Большое влияние на смертельное время оказывает количество микроорганизмов. В качестве примера в табл. 18.2 приведены данные одного исследования, которые подтверждают: чем больше микроорганизмов находится в определенном объеме стерилизуемого продукта перед стерилизацией, тем больше времени требуется на их уничтожение.
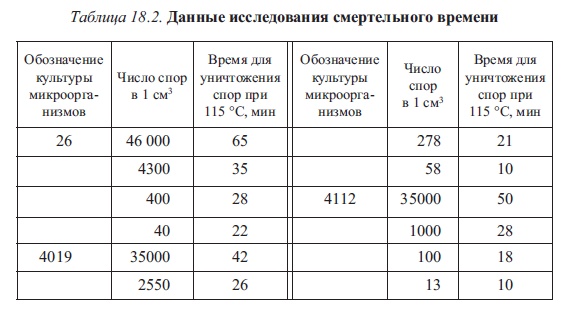
Как следует из табл. 18.2, для культуры 26 при уменьшении начального содержания микроорганизмов с 46 000 до 40 спор, т.е. примерно в 1000 раз, смертельное время сокращается приблизительно в 3 раза.
Возможные объяснения:
- при большом количестве микроорганизмов в продукте они распределены неравномерно, образуют скопления (агрегаты). К микроорганизмам в центре такого скопления тепло проникает хуже;
- при большом количестве микроорганизмов велика вероятность появления термоустойчивых особей (даже в одной колонии термоустойчивость клеток различается).
С физико-химической стороны процессы, вызывающие смерть микробной клетки, представляют собой мономолекулярную реакцию коагуляции белков протоплазмы. Скорость уничтожения микробов поддается математическому анализу, справедливому для реакции первого порядка. Скорость уничтожения микробов dN/dx, представляющую собой производную количества микроорганизмов по времени, можно записать так:
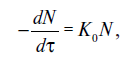
где N - количество микроорганизмов в данный момент стерилизации; К0 - коэффициент скорости уничтожения микробов (величина, обратная времени).
Знак «минус» указывает на то, что при нагревании с течением времени количество микроорганизмов уменьшается.
После математических преобразований получаем
![]()
где N0 - количество микроорганизмов к началу процесса стерилизации; N - количество микроорганизмов в конце процесса стерилизации при определенной температуре; К - константа, учитывающая скорость уничтожения микробов и переход к десятичному логарифму, К = К0/2,303.
Из уравнения (18.2) видно, что скорость уничтожения микроорганизмов подчиняется логарифмическому закону и зависит от их начального количества.
Между количеством выживших микроорганизмов и продолжительностью нагревания, проведенного при постоянной температуре, существует зависимость, графическое изображение которой называют кривой выживаемости. Когда все клетки, составляющие популяцию, обладают одинаковой термостойкостью, зависимость количества выживших микроорганизмов от продолжительности нагревания носит экспоненциальный характер. Это означает, что при определенной температуре за единицу времени погибает одинаковая доля микроорганизмов от их общего количества. Абсолютное число погибших клеток при этом уменьшается со временем. Кривая выживаемости экспоненциального характера, построенная в полулогарифмической системе координат, приобретает вид прямой (рис. 18.1).
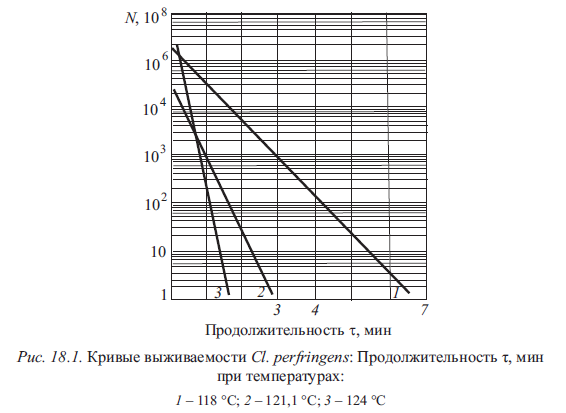
Экспериментально обнаружено, что для одних и тех же видов микроорганизмов кривая выживаемости строго экспоненциального характера (рис. 18.2, 4) не является единственной формой отражения зависимости между количеством выживших клеток или спор и продолжительностью нагревания. Возможны другие зависимости.
Кривые выживаемости 1, 2 (см. рис. 18.2) характерны для случаев, когда имеется увеличение или постоянство количества микроорганизмов в первые моменты нагревания по сравнению с их количеством до прогревания. Такое явление связано с возможной термоактивацией спор, находящихся в состоянии покоя до нагрева и выведенных из него тепловой обработкой.

Сигмоидальная и вогнутая кривые выживаемости характерны для случаев, когда в стерилизуемом материале клетки или споры обладают неодинаковой чувствительностью к нагреванию. В случае, когда кривая выживаемости составлена из двух прямых с разным уклоном, предположительно наличие в прогреваемой популяции двух форм спор, различающихся по устойчивости к нагреванию.
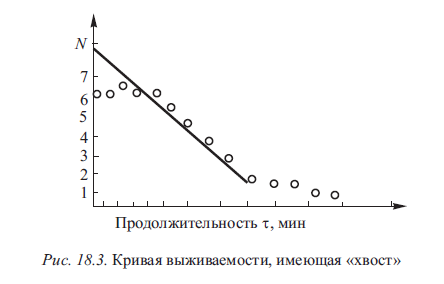
Экспериментально установлена еще одна форма отклонения кривой выживаемости от экспоненциального характера: в ней присутствует часть кривой, отражающей замедленное отмирание последних клеток, так называемый «хвост» кривой (рис. 18.3). К особенностям «хвостовых» клеток относится их неспособность давать подобное себе термостойкое поколение.
Константа скорости уничтожения конкретного вида микроорганизмов К зависит от агрессивного характера среды А, в которой микроорганизмы находятся (например, от активной кислотности, содержания фитонцидов и т.п.), от степени неустойчивости S данного вида микроорганизмов к температурному фактору и от температуры стерилизации Т, т.е.
![]()
Обозначим количество микроорганизмов определенного вида к началу стерилизации через В, количество микроорганизмов данного вида в конце стерилизации - b. Тогда уравнение (18.2) примет вид
![]()
Допустим, что имеются две банки с одинаковым пищевым продуктом, стерилизуемые в одних и тех же условиях, причем количество микроорганизмов в них в конце стерилизации b будет одинаково мало. Начальное количество микроорганизмов в одной из них В*, в другой - В**. Тогда смертельное время для микроорганизмов в первой банке
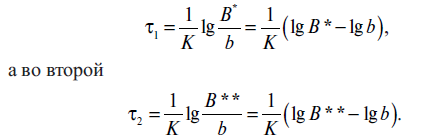
Отношение значений смертельного времени составит
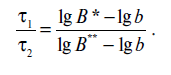
Поскольку величина b мала по условию, значением lgb можно пренебречь, и тогда уравнение (18.6) примет вид
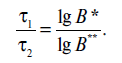
Таким образом, смертельное время зависит от начальной обсемененности продукта.
Из уравнения (18.3) можно определить время т, необходимое для уничтожения в данных условиях определенного количества микроорганизмов:
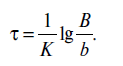
Выражение (18.10) тождественно выражению, связывающему температуру и время стерилизации, и, следовательно, ему должен отвечать аналогичный график в полулогарифмических координатах - график зависимости количества микроорганизмов от времени стерилизации при постоянной температуре. На горизонтальной оси откладывается смертельное время в линейных отрезках, на вертикальной - логарифмические значения количества микроорганизмов. На таком графике зависимость «количество микроорганизмов - смертельное время» характеризуется прямой линией (рис. 18.4). При этом В - количество микроорганизмов к началу стерилизации; b - количество микроорганизмов после стерилизации в течение т мин; т - время, необходимое для уничтожения микроорганизмов от В до b при заданной постоянной температуре стерилизации; D - смертельное время, требующееся для снижения количества микроорганизмов в 10 раз при заданной температуре. Иными словами, D (decimal reduction time) - время обработки, при котором погибает 90 % микроорганизмов, т.е. показатель, характеризующий тепловую устойчивость разных микроорганизмов при соответствующей температуре.
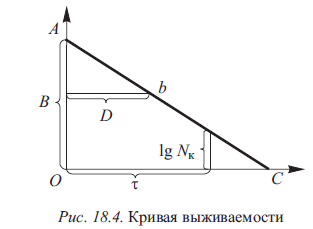
Приведенный на рис. 18.4 график отражает кинетику гибели микроорганизмов при постоянной температуре. Наклон прямой на рисунке характеризует термоустойчивость микроорганизмов в условиях такого прогревания, следовательно, соответствующей константой является тангенс угла наклона этой прямой, т.е. отношение вертикального катета АО к горизонтальному ОС.
Вертикальный катет на логарифмической шкале, означающий десятикратное снижение количества микроорганизмов, равен единице. Таким образом, l/D = tg a = соnst.
Но поскольку в этом отношении числитель (единица) на логарифмической шкале - тоже константа, следовательно, константой является и величина D, характеризующая устойчивость микроорганизмов к нагреванию в постоянном температурном поле (при постоянной температуре).
Чем больше значение константы термоустойчивости D, тем более устойчивыми являются микроорганизмы к температурному фактору. Константа термоустойчивости D характеризует, при прочих равных условиях, биологические особенности данного микроорганизма (табл. 18.3). Кроме того, Z-показатель, характеризующий температурную чувствительность микроорганизмов, можно представить как температурный диапазон, в котором происходит десятикратное изменение значения D.
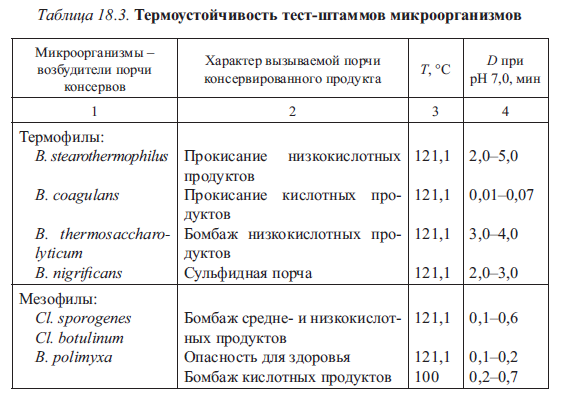

Константа D находится в обратной зависимости от агрессивного характера среды А, например от активной кислотности. Чем выше рН пищевого продукта, тем больше значение D. Пример изменения значения D для В. stearothermophilus в зависимости от рН приведен в табл. 18.4.

Для В. stearothermophilus установлена и эмпирическая зависимость между D и рН среды:
![]()
Термоустойчивость C1. sporogenes в низкокислотных продуктах определяют как
![]()
Константа D растет с понижением температуры стерилизации. Так, например, для С1. botulinum в «Икре из кабачков» (рН 4,8) она имеет следующие значения (в мин): при 121,1 °С - 0,044; при 118 °С - 0,086; при 115 °С - 0,216.
Выражение (18.10) в удобном для расчетов виде можно записать следующим образом:
![]()
Отсюда можно получить выражение для определения количества микроорганизмов в конце стерилизации:
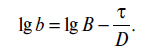

Дробное (меньше единицы) количество микроорганизмов после стерилизации означает, что споры будут обнаруживаться не в каждой банке, а для данного случая - в одной из 10 000 банок.
Из сказанного вытекает, что, учитывая логарифмический характер гибели микроорганизмов при нагревании, теоретически полностью уничтожить их при стерилизации невозможно. Можно уменьшать количество спор при нагревании, доводя их до одной на 1000, на 1 млн банок и т.д., но не уничтожая на 100 %. Следовательно, можно говорить не об абсолютной стерильности, а только о степени стерильности.
Точно так же нельзя говорить о смертельном времени как о времени полного уничтожения микроорганизмов. Смертельным временем следует называть время, необходимое для достижения заданной степени стерильности, т.е. для доведения первоначального количества микроорганизмов до заранее установленного низкого уровня.
Из уравнения (18.6) видно, что при данных условиях стерилизации (т.е. для данного продукта при определенной температуре и времени стерилизации) количество выживших к концу процесса микроорганизмов b прямо пропорционально их начальному содержанию В. Таким образом, чем меньше микроорганизмов в пищевом продукте к началу стерилизации, тем меньше их в консервах к концу процесса. Это очень важный в практическом отношении вывод, говорящий о необходимости поддержания высокого санитарно-гигиенического уровня на консервных заводах.
