Основные изменения в молоке после внесения в него молокосвертывающих энзимов показаны на рис. 2.13. Отщепление от мицеллы МП сопровождается снижением ее заряда примерно в два раза, уменьшением радиуса мицеллы примерно на 5 нм и вязкости молока; гидродинамический объем мицеллы снижается до 2/3 от ее величины для нативной мицеллы; количество связанной мицеллой воды при pH 6,6 уменьшается почти в 1,5 раза. Энзим как бы «сбривает» волосообразный внешний слой мицеллы казеина (разд. 2.3). Снижение поверхностного заряда ослабляет электростатические, а удаление части гидратной оболочки и внешнего слоя, образуемого МП, - стерические (пространственные) факторы стабилизации мицелл, в результате чего мицеллы теряют устойчивость.
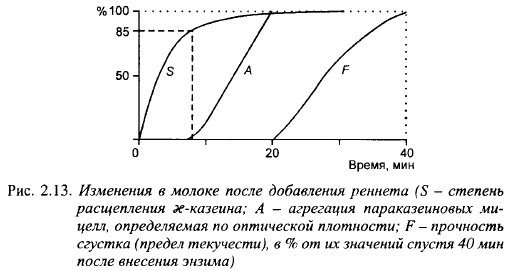
Гидролиз æ-казеина начинается сразу после внесения в молоко молокосвертывающих энзимов. Визуально, свертывание молока становится заметным после расщепления 80-85% æ-казеина (точка К на рис. 2.12), т. е. вторая фаза начинается до окончания первой (энзиматической) фазы. Начало флокуляции характеризуется повышением вязкости. Следует отметить, что небольшое изменение вязкости, улавливаемое вискозиметрами с чувствительностью более 0,01 Па-с, происходит во время индукционного периода: после внесения энзима вязкость снижается, что сопровождается небольшим увеличением степени дисперсности казеиновых частиц, затем вязкость повышается примерно до начальной величины к концу индукционного периода. Раманаускас объясняет первоначальное снижение вязкости распадом внутренних упорядоченных структур казеиновых мицелл, существовавших до внесения в молоко молокосвертывающих энзимов.
Повышение вязкости, начинающееся примерно с середины индукционного периода, он связывает с началом агрегации параказеиновых мицелл. Таким образом, он считает, что первая и вторая фазы сычужного свертывания идут одновременно, по крайней мере, со второй половины первичной фазы. Ранее эта же мысль была высказана голландским ученым Пайенсом, который считал, что флокуляция идет сразу после внесения в молоко молокосвертывающего энзима со все возрастающей скоростью; нарастание скорости обусловлено увеличением количества параказеиновых мицелл и числа эффективных соударений мицелл друг с другом.
Если принять среднюю молекулярную массу мицеллы казеина равной 8·108, содержание æ-казеина в мицелле - 11,9% и молекулярную массу его - 19000, то на каждую мицеллу приходится 5100 молекул æ-казеина. Индивидуальные мицеллы могут связываться с другими мицеллами после расщепления 97% входящего в их состав æ-казеина, т. е. после разрыва примерно 5000 связей. Для образования комплекса из двух мицелл нужно расщепить 10 тыс. связей. При обычно используемой дозе химозина в сыроделии на одну молекулу энзима приходится около ста мицелл казеина. Ясно, что для подготовки их к флокуляции нужно время. Это и объясняет некоторое запаздывание начала второй фазы относительно первой. Кроме того, повышение вязкости молока при незначительном числе агрегированных мицелл не улавливается современными методами. Правильнее считать, что энзиматическое свертывание молока состоит не из двух фаз, а двух процессов, идущих параллельно, но с некоторым запозданием процесса афегации параказеиновых мицелл. Точка же К - не начало, а самый разгар флокуляции, когда образовавшиеся комплексы параказеиновых мицелл становятся видимыми невооруженным глазом. Под микроскопом начало флокуляции можно увидеть значительно раньше. С помощью нефелометрических методов McMahon et al. также установили, что флокуляция начинается значительно раньше появления хлопьев, наблюдаемых визуально.
Энзиматическую и неэнзиматическую фазы сычужного свертывания можно разобщить, используя температурный фактор: первая фаза сычужного свертывания идет даже при 0° С, но обязательно в присутствии молокосвертывающего энзима, а вторая фаза идет без участия молокосвертывающих энзимов, но только в присутствии ионов Са, и прекращается при температуре ниже 15° С. Это различие используют на практике. Для получении сгустка непрерывным способом в производстве некоторых сыров (например, Камамбера) первую фазу проводят при низких температурах, затем молоко в теплообменниках непрерывного действия нагревают до температуры 30-34° С, при которой оно быстро свертывается; образовавшийся сгусток направляют в сырные формы для самопрессования.
Проведение первой фазы при низких температурах увеличивает продолжительность выдержки молока с молокосвертывающими энзимами и позволяет снизить их дозы, однако этот прием редко применяют в промышленности, так как длительная выдержка молока с энзимом даже при низких температурах может отрицательно сказаться на выходе и качестве сыра из-за размножения психротрофной микрофлоры и бактериофагов микрофлоры заквасок. Такой метод свертывания уменьшает степень протеолиза и ухудшает качество сыра за счет снижения выраженности сырного вкуса, появления более плотной, иногда мучнистой консистенции. По-видимому, снижение времени выдержки геля при температуре свертывания не дает возможности окончательно сформироваться его структуре и замедляет накопление биомассы микрофлоры заквасок, которая будет достигать максимума в сыре позднее, чем обычно, когда температура для действия ее энзимов будет менее благоприятна.
Кинетика гидролиза æ-казеина молокосвертывающими энзимами подчиняется закону Михаэлиса-Ментен. В соответствии с этим для нее будет справедливо уравнение:


