Клетки лактобацилл имеют размеры 0,5-1,2x1,0-10 мкм. Как правило, длинные, но иногда почти кокковидные, обычно в коротких цепочках. В редких случаях подвижные. Факультативные анаэробы, иногда микроаэрофилы; лучше растут при пониженном содержании кислорода. Рост обычно стимулируется 5% С02. Типовой вид Lactobacillus delbrueckii.
По характеру вызываемого ими брожения лактобациллы можно разделить на три группы.
Группа 1. Облигатные гомоферментативные микроорганизмы. Сбраживают гексозы почти исключительно до молочной кислоты, пентозы и глюконат не сбраживают. С02 не образуют. Из наиболее важных для сыроделия видов лактобацилл к этой группе принадлежат две подгруппы, сформированные на основе генетического родства: подгруппа Lbc. delbrueckii с подвидами delbrueckii, lactis и bulgaricus (далее Lbc. debrueckii, Lbc. lactis и Lbc. bulgaricus) и подгруппа Lbc. helveticus и Lbc. acidophilus.
Группа 2. Факультативные гетероферментативные микроорганизмы. Представители этой группы сбраживают гексозы по гомофермента- тивному пути, а пентозы по гетероферментативному с образованием молочной и уксусной кислот. К этой группе из важнейших для сыроделия микроорганизмов принадлежит Lbc. casei с подвидами.
Группа 3. Облигатные гетероферментативные лактобациллы. Представители этой группы сбраживают гексозы до молочной и уксусной кислот (или этанола) и С02. В эту группу входят Lbc. brevis и Lbc. fer- mentum, часто встречающиеся в сырах.
В сыроделии находит применение Lbc. plantarum-, этот вид по характеру метаболизма и фенотипическим характеристикам должен входить во вторую группу, а генетически он тесно родственен Lbc. brevis.
Среда обитания
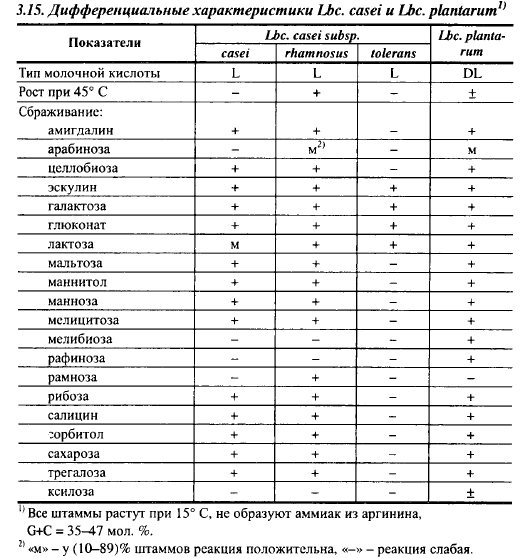
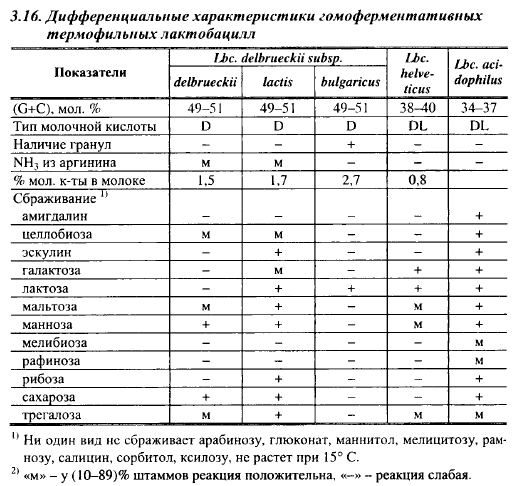
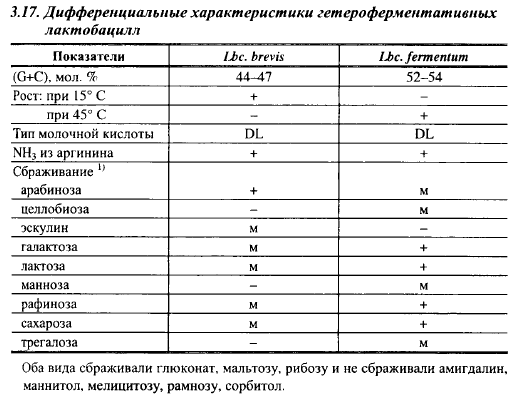
Дифференциальные характеристики наиболее важных для сыроделия видов лактобацилл приведены в табл. 3.15, 3.16 и 3.17.
Лактобациллы отличаются очень сложными питательными потребностями и поэтому могут жить только в субстратах, богатых органическими питательными веществами: растворимыми углеводами, продуктами протеолиза (низкомолекулярными пептидами, свободными аминокислотами), жирными кислотами, витаминами, производными нуклеиновых кислот. Вторым их требованием к среде обитания является низкое содержание в среде кислорода или его отсутствие. Таких мест в природе не так много. Наиболее массивными источниками лактобацилл являются силос, кишечник млекопитающих, хотя их доля в кишечной микрофлоре невелика (104— 109 КОЕ/г при общем содержании бактерий 101 -1011 КОЕ/г), молочные предприятия, где лактобациллы применяются в производстве большинства ферментированных продуктов. Несмотря на это, в небольших количествах лактобациллы обнаруживаются повсеместно, чему способствует их устойчивость к внешним факторам, в частности к pH, у многих видов - к осмотическому давлению, способность размножаться в широком диапазоне температур и высокая антагонистическая активность по отношению к другой микрофлоре.
Молоко при выходе из вымени не содержит лактобацилл, но быстро обсеменяется ими уже на молочных фермах. В молоке высокого бактериального качества от одного стада содержится от 1 до 50 клеток/мл лактобацилл, что намного меньше, чем других групп молочнокислых бактерий или соизмеримо с их количеством. В сыром сборном молоке при поступлении на молочные заводы обычно содержится от 103 до 2104 клеток/мл лактобацилл, но летом их количество может достигать до 105/мл. В сыром молоке доминируют Lbc. casei и Lbc. plantarum, изредка встречаются Lbc. brevis, Lbc. fer- mentum и другие виды. Количество термофильных лактобацилл в сборном молоке составляет 30-50 клеток/мл.
Лактобациллы обнаруживаются в любом сыре, независимо от их наличия в заквасках. Основным источником обсеменения лактобациллами сыров, вырабатываемых на лактококковых заквасках, является внешняя среда на самом заводе.
Мезофильные лактобациллы
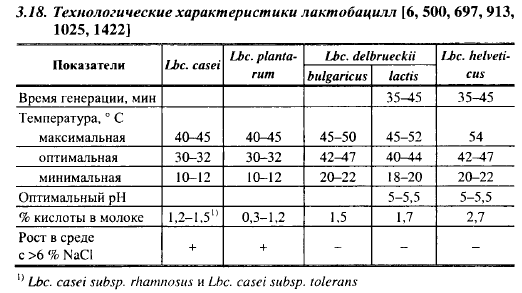
Рост в молоке и сыре. К мезофильным относят лактобациллы, которые растут при 15° С и обычно не растут при 45° С (рис. 3.2). К гомоферментативным или факультативным гетероферментативным видам принадлежат Lbc. casei с подвидами и Lbc. plantarum, к гетероферментативным - Lbc. brevis и Lbc. buchneri, которые отличаются только отношением к мелицитозе: первый микроорганизм сбраживает, второй не сбраживает мелицитозу. Диапазон температур для роста гомо- ферментативных видов показан в табл. 3.18. Оптимальная температура для роста Lbc. brevis равна 30° С, максимальная - 38° С.
Мезофильные лактобациллы отличаются довольно высокой устойчивостью к тепловой обработке: нагревание при 65° С в течение 30 мин выдержали 93% штаммов Lbc. plantarum и 97% штаммов Lbc. casei, хотя сохранившие жизнеспособность после такой обработки клетки на какое-то время теряют активность. В сырах их активность постепенно восстанавливается. Таким образом, молоко является постоянным источником обсеменения сыров лактобациллами. Количество клеток лактобацилл, попадающих в сыр этим путем, зависит от их содержания в сыром молоке и режима тепловой обработки молока. По голландскому стандарту в пастеризованном молоке для выработки сыра должно содержаться менее одной клетки лактобацилл/мл, что возможно, если в молоке перед пастеризацией содержалось не более 50 клеток/мл этих бактерий.
Основным источником лактобацилл в сыре является загрязнение молока после пастеризации, чаще всего через недостаточно хорошо вымытые и продезинфицированные молокопроводы, оборудование, инвентарь. Лактобациллы могут попадать непосредственно в сыр из рассола во время посолки. При современной технике производства выработать в промышленных условиях сыр, не содержащий лактобацилл, невозможно.
Мезофильные лактобациллы, по крайней мере часть штаммов, не только выдерживают II нагревание при выработке мелких сычужных сыров, но способны размножаться на этом этапе. Из 30 свежевыделенных из растительных субстратов штаммов Lbc. plantarum один рос при 45° С, 10 - при 43° С. Нижние температурные границы для их роста совпадают с нижними температурами созревания отечественных мелких сыров. В табл. 3.18 указана минимальная температура для роста большинства штаммов мезофильных лактобацилл, равная 10° С, но некоторые штаммы очень медленно утилизируют цитраты и при 8° С. Таким образом, с точки зрения температуры, мезофильные лактобациллы могут расти на любом этапе производства мелких сычужных сыров, хотя во время созревания довольно медленно. Повышение температуры созревания в интервале от 10 до 15° С увеличивает скорость размножения и выход биомассы лактобацилл в сырах.
Температуры II нагревания при производстве крупных сыров выше максимальных для роста мезофильных лактобацилл, но ниже температур, которые вызывают их гибель. Они сохраняют жизнеспособность после двухчасовой выдержки при 55° С. В финском блочном Эмменталь- ском сыре температура сырной массы в середине блока снижается до допускающего рост мезофильных лактобацилл уровня за 5 ч, а на расстоянии 6 см от поверхности блока - за 13 ч до окончания прессования. Следовательно, мезофильные лактобациллы могут размножаться в крупных сырах уже во время прессования. Условия же созревания крупных сыров (температура, pH, активность воды) более благоприятны для роста этих микроорганизмов, чем в мелких сырах. Таким образом, и в сырах с высокими температурами И нагревания температурный режим не препятствует размножению мезофильных лактобацилл на всех этапах производства, кроме II нагревания и первой половины прессования.
Штаммы мезофильных гомоферментативных лактобацилл очень различаются по скорости кислотообразования и предельной кислотности в молоке. Остроумова разделила 30 штаммов Lbc. plantarum и 13 штаммов Lbc. casei по приросту кислотности молока при 30° С и 1 %-ной посевной дозе на три группы, дающих прирост кислотности (°Т), соответственно, 1,0-1,3; 2,4—2,6; 4,7-7,3 через 6 ч и 18-19; 30-34; 46-47 через 24 ч инкубации. Сильные кислотообразователи сквашивали молоко в этих условиях через 1,2-1,4 сут, слабые - за 4,5-6,0 сут. Лактококки, за исключением штаммов диацегильного лактококка с низкой скоростью кислотообразования, при 30° С и 1 %-ной посевной дозе сквашивают молоко менее чем за 24 ч. Таким образом, скорость развития лактококков в молоке значительно выше скорости роста мезофильных гомоферментативных лактобацилл. Обратная картина наблюдалась с предельной кислотностью в молоке: в опытах Макаровой она равнялась для штаммов Lbc. plantarum (101-147)° Т; для Lbc. casei - (154-170)° Т, что выше предельной кислотности лактококков (110-120)° Т. Это свидетельствует о высокой кислотоустойчивости лактобацилл. Они могут расти в молоке при pH ниже 3,8. Подобные результаты получены другими авторами со штаммами Lbc. plantarum. Они близки к данным зарубежных авторов, обобщенным Квасниковым и Нестеренко (табл. 3.18).
Большой разброс данных в пределах вида можно объяснить происхождением штаммов - часто свежевыделенные из природных источников, в том числе из сыра, штаммы мезофильных лактобацилл медленно образуют кислоту в молоке и могут его не свертывать, но после продолжительного выращивания в молочных средах их активность в молоке повышается.
Недиссоциированная молочная кислота вызывает гибель клеток Lbc. casei при концентрации более 94 мМ (8,46%), что выше, чем содержание лактатов в водной фазе сыров [1430], и выше, чем допустимые концентрации лактатов для роста лактококков. Таким образом, содержание в сырах лактатов ниже уровня, препятствующего размножению ме- зофильных лактобацилл. Следует отметить, что ингибирование роста Lbc. delbrueckii начинается в среде с 40-60 г/л молочной кислоты.
Скорость роста в молоке мезофильных гомоферментативных лактобацилл тесно коррелирует с их протеиназной способностью. Многие штаммы мезофильных лактобацилл слабо атакуют казеин. Обогащение молока продуктами гидролиза белков и факторами роста ускоряет рост неактивных в молоке штаммов молочнокислых бактерий. Плотность популяции Lbc. plantarum в двенадцатичасовых культурах в зависимости от типа белкового гидролизата, вносимого в молоко для стимуляции их роста, колеблется в пределах от 785 до 1605 млн. КОЕ/мл. При этом некоторые штаммы этого вида лучше росли в средах, в которых необходимые им аминокислоты находились не в свободном виде, а в составе низкомолекулярных пептидов. Оптимальный для роста лактобацилл pH находится в интервале 5,5-6,0, что значительно ниже, чем pH свежего молока.
Лактококки в молоке стимулируют развитие лактобацилл путем образования низкомолекулярных азотистых соединений из казеина и других факторов роста, снижения pH и Eh. Интересно, что и в сырах мезофильные лактобациллы при их введении в состав заквасок не увеличивают содержание водорастворимых белков, повышая количество продуктов более глубокого расщепления белков. Чем выше казеолитическая активность лактококков закваски, тем быстрее размножаются мезофильные лактобациллы в сыре. В совместных с лактококками культурах в молоке выход биомассы Lbc. casei увеличился на 39%, протеолитическая активность по отношению к аs1казеину возросла в 5,8 раза. При автолизе клеток лактококков также высвобождаются вещества, стимулирующие рост лактобацилл.
Штаммы Lbc. plantarum при введении в закваску в количестве 5-10% от количества лактококков повышают общее содержание жизнеспособных клеток бактерий в закваске в 1,33-1,5 раза, особенно сильно стимулируя рост диацетильного лактококка и лейконостоков. Lbc. casei повышает кислотообразующую активность в совместных с лактококком культурах путем образования валина, лейцина и пептидов.
Влияние лактобацилл на развитие лактококков в смешанных культурах в значительной степени зависит от их исходного соотношения и свойств штаммов. При достаточно высокой дозе штаммы лактобацилл с высокой скоростью кислотообразования вытесняют лактококки благодаря большей устойчивости к pH [646].
Накопление биомассы микроорганизмами в сыре пропорционально их исходному количеству, удельной скорости и времени размножения.
В молоко вносят с закваской при выработке мелких сыров около 107 КОЕ/мл лактококков, что примерно в миллион раз больше, чем исходное содержание лактобацилл. Лактококки размножаются во время выработки и в первые дни созревания сыра, достигая максимума в 5-10-суточных сырах, равного примерно 2109 КОЕ/г, т. е. количество их клеток по отношению к исходному количеству в смеси возрастает примерно в 200 раз. Параллельно с ними размножаются лактобациллы. При одинаковой интенсивности размножения лактококков и лактобацилл в сыре содержание лактобацилл тоже должно увеличиться примерно в 200 раз и составить примерно 2-103 КОЕ/г. На самом деле при нормальном развитии микробиологических процессов в мелких сырах содержание лактобацилл в сыре составляет не менее 2106 КОЕ/г, т. е. отношение между лактококками и лактобациллами в сыре в период их максимального содержания становится равным примерно 1000:1, т. е. оно уменьшилось по сравнению с исходным примерно в 1000 раз. Это говорит о том, что в мелких сырах условия для роста лактобацилл поддерживаются более продолжительное время, чем для лактококков.
Прекращение увеличения содержания жизнеспособных клеток лактококков в сыре обычно совпадает с изчезновением лактозы в сыре или происходит немного раньше этого момента. Однако истощение лактозы не единственная причина прекращения накопления жизнеспособных клеток лактококков в сыре, поскольку в экстрактах трехмесячного сыра, обогащенных лактозой и дрожжевым экстрактом, их рост не вознобновляется. Скорее всего рост лактококков в сырах прекращается в результате воздействия комплекса неблагоприятных факторов, о чем сказано в разд. 3.2.4.
С прекращением увеличения содержания биомассы лактококков заканчивается лактококковая фаза молочнокислого брожения в сыре, но продолжается фаза развития молочнокислых палочек. Интересно, что двухфазность характерна и для спонтанного молочнокислого процесса в сыром молоке и силосе. Одна причина продолжения роста лактобацилл в сыре после прекращения роста лактококков понятна - это их более высокая кислотоустойчивость. Встает вопрос об источниках энергии для их роста. На этот вопрос отвечает новозеландский ученый Thomas. В период максимума в мелких сырах насчитывается (2-3)Т09 КОЕ/г лактококков, что эквивалентно 1-3 мг сухого веса бактерий/г. Под действием собственных энзимов клетки лактококков постепенно лизируются. В модельных условиях, имитирующих условия в сыре, лактобациллы и педиококки используют продукты автолиза клеток микрофлоры заквасок в качестве источников энергии.
Возможны и другие источники энергии, в частности лактаты, количество которых в сырах снижается к концу созревания. Lbc. plantarum после длительного культивирования в среде с глюкозой может трансформировать лактаты в ацетаты в присутствии 02, но нет доказательств, что они могут это делать в сырах. Содержание лактатов в мелких сырах в конце созревания всегда ниже, чем в период максимума, даже при отсутствии их ферментации пропионовокислыми и маслянокислыми бактериями.
Mabbitt & Zielinska (1956) считают, что даже в зрелых сырах сохраняются небольшие количества лактозы и галактозы (около 0,15%), которые не могут быть выявлены обычными методами. По Перфильеву и Рогову, развитие лактококков лимитируется при содержании в среде меньше 0,3% лактозы и увеличении концентрации лактатов больше 2%. Галактоза в мелких сырах, как продукт гидролиза лактозы термофильным стрептококком, который размножается в этих сырах во время II нагревания и прессования, всегда может присутствовать. Количество термофильных стрептококков в мелких сырах варьирует, особенно в зависимости от соблюдения правил эксплуатации пастеризационных установок. В сырах из сырого молока галактоза забродила на 25-е сутки, из пастеризованного молока - на 53. Это показывает, что галактозу в сырах сбраживает незаквасочная микрофлора. Показательно, что из твердых, мягких и полумягких сыров были выделены в основном Lbc. plantarum и Lbc. casei subsp. casei, хотя штаммы последнего подвида или не сбраживают, или медленно сбраживают лактозу, но активно сбраживают галактозу.
Прекращается рост мезофильных лактобацилл в сыре после использования доступных источников энергии, так как при добавлении в экстракты трехмесячного сыра лактозы рост в них Lbc. casei вознобновлялся.
На рис 3.7 показано развитие мезофильных лактобацилл в Костромском и Российском сырах со специальным внесением и без внесени- ия в смесь для их выработки этих микроорганизмов. В кривых, характеризующих изменение содержания мезофильных лактобацилл в сырах без внесения в смесь (лактобациллы в этих сырах представлены «дикими» штаммами), есть перегибы, обычно отражающие временную остановку размножения культур при переходе на другой источник энергии. По времени они совпадают с изчезновением в сырах лактозы. Поскольку в сырах этого возраста автолиз микрофлоры заквасок еще незначителен, лактобациллы в этот момент, очевидно, переходят от сбраживания лактозы к сбраживанию галактозы. Lbc. casei и Lbc. plantarum сбраживают галактозу, тогда как скорость роста лактококков резко снижается при содержании в среде менее 30 мМ галактозы. Содержание галактозы в мелких сырах колеблется в зависимости от интенсивности размножения термофильного стрептококка. В сырах с высокими температурами II нагревания мезофильные лактобациллы могут быть лучше обеспечены источниками энергии, чем в мелких сырах. Это обусловлено большей интенсивностью размножения в них термофильного стрептококка, неспособностью микрофлоры заквасок сбродить всю лактозу из-за гибели лактококков во время II нагревания и слишком быстрого остывания сырной массы до температуры ниже минимальной границы для роста термофильной микрофлоры. Это особенно касается Советского сыра, имеющего меньшие размеры головки и более короткий срок прессования, чем другие представители сыров этого класса.
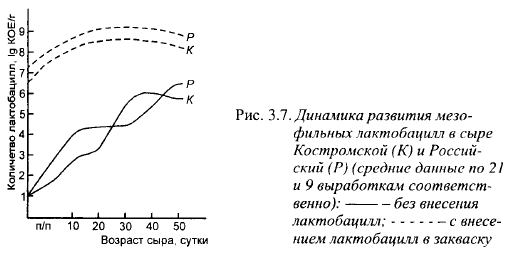
Перегиб кривой изменения содержания лактобацилл в Российском сыре начался раньше, чем в Костромском, что может быть обусловлено более быстрым сбраживанием лактозы в Российском сыре.
Из-за низкого исходного количества и сравнительно небольшой скорости размножения в молоке и сырах наиболее интенсивное накопление биомассы лактобацилл происходит во второй половине созревания, когда соль уже проникла в глубину головки сыра. При содержании в сырах 2% соли и 40% влаги, содержание соли в водной фазе сыра составляет около 4,8%. Штаммы мезофильных лактобацилл, способные размножаться при высоких концентрациях соли, широко распространены в природе, в частности, они являются постоянными обитателями солильных бассейнов [463]. Они растут в молоке с 9-12% соли. Штаммы мезофильных лактобацилл, выделенные из сыров, росли в средах с 12% соли (Smith & Gunnin- gamham, 1962). Следовательно, концентрация соли в сырах не является препятствием для роста мезофильных лактобацилл. Следует отметить, что при концентрации в среде 2, 4 и 6% NaCl рост 103 штаммов Lbc. plan- tarum в среднем замедлялся на 35, 59 и 74% соответственно. Только некоторые штаммы Lbc. brevis могут расти в средах с 6,5% соли.
При нормальном развитии микробиологических процессов масштабы размножения лактобацилл в мелких сырах, вырабатываемых на лактококковых заквасках, невелики: по голландским стандартам в сырах 2-недельного возраста должно быть не более 2106 КОЕ/мл лактобацилл. Примерно такой их уровень наблюдается в отечественных сырах при нормальном развитии молочнокислого брожения. Положение может измениться при снижении активности лактококковой микрофлоры заквасок. В этом случае большая доза лактозы достанется мезофильным лактобациллам и максимальный выход их биомассы может возрасти до нескольких сотен миллионов КОЕ/г. Содержание лактобацилл в сыре Чеддер в период максимума колеблется от 3106 до 9-108 КОЕ/г.
Мезофильные лактобациллы активно размножаются и в крупных сырах. В Советском сыре промышленных выработок после прессования их было 2-106 КОЕ/г, в течение 20-суточной выдержки сыров при 10-12° С количество мезофильных лактобацилл увеличилось до 2107 КОЕ/г, после 25-суточной выдержки в теплой камере при 22-24° С оно возросло до 8,4-107, затем во время созревания сыров при 10-12° С количество жизнеспособных клеток мезофильных лактобацилл постепенно снижалось и в 90-суточных сырах составило 5,9107 КОЕ/г. Максимальное содержние мезофильных превысило максимальное содержание термофильных лактобацилл в 1,22 раза. Из Эмментальского сыра, выработанного на трех сыродельных заводах, было выделено 609 штаммов лактобацилл, из которых 68,31% были мезофильными гомоферментативными видами, 19,21% термофильными и 9,03% гетероферментативными видами. Любопытно, что в итальянском сыре Пекорино, вырабатываемом при температуре II нагревания 45—48° С на естественных (сывороточных) заквасках, уже на ранних стадиях созревания из лактобацилл преобладают Lbc. casei и Lbc. plantarum. Lbc. casei преобладала в сыре Пармезан во время созревания. При испытании заквасок различного видового состава для выработки сыра Рас (Египет) со средней температурой II нагревания (44° С), по содержанию влаги (30-34%) близкому к терочным сырам, лучшие результаты дала комбинация Lbc. casei с болгарской палочкой или закваской для йогурта.
В сыре Рокфор мезофильные лактобациллы составляют от 1/3 до 1/8 общего количества бактерий: до посолки в нем размножался в основном Lbc. casei, после посолки - штаммы Lbc. plantarum, способные расти в средах с более чем 10% соли.
В сыре Качкавал, вырабатываемом с чеддеризацией и плавлением сырной массы, в трехмесячном возрасте содержание мезофильных лактобацилл было в 10-1000 раз выше, чем содержание лактококков. Из него было выделено и идентифицировано 439 культур лактобактерий, из которых 89,1% были гомоферментативными мезофильными лактобациллами.
Из испанского твердого сыра с низкой температурой II нагревания из овечьего молока Манчего на разных стадиях созревания выделено 296 штаммов лактобацилл, из которых 134 (45,3%) идентифицировано как Lbc. casei, 148 (50%) - как Lbc. plantarum, 5 (1,7 %) - как Lbc. brevis.
Таким образом, и в мелких, и в крупных сычужных сырах имеются ниши для мезофильных лактобацилл. Преобладающими видами мезофильных лактобацилл в мелких сырах являются Lbc. casei и Lbc. plantarum, постоянно встречаются в них представители гетероферментативного вида Lbc. brevis, но в значительно меньших количествах.
Роль в мелких сырах. По количеству биомассы лактобациллы занимают в мелких сырах второе место после лактококков. Резонно полагать, что они должны оказывать существенное влияние на органолептические показатели сыров. Выработки сыров с низкой температурой II нагревания и Чеддера в асептической ванне в большинстве случаев показали, что лактобациллы при внесении в закваску вместе с лактококками положительного влияния на качество сыра не оказывали. Более того, они иногда ухудшали органолептические показатели сыра. Исходя из этого, авторы этих опытов не рекомендуют включать мезофильные лактобациллы в закваски для выработки сыров с низкими температурами II нагревания, хотя сыры, вырабатываемые в открытой ванне, в которых всегда есть «дикие» штаммы лактобацилл, как правило, быстрее созревают и имеют более выраженный вкус, чем сыры, вырабатываемые в асептической ванне с участием только лактококков. Неоднозначны и результаты экспериментов по выработке сыров в асептической ванне. В опытах ирландских ученых с выработкой сыров в асептической ванне внесение в молоко специально отобранных штаммов лактобацилл увеличило содержание молочнокислых палочек в сыре по сравнению с контролем (сыры, вырабатываемые только с участием лактококков) на 3-5 порядков и сделало вкус сыра более приемлемым для потребителя.
По-видимому, выводы большинства авторов по экспериментам с выработками сыров в асептической ванне относительно роли мезофильных лактобацилл в мелких сырах требуют корректировки с учетом результатов исследований сыров, вырабатываемых в обычной ванне и производственных условиях. Имеются теоретические предпосылки и экспериментальные доказательства важной роли мезофильных лактобацилл в сырах.
Выше показано, что лактококки не могут достаточно быстро сбродить все углеводы в сыре. Мезофильные лактобациллы сбраживают углеводы, остающиеся после окончания размножения лактококков, и тем самым лишают технически вредную и патогенную микрофлору энергетических источников. Примерами технически вредной микрофлоры являются гетероферментативные лактобациллы (Lbc. brevis, Lbc. buchneri, Lbc. fermentum), способные вызвать в сырах броженый, дрожжевой привкусы и образовывать токсичные амины при длительном созревании или хранении сыров. При выработке сыров в асептической ванне эта положительная роль лактобацилл не реализуется из-за отсутствия в сыре посторонней микрофлоры.
Слишком высокая скорость кислотообразования в сырах, вырабатываемых на комбинированных заквасках (лактококки + лактобациллы), является одним из препятствий широкого применения мезофильных лактобацилл в сыроделии. Однако, еще больший ущерб сыроделию приносит недостаточная скорость молочнокислого брожения во время выработки сыров, что может быть обусловлено действием бактериофагов, низким качеством молока, нередко наблюдаемым весной.
Мезофильные гомоферментативные лактобациллы нечувствительны к бактериофагам лактококков, стимулируют развитие лактококков в совместных культурах, поэтому включение в закваску специально отобранных штаммов мезофильных лактобацилл, заведомо не образующих пороки в сырах, может в определенной степени повысить стабильность молочнокислого брожения при выработке сыра, которая является необходимым условием получения сыров высокого качества. В асептически выработанных сырах эта роль лактобацилл также не проявляется из-за отсутствия фага.
Чем быстрее будет сброжена в сыре лактоза, тем меньше шансов у технически вредной и патогенной микрофлоры размножиться до опасного для качества сыра уровня. В сырах типа Российский для ускорения молочнокислого брожения увеличена продолжительность прессования с таким расчетом, чтобы после прессования его pH был на уровне 5,1-5,25 (во время прессования сыр остывает медленнее, чем во время посолки, что обусловливает более длительное сохранение на высоком уровне скорости молочнокислого брожения). Этот прием повышает стойкость сыра по отношению к вредной микрофлоре только при условии нормального размножения микрофлоры заквасок во время прессования сыра. При низкой активности молочнокислого процесса стойкость к вредной микрофлоре сыров с продолжительным прессованием ниже, чем сыров с обычной продолжительностью прессования, поскольку более медленное остывание сырной массы во время прессования при неактивном молочнокислом процессе ускоряет размножение нежелательной микрофлоры.
Изучено влияние внесения в молоко, наряду с лактококковой закваской, 1% закваски Lbc. plantarum или 0,04 % закваски Lbc. acidophilus на скорость молочнокислого брожения и развитие БГКП в Российском сыре. Если в контрольных сырах, вырабатываемых с лактококковой закваской, pH во время прессования снижался до заданного уровня за 6-7 ч, то в опытных сырах с включением в состав закваски Lbc. plantarum для этого требовалось 4-5 ч, в сырах с включением в закваску Lbc. acidophilus - 3-4 ч. Влияние видового состава закваски на развитие в Российском сыре БГКП показано на рис. 3.8.
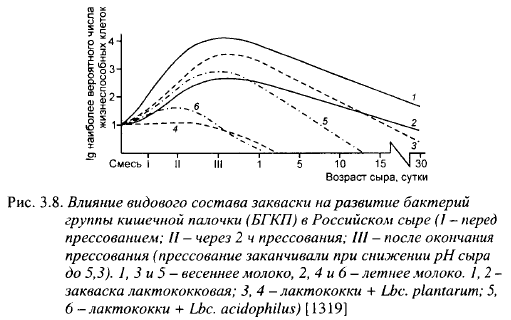
Комбинированный бакпрепарат лактококков и специально подобранных штаммов Lbc. plantation был использован для выработки Российского сыра на нескольких сырзаводах весной. В качестве контроля вырабатывали сыр на той же лактококковой закваске, но без добавления Lbc. plantarum. Характеристика опытной и контрольных заквасок приведена в табл. 3.19. Закваска с лактобациллами имела более высокую кислотообразующую активность и скорость накопления, биомассы, что отразилось на процессе выработки сыра: прирост кислотности сыворотки в опытных сырах был выше, чем в контрольных, на 1,7° Т, содержание влаги и pH после прессования в контрольных и опытных сырах равнялись соответственно 46 и 44,8; 5,4 и 5,25. Таким образом, введение в состав производственной закваски Lbc. plantarum позволило нормализовать технологический процесс и выработать Российский сыр из весеннего молока с оптимальными содержанием влаги и кислотностью. Общая оценка опытных сыров была в среднем на 1,7 балла выше, чем контрольных, в т. ч. на 1,2 балла за вкус и запах и на 0,5 балла за консистенцию.

Введение в закваски достаточно больших количеств Lbc. plantarum для стабилизации скорости молочнокислого процесса можно рекомендовать для производства сыров с повышенным уровнем молочнокислого брожения, для которых характерен более кислый вкус и специфичные структура (отсутствие правильного рисунка) и консистенция. В опытах Lee et al. использование некоторых штаммов Lbc. casei, наряду с обычной закваской, позволило выработать сыр Чеддер высокого качества.
Преобладание мезофильных лактобацилл в сырах типа Качкавал можно объяснить гибелью стрептококков во время плавления сырной массы и неспособностью термофильных лактобацилл размножаться при температурах созревания Качкавала. Таким образом, в сырах типа Качкавал мезофильные лактобациллы необходимы для выполнения одной из главных функций молочнокислых бактерий в сырах - сбраживания углеводов. Это относится и к рассольным сырам, в которых при использовании только лактококковой закваски углеводы обнаруживаются вплоть до 100-суточного возраста. Уместно напомнить, что при наличии углеводов стафилококки могут размножаться в анаэробных условиях при концентрациях соли, характерных для рассольных сыров. При выработке рассольных сыров с заквасками, содержащими солеустойчивые штаммы лактококков и Lbc. casei, увеличились скорости сбраживания лактозы, цитратов и протеолиза и повысилось качество сыров.
Многие авторы отмечают ускорение созревания, смягчение консистенции, уменьшение горечи в сырах под действием гомоферментативных мезофильных молочнокислых палочек. Это может быть связано с особенностями их протеолитической системы, характеризуемой невысокой скоростью расщепления казеина, но высокой ами- нопептидазной активностью. Перенос фрагмента ДНК, кодирующего аминопетидазную активность лактобацилл, в Lc. lactis subsp. cremoris и применение полученного реципиента для выработки сыра в 1000 раз увеличило аминопептидазную активность в Чеддере, но не ускорило формирование сырного вкуса. Однако в сырах, выработанных с полученными трансформантами, не было горечи, наблюдаемой в сырах с родительским штаммом лактококков.
В одной из первых серий выработок Чеддера в асептической ванне, в опытных сырах, выработанных только с применением лактококковой закваски, в 7 из 15 сыров был обнаружен прогорклый вкус, который в более легкой форме ощущался только в 3-х контрольных сырах, выработанных из этого же молока в открытой ванне [398]. Порок был вызван размножением в сырах Clostridium tyrobutyricum, споры которого были в исходном молоке. В контрольных сырах рост маслянокислых бактерий был подавлен или в большой степени ингибирован штаммами Lbc. plantarum, обладающими специфической антагонистической активностью к маслянокислым бактериям. В опытных сырах лактобациллы не были обнаружены, следовательно, в контрольные сыры они попали во время выработки из внешней среды.
Исследования природы антагонизма показали, что выделенные из контрольных сыров штаммы Lbc. plantarum образуют в средах перекись водорода в концентрациях, ингибирующих или подавляющих развитие споровых анаэробных бактерий. В зависимости от штамма, мезофильные лактобациллы образовывали в период максимума от 0,06 до 2,06 мМ Н202 (2-70 мкг/мл). ЛД50 для различных видов маслянокислых бактерий равняется 8-11 мкг/мл, для молочного и диацетильного лактококков - 17-18 мкг/мл. Следовательно, можно подобрать штаммы мезофильных лактобацилл, которые будут продуцировать столько перекиси водорода, сколько нужно для подавления роста маслянокислых бактерий, но недостаточно для ингибирования роста лактококков. Кроме этого, мезофильные лактобациллы образуют Н202 в течение первых 72 ч с достижением максимума к концу этого периода. Таким образом, во время выработки сыра максимальное количество Н202 лактобациллы образуют в 3-суточных сырах, когда содержание лактококков уже близко к максимальному. Наиболее часто и наиболее сильные штаммы-антагонисты маслянокислых бактерий среди мезофильных гомоферментативных лактобацилл встречались у Lbc. plantarum с низкой скоростью кислотообразования в молоке (82%), в то время как среди штаммов Lbc. casei только 39% обладали слабой антагонистической активностью к маслянокислым бактериям. Штаммы Lbc. casei образуют также пироглутаминовую кислоту, обладающую широким спектром антибиотической активности.
У Lbc. plantarum обнаружены штаммы с сильной специфической антагонистической активностью типа бактериоцинов к энтеробактериям. Влияние включения в закваски штаммов лактобацилл на развитие БГКП в Костромском сыре показано в табл. 3.20.
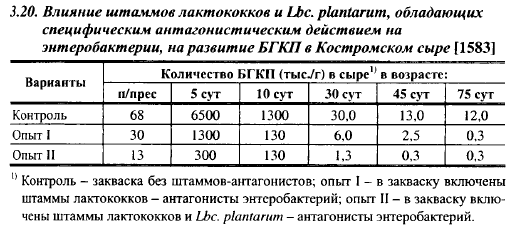
В нашей стране закваски, содержащие, наряду с лактококками, штаммы Lbc. plantarum со специфической антагонистической активностью к маслянокислым и энтеробактериям, широко применяются в промышленности с начала 70-х годов, и они получили высокую оценку производственников. В настоящее время биологические методы борьбы с вредной для сыроделия микрофлорой разрабатываются и за рубежом. Специфический антагонизм мезофильных лактобацилл по отношению к технически вредной и патогенной микрофлоре выпал из поля зрения авторов экспериментов по выработке сыров в асептических условиях, так как, за исключением указанной выше серии «асептических» сыров, в которой исходное молоко оказалось случайно загрязненным спорами С. tyrobutyricum, опыты по взаимодействию различных групп микроорганизмов в сырах, вырабатываемых в асептической ванне, не ставились.
Противоречие в оценке роли мезофильных лактобацилл в производстве мелких сыров авторами опытов по выработке сыров в асептической ванне с полностью контролируемым составом микрофлоры и научно-обоснованной практикой их применения в промышленности могут быть вызваны различиями в методике применения лактобацилл для выработки сыра. При оценке роли мезофильных лактобацилл в сыроделии их вносили в пастеризованное молоко, подготовленное для выработки сыра в количествах, намного превышающих количество лактобацилл в пастеризованном молоке в реальных производственных условиях. Внесение в молоко для выработки сыра достаточно большого количества лактобацилл стимулирует кислотообразующую активность лактококков, что приводит к резкому увеличению скорости нарастания кислотности сыворотки и сырной массы во время выработки сыра, изменению состава и структуры сырной массы, в частности, к снижению в ней содержания Са и Р и образованию кислого и горького вкуса и крошливой консистенции. Это наиболее распространенные пороки сыров, вина в которых возлагается на лактобациллы. В опытах Tittsler et al. добавление в закваски Lbc. easel повысило кислотность сыра, сделало его консистенцию более мягкой, ускорило созревание, но привело к появлению в сырах кислого и горького вкуса, крошливой консистенции. Именно из-за этого не получили признания комбинированные закваски (лактококки + лактобациллы), предложенные Руновым (1947).
Введение в состав закваски для производства мелких сыров штаммов Lbc. plantarum с низкой скоростью кислотообразования и роста в молоке в количестве, обеспечивающем внесение в смесь для выработки сыра 50-100 КОЕ/мл (0,5-1,0% от количества лактококков), не повышает скорость нарастания кислотности во время выработки сыра сверх допустимого уровня и не вызывает появления в сыре кислого, горького вкуса, крошливой консистенции. Не случайно Tittsler сумел получить положительные результаты при использовании в заквасках лактобацилл только со штаммами Lbc. plantarum. При целесообразности использования для выработки сыров штаммов лактобацилл с высокой скоростью кислотообразования, их нужно вносить в смесь в количестве нескольких клеток/мл с таким расчетом, чтобы они не оказали воздействие на процессы в сырной ванне. В настоящее время для ускорения созревания и улучшения качества сыров с помощью лактобацилл их клетки вносят в молоко для выработки сыра в состоянии шока (теплового или вызываемого неоднократным замораживанием и размораживанием), в результате которого они не размножаются во время выработки сыра и тем самым не изменяют скорость молочнокислого процесса. Перспективные с точки зрения качества сыра штаммы можно вносить не в молоко для выработки сыра, а в зерно после удаления сыворотки, когда их возможности изменить скорость кислотобразования в сырной массе будут ограничены.
Непосредственное влияние лактобацилл на органолептические показатели сыров как и любых других микроорганизмов зависит от их количества в сыре и характера метаболизма видов и штаммов. Голландские ученые считают, что если численность лактобацилл в сырах 4-6-недельного возраста не превышает 2107/г, то они не вызывают пороков в сырах независимо от видовой и штаммовой принадлежности. Это понятно, так как в этом случае биомасса лактобацилл составляет только около 1 % от биомассы лактококков, и поэтому влияние их на органолептические показатели не может быть заметным, особенно в связи с одинаковыми продуктами энергетического обмена лактококков и го- моферментативных лактобацилл. При использовании в составе заквасок штаммов мезофильных лактобацилл с достаточно высокой кислотообразующей активностью или при низкой активности лактококков, например, при загрязнении заквасок бактериофагами, количество лактобацилл в сыре достигает больше 109/г, что составляет уже около 50% от количества лактококков. В этом случае влияние лактобацилл на органолептические показатели сыров будет заметным и зависимым от индивидуальных свойств видов и штаммов. Так, при содержании в сырах (1-8)108/г гетероферментативных лактобацилл в мелких сырах часто наблюдается самокол. Большие количества мезофильных лактобацилл в сыре в зависимости от вида и штамма могут быть причиной появления в сырах нечистого, броженого, дрожжевого, горького, металлического, пряного вкуса и привкусов, самокола и щелевидного рисунка, пороков цвета, но могут и улучшать качество сыра. Это не является специфическим свойством лактобацилл: многие штаммы лактококков также вызывают пороки в сырах (горький вкус, фруктовый, солодовый привкусы и даже вспучивание). В связи с этим в закваски для приготовления сыра включают только специально отобранные штаммы лактококков. То же самое нужно делать и в отношении лактобацилл.
Высокое содержание лактобацилл в сырах приводит к трансформации L(+)-молочной кислоты, образуемой лактококками, в рацемическую смесь с образованием пентагидрата лактата Са, что вызывает пеструю окраску сырной массы (порок «мраморное тесто») и, возможно, мучнистую консистенцию.
Большинство авторов считает, что чаще всего пороки в мелких сырах вызывает гетероферментативная мезофильная лактобацилла вида Lbc. brevis; способность Lbc. plantarum вызывать пороки в сыре зависит от штамма, но обычно сыры с этим видом имеют более высокое качество и реже встречаемую и менее выраженную горечь.
Солеустойчивые штаммы Lbc. plantarum, попадающие в сыр из рассола и с сычужным порошком, могут вызывать в мелких сырах в старом возрасте самокол (они образуют СОг из аминокислот), фенольный и гнилостные запахи, мучнистую консистенцию. Для предотвращения появления пороков, вызываемых солеустойчивыми лактобациллами, их содержание в рассоле не должно превышать 1000 КОЕ/мл [884]. Некоторые штаммы Lbc. plantarum при достаточно высоком содержании в сыре вызывают появление пряного вкуса. Описан порок сыра Моцарелла «мягкое тесто», выражающийся в потере сыром плавимости и невозможности разрезать его на ломтики. Порок вызвала Lbc. casei subsp. casei. Из изложенного выше можно сделать следующий вывод: мезофильные гомоферментативные лактобациллы в зависимости от индивидуальных свойств штаммов и видов могут и ускорять созревание и улучшать качество мелких сыров, но могут и вызывать в них пороки. В состав заквасок для производства этих сыров нужно включать специально отобранные, проверенные штаммы лактобацилл, обладающие ценными для сыров свойствами.
Роль в крупных сырах. Размножение мезофильных гомофермента- тивных лактобацилл в Советском сыре показано в табл. 3.21.
В молоко для опытных сыров вносили около 50 КОЕ/мл мезофильных лактобацилл, в сырах после прессования их насчитывалось 1,7 млн. КОЕ/г, следовательно, они выдержали II нагревание и начали активно размножаться во время прессования сыров. Количество жизнеспособных клеток мезофильных лактобацилл продолжало увеличиваться до конца выдержки сыра в теплой камере. В контрольных сырах размножение мезофильных лактобацилл началось после прессования и так же, как в опытных сырах, прекратилось к концу выдержки в теплой камере.
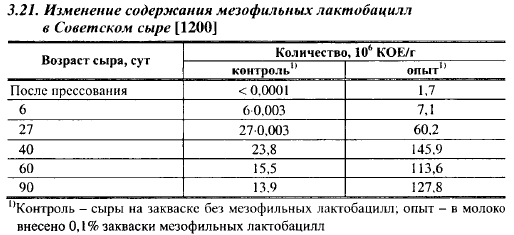
В зрелых сырах мезофильные лактобациллы составляли 62-98% от общего количества бактерий. В опытных сырах после прессования количество мезофильных лактобацилл примерно в 1200 раз превышало их количество в контрольных сырах, в 40-суточном возрасте (период максимума) - только в 6,3 раза. Изменение в соотношении, очевидно, обусловлено тем, что в сырах имеется ограниченное количество источников энергии, доступных для мезофильных лактобацилл, которое в опытных сырах было быстрее исчерпано этими бактериями. То, что максимальное содержание мезофильных лактобацилл в опытных сырах было выше, чем в контрольных, можно объяснить тем, что в опытных сырах они начали раньше размножаться, когда в сырной массе было достаточно лактозы.
Таким образом, главной функцией мезофильных лактобацилл в крупных сырах является сбраживание источников энергии, по каким-либо причинам несброженных микрофлорой традиционных заквасок, в состав которых входят лактококки, термофильный стрептококк и термофильные лактобациллы. Лактококки во время выработки крупных сыров погибают в процессе II нагревания, термофильный стрептококк перестает размножаться после прессования, термофильные лактобациллы - во время по- солки сыра. От того, каким бактериям достанутся несброженные микрофлорой закваски источники энергии, зависит качество сыра. В опытах, результаты которых изложены выше, в молоко, из которого вырабатывали опытные сыры, вносили специально отобранные штаммы мезофильных лактобацилл, в частности, не ингибирующие рост пропионовокислых бактерий. Опытные сыры в этих опытах имели более выраженный вкус, хорошую консистенцию и развитый рисунок: их оценка была на 3,4-4,6 балла выше оценки контрольных сыров, в которых мезофильные лактобациллы были представлены «дикими» штаммами.
В Эмментальском блочном сыре, вырабатываемом с применением традиционных заквасок, дикие штаммы мезофильных лактобацилл начали активно размножаться в сыре во время посолки, и их количество увеличивалось на всем протяжении созревания. Качество сыров в этих опытах снижалось при увеличении доли мезофильных лактобацилл в микрофлоре сыра. Среди «диких» штаммов мезофильных лактобацилл есть антагонисты пропионовокислых бактерий. В то же время применение в составе заквасок специально отобранных штаммов Lbc. casei ускорило созревание и улучшило вкус сыра Эмменталь.
Постоянное присутствие мезофильных лактобацилл в крупных и мелких сырах, влияние их видовых и штаммовых особенностей на качество продукта диктуют необходимость замены «диких» штаммов мезофильных лактобацилл культурными, специально подобранными для сыроделия. Закваски для крупных сыров, в состав которых входят мезофильные лактобациллы, вырабатываются в России с начала 80-х годов. Они присутствуют в сывороточных заквасках, применяемых в производстве итальянских сыров, а в настоящее время ставится задача их включения в промышленные бактериальные препараты для производства крупных сыров.
Термофильные лактобациллы
В сыроделии для производства твердых сыров с высокими температурами II нагревания применяют Lbc. helveticus (швейцарская палочка) и Lbc. delbrueckii с подвидами: bulgaricus (болгарская палочка) и lactis (молочная палочка). Для производства кисломолочных сыров применяют Lbc. acidophilus (ацидофильная палочка). В крупных сырах часто обнаруживают гетероферментативный вид термофильных лактобацилл - Lbc. fermentum, обычно обнаруживаемый и в естественных (сывороточных) заквасках для производства крупных сыров, но в состав промышленных заквасок этот вид, за небольшим исключением, не включают. Дифференциальные характеристики гомоферментативных термофильных лактобацилл, применяемых в сыроделии, приведены в табл. 3.16.
Термофильные лактобациллы выдерживают пастеризацию молока, принятую в сыроделии; кислотоустойчивы: предельная кислотность в молоке (% молочной кислоты) у болгарской и молочной палочек равна 1,5-1,7, у швейцарской - 2,7 (табл. 3.18). Средняя кислотность молока после инкубации в течение 24 ч при 37° С (г мол. кислоты/100 мл) была для штаммов Lbc. helveticus 1,25, болгарской и молочной палочек 0,78; pH, соответственно, равнялся 3,81 и 4,0. В опытах Тер-Казарьяна с соавт. прирост кислотности в 27 культурах Lbc. helveticus после инкубации при 45° С и 5 %-ной посевной дозе в течение 6 ч равнялся 33° Т. При средней кислотности суточных культур 144° Т колебания составили от 86 до 195° Т. «Медленные» штаммы Lbc. helveticus свертывали молоко при 5% посевной дозе и оптимальной температуре за 7 сут, «быстрые» - за 16 ч. Они обладали низкой казеолитической активностью. Болгарская палочка при инокуляции 3% и оптимальной температуре свертывает молоко за 3-4 ч. В совместных культурах термофильные лактобациллы и термофильный стрептококк, а также лактококки стимулируют рост друг друга.
Температура II нагревания в производстве крупных сыров равна 50-57° С, т. е. близка к максимальной границе для роста термофильных лактобацилл или немного выше ее (табл. 3.18). После II нагревания во время выработки крупных сыров они быстро восстанавливают активность и начинают размножаться во время прессования сыров. Термофильные лактобациллы прекращают размножаться во время посолки, когда температура сыра снизится до 18-20° С, и вознобновляют на какое-то время размножение после помещения сыра в теплую камеру.
Термофильный стрептококк стимулирует рост болгарской палочки путем образования небольших количеств муравьиной кислоты и С02; лактобациллы, в свою очередь, обеспечивают лактококки низкомолекулярными продуктами протеолиза. Взаимостимулирующее действие отмечено и между термофильным стрептококком и Lbc. helveticus (Accolas et al., 1971). Лактококки в смешанных культурах в соотношении 4:1 стимулируют рост в молоке термофильной микрофлоры заквасок для производства крупных сыров.
Характер взаимодействия между термофильными лактобациллами и маслянокислыми бактериями зависит от индивидуальных свойств штаммов. В средах с контролируемым pH (не ниже 5,2) 75% штаммов Lbc. helveticus, 62,5% штаммов болгарской и 60% штаммов молочной палочек стимулировали рост маслянокислых бактерий. Komonen нашел луночным методом штаммы Lbc. helveticus и болгарской палочки, которые ингибировали рост маслянокислых бактерий в твердой среде. Антагонизм по отношению к маслянокислым бактериям у болгарской палочки был сильнее, но она угнетает рост и пропионовокислых бактерий.
Термофильные лактобациллы выполняют в крупных сырах две функции: сбраживают углеводы и вносят большой вклад в формирование специфических для этого класса сыров органолептических показателей. Они не могут быть заменены термофильным стрептококком из-за особенностей сбраживания последним лактозы, что обусловливает накопление в сырах галактозы. Термофильные стрептококки чаще всего не сбраживают галактозу или сбраживают ее очень медленно.
Галактоза при использовании в составе закваски Гал штаммов термофильного стрептококка может быть сброжена другими микроорганизмами закваски: лактококками и лактобациллами или посторонней микрофлорой. Лактококки сбраживают галактозу в составе лактозы го- моферментативным путем, а свободную галактозу, за исключением отдельных штаммов, трансформируют в лактат, формиат, ацетат, этиловый спирт и, возможно, в небольшие количества СОг, причем доля лактатов составляет от 34 до 74% от количества сброженной галактозы. Однако, лактококки почти полностью погибают во время II нагревания. Из термофильных лактобацилл галактозу сбраживает Lbc. helveticus с образованием D(-)- и L(+)-молочной кислоты; Lbc. del- brueckii subsp. lactis и Lbc. delbrueckii subsp. bulgaricus галактозу не сбраживают (из глюкозы образуют D-молочную кислоту). Сбраживание углеводов в Эмментальском сыре показано на рис. 3.9, образование пропионатов и ацетатов - на рис. 3.10.
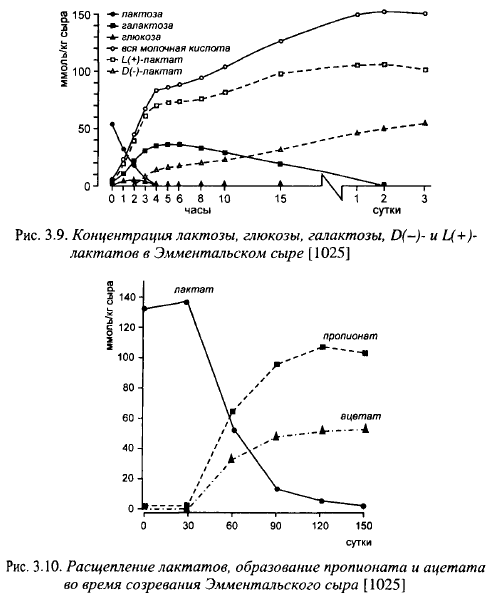
Таким образом при участии термофильного стрептококка в сбраживании лактозы во время выработки сыров с высокими температурами II нагревания в состав заквасок должна входить Lbc. helveticus, сбраживающая галактозу, остающуюся после сбраживания термостойким стрептококком лактозы. Если в состав закваски включены друг ие виды термофильных лактобацилл, не ферментирущие галактозу, галактоза в сыре не будет сброжена заквасочной микрофлорой. Чем больше останется несброженной термофильными лактобациллами закваски галактозы, тем больше будет возможностей для размножения в крупных сырах мезофильных лактобацилл, в том числе гетероферментативных, которые ухудшают вкус, консистенцию и рисунок сыров. Кроме этого, Lbc. helveticus образует D(-)- и L(+)-молочную кислоту, болгарская и молочная палочки только D(-)-молочную кислоту, а пропионовокислые бактерии более охотно используют L(+)-лактаты.
Lbc. helveticus не размножается при посеве в среду, содержащую 2% и больше NaCl, но если ее добавить в среду после роста этого вида в течение 20 ч при 42° Сив присутствии мела, то предельная концентрация соли повышается до 4%.
В свою очередь, термофильные лактобациллы не могут заменить лактококки и термофильный стрептококк в заквасках для производства крупных сыров, что изменило бы соотношение между L(+)- и D(-)-лак- татами в сыре, и, главное, большое количество термофильных лактобацилл в молоке во время выработки сыра резко увеличивает скорость кислотообразования, изменяет состав молочного сгустка и ухудшает качество сыра. В опытах Biede et al. добавление в молоко для выработки Швейцарского сыра в дополнение к обычной закваске даже 0,04% закваски болгарской палочки вызвало такие пороки в сыре, как горький и кислый вкус, правда, в легкой форме. В малых же дозах термофильные лактобациллы не успевают сбродить все сахара в сыре даже в присутствии лактококков и термостойкого стрептококка. Таким образом, в сбраживании сахаров в производстве крупных сыров принимают участие лактококки, термофильный стрептококк, Lbc. helveticus и, как показано в предыдущем разделе, мезофильные лактобациллы. Такое положение обусловлено сочетанием в технологии этих сыров высоких температур II нагревания и низких температур созревания.
Из «диких» термофильных лактобацилл гетероферментативный вид Lbc. fermentum сбраживает лактозу и галактозу с образованием С02, который, по мнению итальянских ученых, причастен к вспучиванию крупных сыров. Это может произойти, если достаточно большое количество углеводов не будет сброжено микрофлорой заквасок. В то же время среди представителей этого вида обнаружены штаммы, обладающие специфическим антагонистическим действием по отношению к маслянокислым бактериям. Разработан и вырабатывается в промышленных масштабах бактериальный концентрат для производства крупных сыров, в состав которого входят штаммы термофильных лактобацилл, обладающие специфичным антагонизмом к маслянокислым бактериям.
Важную роль в формировании специфического сырного вкуса в крупных сырах играет высокая протеолитическая активность термофильных лактобацилл, особенно способность высвобождать пролин и другие аминокислоты. По протеолитической активности термофильные лактобациллы намного превосходят термофильный стрептококк и пропионовокислые бактерии. Недостаточный протеолиз приводит к появлению в сырах таких дефектов, как слабовыраженный сырный вкус и аромат.
Высокая внутриклеточная протеолитическая активность Lbc. helveticus, проявляющаяся в образовании низкомолекулярных продуктов расщепления казеина, в частности свободных аминокислот, в настоящее время используется для ускорения созревания и предупреждения горечи в мелких сырах. Для того чтобы использование этого вида не изменило характер кислотообразования во время выработки сыра, клетки Lbc. helveticus вносят в молоко в состоянии шока, достигаемого высушиванием, сублетальным нагреванием, замораживанием, или в виде бесклеточных экстрактов или протопластов, получаемых под действием лизоцима. Эти факторы блокируют кислотообразование, но не активность внутриклеточных протеолитических энзимов, которая проявляется во время созревания сыра. Положительные результаты получены при использовании Lbc. helveticus и Lbc. lactis в производстве блочного сыра с низкими температурами II нагревания, что можно объяснить более медленным остыванием сырной массы в этом сыре.
Испытано влияние внесения в лактококковую закваску Lbc. helveticus WSU19 на химический состав и сенсорные показатели сыра Чеддер. Качество опытного сыра через 3 и 6 месяцев созревания было выше, чем контрольного. В опытных сырах интенсифицировался протеолиз, обнаружена азотистая фракция с молекулярной массой 11 кДа, которая не обнаружена в контрольных сырах.
