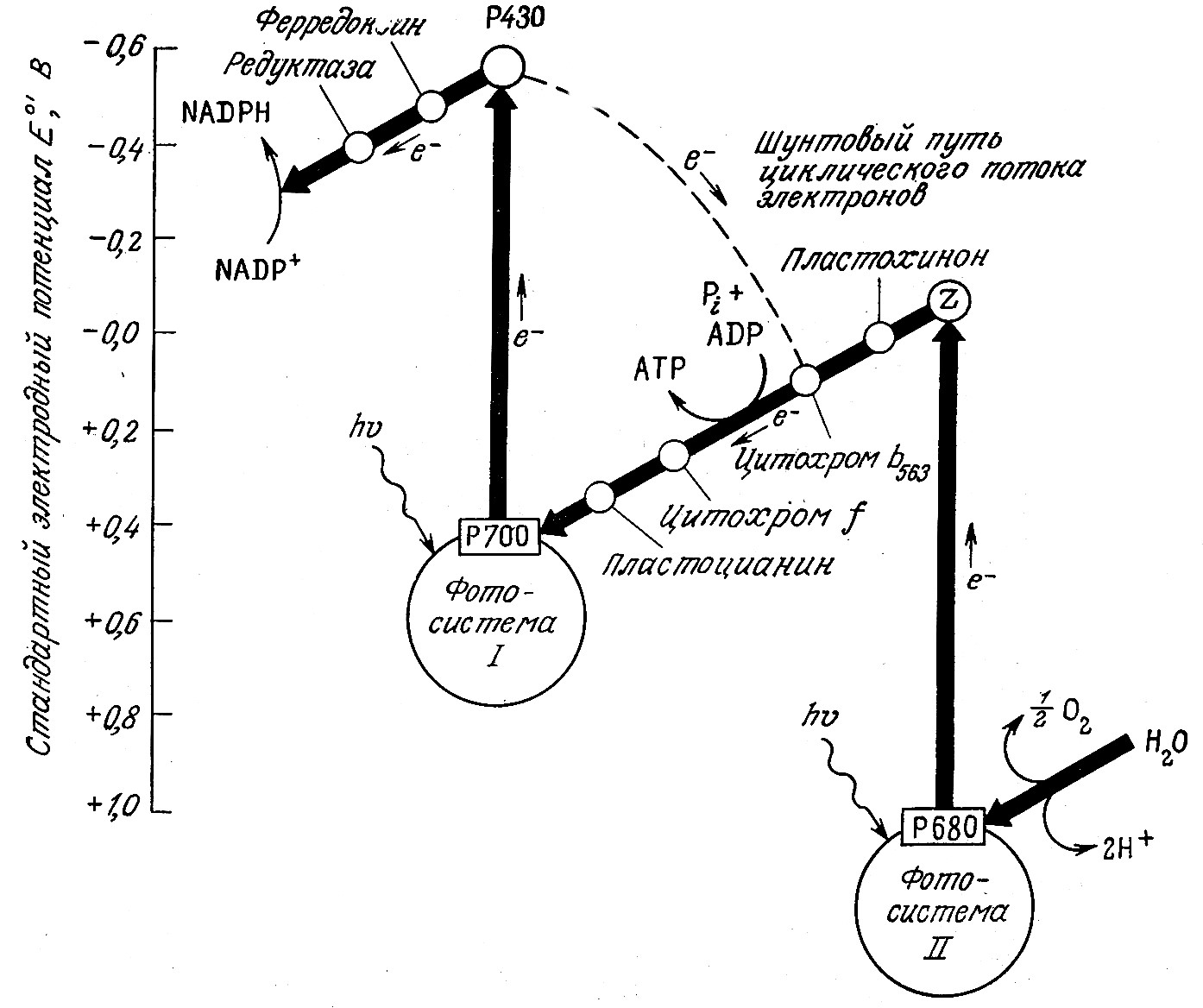
РИС. 5.10. Инициированное поглощением света возбуждение электронов (вертикальные стрелки) и их транспорт в фотосинтезирующих системах
B используемом диапазоне энергий светового возбуждения только при совместной работе двух фотосистем вода может служить донором (Eoi = + 0,82 В), а NADP+ — акцептором (EOi = —0,32 В) электронов. Уравнение (5.10) объясняет, почему, например, в бактериях, использующих в качестве донора электронов H2S, фотосинтез может осуществляться только фотосистемой I: это связано с величиной стандартного электродного потенциала восстановления серы Е°', равного —0,23 В.
Поглощенная световая энергия может участвовать в регенерации дополнительных количеств ATP по пути циклического фотофосфорилирования, изображенному на рис. 5.10 штриховой линией. B этом случае возбужденные электроны транспортируются от P700 к P430, затем к цитохрому Ь563 и снова к P700; на последнем этапе этого путп электроны принимают участие в фосфорилировании ADP по следующему простому уравнению:
3hv+ADP + Pi → ATP + H2O (5.29)
Использование накопленных рассмотренными выше путями энергии и восстанавливающих эквивалентов длясинтеза глюкозы из CO2 и H2O мы опишем в следующем разделе. Здесь же отметим, чтоизменение свободной энергии вреакции фотосинтеза
6CO2+6H2O → С6Н12O6+6O2 (5.30)
равно + 686 ккал/моль. Поскольку в любой из фотосистем для возбуждения одного электрона необходимы два фотона и поскольку генерирование одной молекулы O2 сопровождается переносом четырех электронов, то обрааование 6O2 требует поглощения 48 квантов света. Согласно уравнению (5.27), энергия фотонов зависит от длиныихволны.Еслипринять λ = 700нм, то минимальное количество световой энергии, необходимой для инициирования реакции (5.30) с помощью фотосинтезирующей системы растений, составит 48-41 = 1968 ккал/моль глюкозы. Следовательно, в первом приближении эффективность утилизации поглощенного света будет равна 686: 1968 = 0,35 = 35%. Действительную эффективность фотосинтеза в растениях и микроорганизмах мы рассмотрим позднее, в ходе обсуждения общей стехиометрии метаболизма.
