Нормальная жизнедеятельность клетки возможна только при условии регулируемого транспорта ионов и молекул между клеткой и средой. Клеточные мембраны обладают селективной проницаемостью по отношению к различным компонентам и составным частям клетки и тем самым регулируют транспортные процессы, выполняющие ряд важных функций. Во-первых, благодаря процессам транспорта рН и состав внутриклеточной среды поддерживаются на одном уровне в достаточно узком диапазоне, обеспечивающем необходимую активность ферментов. Кроме того, мембранный транспорт регулирует объем клетки, обеспечивает доступ и нужную концентрацию питательных веществ, удаление токсичных соединений.
Известны по меньшей мере три способа транспорта через клеточные мембраны: пассивная диффузия, сопряженныйтранс- порт и активный транспорт. Независимо от механизма способность данной мембраны транспортировать данный субстрат часто выражают как проницаемость мембраны К, которую рассчитывают по уравнению
![]()
где V — объем клетки; A — наружная поверхность клетки; Cе — наружная концентрация субстрата; Cl0 — начальная внутриклеточная концентрация субстрата; Ci(t)—внутриклеточная концентрация субстрата через время t. Из уравнения (5.40) следует, что проницаемость имеет ту же размерность, что и скорость (сантиметров в секунду). Как мы увидим далее в ходе решения упражнений, лежащие в основе этого уравнения допущения ограничивают его применимость.
5.7.1. Пассивная диффузия и сопряженный транспорт
Пассивная диффузия представляет собой транспорт вещества через мембрану из области с высокой концентрацией в область с низкой концентрацией (рис. 5.17). Скорость диффузии пропорциональна общей движущей силе, которой в данном случае является разность концентраций по обе стороны мембраны. Термодинамический анализ показывает, что пассивная диффузия спонтанна, поскольку транспорт вещества из области с концентрацией с2 в область с концентрацией C1 сопровождается изменением свободной энергии ΔGo, равным
![]()
Так как c1 меньше с2, то ΔGo в случае пассивной диффузии всегда отрицательно. Если транспортируемое вещество заряжено, то уравнение (5.41) следует видоизменить следующим образом:
![]()
где Z1 — заряд транспортируемых молекул; F — число Фарадея [23,062 ккал/(В-моль)]; Δψ — разность потенциалов на мембране, В.
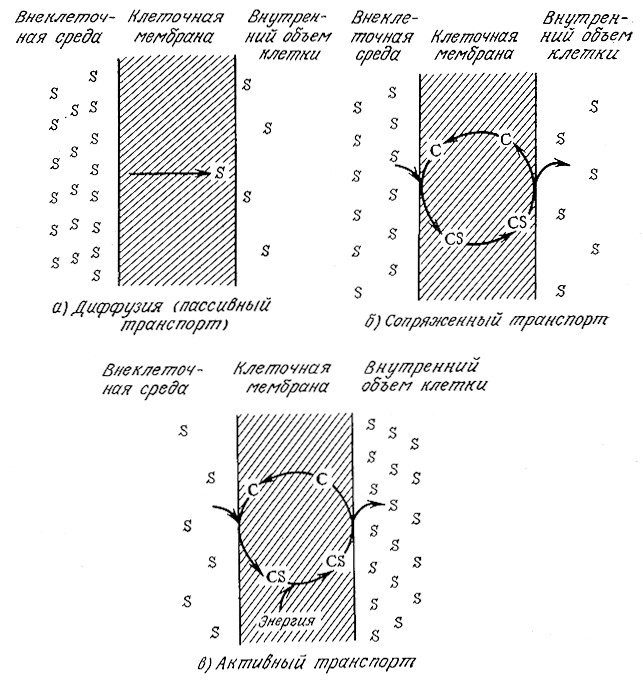
РИС. 5.17. Различные механизмы транспорта через мембрану:
а — диффузия; б — сопряженный транспорт;
в — активный транспорт (S—транспортируемое вещество;
C — белок-переносчик; CS — комплекс).
B силу специфичности структуры плазматических мембран, окружающих любые клетки и органоиды эукариот, не все соединения проникают через мембрану одинаково легко. Существует корреляция между скоростью диффузии большинства сравнительно объемных молекул и их растворимостью в липофильных средах (рис. 5.18). Это и неудивительно, поскольку основой плазматических мембран, как полагают, является главным образом липидный бислой (вспомните рис. 2.2). Аномально высокую скорость диффузии воды и других очень малых молекул объясняет другая модель, предусматривающая наличие в мембранах пор небольшого диаметра.
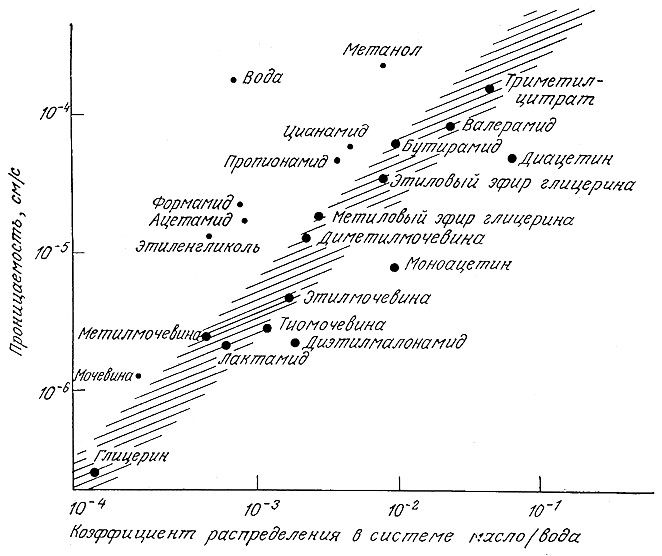
РИС. 5.18. Способность многих веществ проникать через мембрану водорослей Chara
коррелирует с их растворимостью (коэффициентом распределения) в липофильных растворителях.
Размеры точек примерно пропорциональны размерам соответствующих молекул.
Для ионизированных и полярных веществ характерна очень низкая растворимость в липофильных растворителях, поэтому такие вещества практически не транспортируются через мембрану за счет пассивной диффузии. Важность этого обстоятельства мы уже не раз подчеркивали: промежуточные метаболиты большей частью представляют собой ионизированные соединения и поэтому удерживаются внутри клетки. Впрочем, некоторые полярные вещества очень легко преодолевают мембранный барьер.
Одним из механизмов, объясняющих такое аномальное поведение полярных веществ, является механизм сопряженного транспорта. Как схематично показано на рис. 5.17, согласно это му механизму субстрат на наружной поверхности мембраны соединяется с молекулой-переносчиком·, образующийся комплекс далее «диффундирует» к другой поверхности мембраны и там расщепляется, выделяя перенесенную молекулу во внутренний объем клетки. Такой способ биологического транспорта имеет ряд характерных особенностей. Так, скорость транспорта вещества в этом случае изменяется в зависимости от разности концентраций не линейно, а достигает некоторого максимума (уровня насыщения), после которого дальнейшее повышение разности концентраций вещества не изменяет скорость его транспорта.
Зная уравнение Михаэлиса— Ментен и другие математические модели кинетики ферментативного катализа, нетрудно понять, почему изображенный на рис. 5.17, б процесс назван сопряженным транспортом.
Другие важные свойства сопряженного транспорта также напоминают особенности ферментативного катализа; так, через мембрану транспортируются только определенные вещества, а специфические ингибиторы замедляют процесс. Специфичность и кинетические характеристики свидетельствуют о том, что носители представляют собой белки. Многие из носителей удалось выделить и охарактеризовать; их называют пермеазами. Вероятно, самым известным примером сопряженной диффузии является транспорт глюкозы в эритроцитах (красных кровяных тельцах) человека. Подробные сведения об этой системе и дополнительную информацию о сопряженном транспорте читатель может найти в литературе.
5.7.2. Активный транспорт
Как мы уже показывади на рис. 5.17, для активного транспорта характерно, во-первых, движение вещества против его химического (или электрохимического) градиента, т. е. из области с низкой концентрацией в область с высокой концентрацией, и, во-вторых, как показывает уравнение (5.41), необходимость затрат метаболической энергии. Например, если соединение, не несущее заряда, транспортируется из среды, где его концентрация равна 0,001M, в клетку, где его концентрация составляет 0,1 M, то
![]()
Следовательно, для реализации этого процесса необходимо затратить по меньшей мере указанное количество энергии. Активный транспорт также осуществляется при участии специфических псрмеаз.
Активный транспорт лежит в основе передачи нервных импульсов, но клетки используют этот механизм в гораздо более широких масштабах. Так, почти все клетки обладают системами активного транспорта (ионными насосами), позволяющими поддерживать необходимые концентрации ионов K+, Na+ и воды внутри клетки. Эти системы перемещают ионы Na+ из клетки, а ионы K+ в клетку, что позволяет компенсировать пассивную диффузию этих ионов. Указанные системы транспорта взаимосвязаны и сопряжены с гидролизом АТР (рис. 5.19). B красных кровяных тельцах роль натрий-калиевого насоса выполняет оли- гомерный белок, называемый Na+—К+-АТРазой. Как показано на рис. 5.19, этот белок располагается втолще плазматической мембраны и пронизывает ее насквозь; такие белки называют трансмембранными.
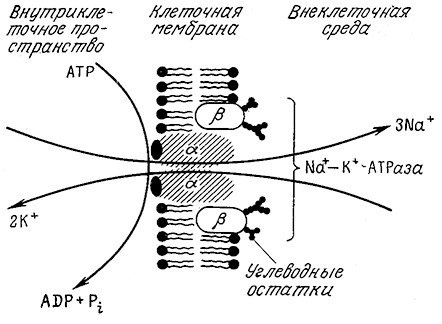
РИС. 5.19. Na+—K+-Hacoc (Na+—K+-ATPaзa) транспортирует Na+ из клетки
и K+ в клетку; движущей силой этого процесса является гидролиз ATP
Вторая обычная группа систем активного транспорта обеспечивает поступление в клетку молекул питательных веществ, например, глюкозы и аминокислот, со скоростями, значительно превышающими скорость их транспорта за счет пассивной диффузии. B клетках высших животных активный транспорт глюкозы совмещен с транспортом Na+ в клетку, а удаление Na+ из клетки осуществляется параллельно с гидролизом АТР под действием Na+—К+-АТРазы. Весь процесс схематично изображен на рис.5.20. B бактериях для активного транспорта глюкозы используется другой процесс, называемый транслокацией групп. B этом процессе глюкоза высвобождается во внутриклеточное пространство в виде высокоэнергетического глюкозо-6-фосфата, с трудом проникающего через мембраны (рис. 5.21). Считается, что этот процесс является той стадией, скорость которой определяет скорость роста ряда клеток.
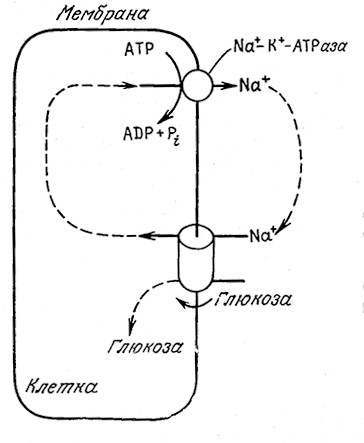
РИС. 5.20. Ионы Na+ и глюкоза транспортируются из среды
в клетку одним и тем же пассивным переносчиком.
Низкая внутриклеточная концентрация Na+ поддерживается
за счет активного транспорта.
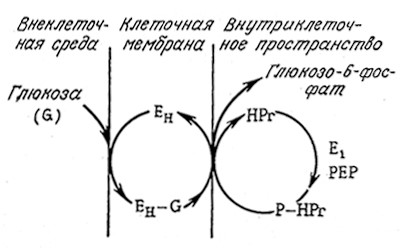
РИС. 5.21. Фосфорилирование глюкозы в клеточной мембране
создает постоянную движущую силу для транспорта глюкозы через мембрану
и удерживает глюкозу (в виде производного) внутри клетки.
Здесь G — глюкоза; Ен— мембранный фермент; E1 — цитоплазматический фермент;
HPr — цитоплазматический белок; PEP — фосфоенолпируват.
Движущей силой многих систем транспорта в бактериях служит поток протонов через плазматическую мембрану. Основная роль потока протонов в энергетике бактерий описывается хеми- осмотической теорией, разработанной Митчеллом в 1961 г. Ha рис. 5.22 изображена схема совместного транспорта протонов и лактозы в процессе дыхания бактерии E. coli. Протоны и лактоза совместно транспортируются в клетку лактозопермеазой (продуктом транскрипции гена у оперона lac). Одновременно в дыхательной цепи протоны вместе с потоком электронов выводятся из клетки. B целом pH внутри клетки выше, чем в непосредственном ее окружении.
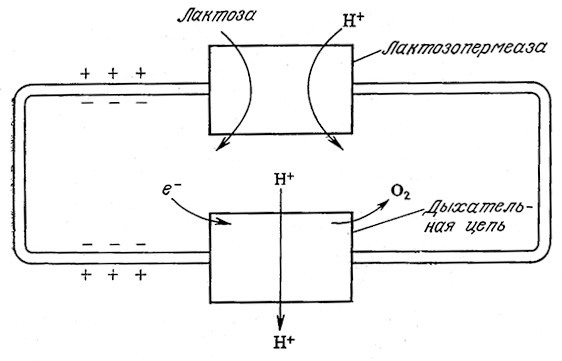
РИС. 5.22. Сочетание активного транспорта лактозы с транспортом протонов
по градиенту электронного потока дыхательной цепи.
Эукариотические клетки должны обладать целым комплексом систем транспорта, поскольку концентрации многих веществ в органоидах поддерживаются на уровне, отличающемся от концентрации этих веществ в цитоплазме. Этот факт лишний раз говорит о том, что любая клетка, и особенно эукариотическая, представляет собой не однородную смесь множества компонентов, а высокоорганизованную, вплоть до молекулярного уровня, систему.
Прежде чем перейти к изучению потоков и регуляторных систем в последовательностях метаболических реакций, полезно несколько полнее и детальнее рассмотреть структуруи организацию клеточной мембраны. Ha рис. 5.23 схематично изображено строение клеточной оболочки бактерии; нетрудно видеть, что ее структура значительно сложнее описанной в гл. 2 упрощенной модели, включающей липидный бислой и белковую оболочку. Ha рисунке показано, что в толще мембраны находится множество белковых молекул одни из них располагаются у наружной или внутренней поверхности мембраны, другие молекулы пронизывают всю толщу мембраны. Эти белки (пермеазы и белки-рецепторы) узнают специфические соединения в среде и контактируют с регуляторными системами клетки, обеспечивая ее ответную реакцию на изменения в среде. B случае бактерий белковые компоненты мембран позволяют клеткам быстро приспособиться к изменившимся условиям окружения. Поверхностные рецепторы высших эукариотичеоких клеток участвуют в процессах клеточного узнавания, играющих большую роль как в организации тканей, так и в иммуннойреакции клеток.
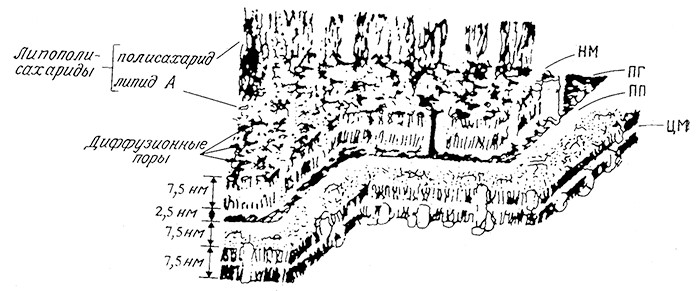
РИС. 5.23. Детали строения клеточной оболочки E. coli. Здесь белые глобулярные объекты
на поверхности и в толще клеточных мембран обозначают отдельные белки
(HM — наружная мембрана; ПГ — пептидогликан; ПП — периплазматическое пространство;
ЦМ — цитоплазматическая мембрана)
Мембраны в общем случае асимметричны; их наружные и внутренние повёрхности содержат различные компоненты и выполняют различные функции. Липиды и разнообразные мембранные белки диффундируют в плоскости мембраны очень быстро; по некоторым оценкам скорость диффузии липидов в рядё мембран составляет 10–8 см2/с.
