Данные о термоустойчивости пропионовокислых бактерий противоречивы. По результатам опытов Алексеевой с соавт., во время пастеризации молока при 71° С в течение 15 с погибает 99,8-99,97% клеток пропионовокислых бактерий и в пастеризованном молоке, поступающем в сырную ванну, их количество составляло от менее 1 до 250 клеток/мл. В опытах Белоусовой 10 штаммов пропионовокислых бактерий выдерживали нагревание при 55, 60 и 65° С в течение 60,15 и 5 мин соответственно. При этом температуры 71-72° С явились критическими: нагревание при 72° С в течение 10 с они выдерживают, а в течение 20 с уже не выдерживают. Молодые клетки более чувствительны к нагреванию, термоустойчивость в молоке выше, чем в бульоне. Двухчасовая выдержка молока при 55° С - температуре II нагревания при выработке крупных сыров - сильно уменьшает содержание пропионовокислых бактерий.
Для сыроделия особый интерес представляют минимальные температуры для роста пропионовокислых бактерий. Изучение 33 коллекционных и 16 неидентифицированных свежевыделенных штаммов пропионовокислых бактерий показало, что при 2,8° С только 3 свежевыделенные культуры дали сомнительный рост, при 7,2° С могли расти 12 свежевыделенных и 16 коллекционных штаммов, при 12,8° С росли все исследованные культуры за исключением двух коллекционных. Большинство штаммов пропионовокислых бактерий, выделенных из Советского сыра, не росли при 8-10° С, но один штамм, отнесенный к Р. jensenii, в 100 мл лактатной среды за 5 сут при 8-10° С образовал 14 мг С02.
Рост пропионовокислых бактерий в сырах во время их созревания в прохладных камерах вызывает самокол, что, очевидно, связано с недостаточной пластичностью сырной массы при низких температурах. В связи с этим в производстве крупных сыров следует использовать только штаммы пропионовокислых бактерий, неспособные ферментировать лактаты в прохладных камерах для созревания. Следует отметить, что Р. shermanii могут образовывать СО2 при 7° С не только из лактатов, но и из свободных аминокислот.
Разработан способ экспериментальной селекции чувствительных к холоду мутантов пропионовокислых бактерий, заключающийся в обработке родительской культуры Nметил-N'-нитро-нитрозогуанидином (150 мг/мл в течение 30 мин при 32° С), достаточной для уничтожения 99% клеток, с последующей инкубацией в среде с 2000 мг/мл пенициллина G при 14° С в течение 48 ч для отбора мутантов, неспособных расти при температуре 14° С и ниже. Получено девять мутантов, которые росли при 37° С с такой же скоростью, с какой росли родительские штаммы, но намного медленнее родительских при 14° С.
В молоке даже при оптимальных температурах пропионовокислые бактерии размножаются медленно, некоторые виды и штаммы вообще не свертывают молоко, другие свертывают его в течение 2-9 сут. Скорость образования пропионовой и уксусной кислот повышалась в 2,25-1,5 раза при внесении в молоко дрожжевого экстракта и в 1,6-4,6 раза в присутствии лактата натрия. Пептонизированное молоко, гидролизат казеина и печеночный экстракт оказали менее сильное стимулирующее действие на пропионовокислые бактерии.
На накопление биомассы пропионовокислых бактерий оказывает влияние внесение в среду бесклеточных фильтратов или культуральной жидкости молочнокислых бактерий, используемых в производстве сыров с высокими температурами II нагревания. Характер влияния зависит от вида и штамма молочнокислых бактерий. Так, фильтраты термофильного стрептококка и L. helveticus, но не молочного лактококка, стимулировали развитие Р. freudenreichii subsp. freudenreichii и в меньшей степени subsp. shermanii. Выдержка при 80° С в течение 20 мин существенно уменьшила стимулирующее действие фильтратов. Аминокислоты и пептиды, образуемые молочнокислыми бактериями, в частности L helveticus, за исключением треонина и цистина, использовались про- пионовокислыми бактериями. Сычужный порошок и плазмин в отсутствие молочнокислых бактерий не влияют на размножение; в присутствии молочнокислых бактерий, осуществляющих дальнейший гидролиз (до свободных аминокислот и низкомолекулярных пептидов) продуктов расщепления казеина сычужным порошком и плазмином, рост пропионовокислых бактерий стимулируется. Однако слишком высокое содержание свободных аминокислот в среде может ингибировать рост пропионовокислых бактерий. Добавление в сывороточную среду аспартата стимулировало рост Р. freudenreichii, но понизило долю пропионовой кислоты. образующейся при сбраживании лактата.
Выделенные из крупных сыров штаммы L rhamnosus и L. casei отрицательно влияли на рост пропионовокислых бактерий в сырах с высокими и средними температурами II нагревания. Это же отмечали Климовский с соавт. Другие авторы отмечают стимулирование роста пропионовокислых бактерий штаммами L casei и их положительное влияние на образование глазков и вкус сыра. Различие в результатах этих опытов вызвано условиями их проведения, в частности способами нейтрализации кислоты, образуемой лактобациллами, хотя нельзя исключить и образование некоторыми штаммами лактобацилл специфических антибиотических веществ в дополнение к органическим кислотам.
В опытах Lee et al. при совместном развитии в среде с минимальным содержанием глюкозы в анаэробных условиях между L. plantarum и Р. shermanii наблюдался комменсализм, несмотря на снижение pH, которое однако лимитировалось содержанием глюкозы.
Из 12 штаммов термофильных лактобацилл рост пропионовокислых бактерий ингибировали 2 штамма L lactis, 2 штамма L. helveticus и I штамм L acidophilus. L. helveticus и L. acidophilus ингибировали II из 17 штаммов пропионовокислых бактерий, lactis ингибировал развитие небольшого количества штаммов. Ингибирующие факторы были термостабильные и термолабильные.
С другой стороны, использование культуральной среды некоторых штаммов мезофильных и термофильных лактобацилл в производстве концентрата пропионовокислых бактерий позволило не только увеличить выход их биомассы, но и повысило скорость развития пропионовокислых бактерий в крупных сырах. По-видимому, влияние молочнокислых бактерий на рост пропионовокислых многопланово, включает образование стимулирующих и ингибирующих факторов.
Молочнокислые бактерии, в зависимости от рода, образуют разные изомеры молочной кислоты, а пропионовокислые бактерии сбраживают изомеры молочной кислоты с неодинаковой скоростью. В среде со смесью изомеров молочной кислоты пропионовокислые бактерии прежде всего используют L(+)-лактат, что должно учитываться при подборе микрофлоры заквасок для производства крупных сыров. В культуральной среде L helveticus концентрация L(+)-лактата выше, чем D(-)-лактата.
Противоречивы данные об устойчивости пропионовокислых бактерий к поваренной соли, что, по-видимому, обусловлено ее зависимостью от штамма и других показателей среды, в частности от pH [401]. Antilla (1955) сообщал, что ингибирование роста пропионовокислых бактерий начинается при содержании в среде 3% NaCl. Рост быстрорастущих штаммов пропионовокислых бактерий в лактатной среде при pH 7,0 ингибировали 6% соли, при pH 5,2 - уже 3%, медленно растущие штаммы, наоборот, были более солеустойчивы при pH 5,2 (Rollman & Sjostrom, 1946). Наиболее устойчив к соли Р. shermanii (предельная концентрация 6,7%, Ав = 0,955), что, очевидно, и является причиной преобладания этого подвида в Советском сыре. В опытах Алексеевой и Анищенко солеустойчивость пропионовокислых бактерий, выделенных из Советского сыра, изучалась в лактатной среде с pH 7,0 при 22° С. Из 23 исследованных штаммов наибольшую устойчивость к соли показали два штамма Р. shermanii и Р. technicum: они росли при 5,2% соли. В среде с 6,5% соли жизнеспособные клетки после 27 сут инкубации сохранились только у 6 штаммов. Скорость накопления биомассы в среде с 4,5% NaCl была в 10 раз ниже, чем в присутствии 1,4% NaCl. Чувствительность к соли зависит от штамма.
Содержание соли в зрелых отечественных сырах с высокой температурой II нагревания (Советском, Швейцарском) обычно находится на уровне 1,5-2,0%, что составляет 3,8-5,0% в водной фазе; в сырах этого класса, вырабатываемых в Швейцарии, оно в среднем равно (%): в Эм- ментальском - 0,65 (1,8 в водной фазе), в Грюйере - 1,45 (3,9), в Аппен- целлере - 1,6 (3,9). Таким образом, уровень посолки зарубежных сыров с высокими температурами II нагревания ниже, чем отечественных. В то же время интенсивность развития пропионовокислых бактерий в сырах зависит от уровня их посолки (табл. 3.23). Содержание пропионовокислых бактерий неодинаково в различных слоях головки сыра: в наружном слое Советского сыра, в котором концентрация соли в водной фазе составила 4,4%, в 90-суточном возрасте содержание пропионовокислых бактерий равнялось 32-106, в центральном слое с концентрацией соли в водной фазе 2,9% - 69-106 КОЕ/г. Для сравнения укажем, что в сырах этого класса, вырабатываемых в Швейцарии, количество пропионовокислых бактерий в период максимума равняется (1-10)-108 КОЕ/г , а содержание кислот в них показано в табл. 12.5. Сыр Советский отличается от Швейцарского (Эмментальский сыр часто называют просто «Швейцарским») не только значительно более низким содержанием соли, но и меньшей массой головки (11-18 по сравнению с 60-130 кг), что ускоряет проникновение соли во внутренние слои головки и вызывает ингибирование развития пропионовокислых бактерий на более ранних стадиях созревания сыра. Однако и в Советском сыре соль довольно медленно проникает во внутренние слои. Так, в 90-суточных сырах концентрация соли в водной фазе в центральных слоях была ниже, чем в слоях, прилегающих к поверхности головки, в зависимости от продолжительности посолки в 1,3-1,5 раза, а содержание пропионовокислых бактерий, наоборот, выше в 2,1-2,6 раза. Таким образом, пропионовокислые бактерии практически на всем протяжении созревания имеют возможность размножаться в центральных слоях Советского сыра даже с учетом высокой степени его посолки, однако неодинаковая скорость их развития в пределах головки влечет за собой и различия в органолептических показателях: степень выраженности сырного вкуса усиливается, а твердость сырного теста снижается по направлению от периферии к центру.
Большая интенсивность пропионовокислого брожения в Эмменталь- ском сыре обусловливает его более выраженный вкус и аромат, но повышает чувствительность к маслянокислому брожению, по сравнению с Советским сыром. Из крупных сыров, вырабатываемых в Швейцарии, Эмментальский обладает более выраженным специфическим для этого класса сыров вкусом и более развитым рисунком, чем Грюйер и Аппенцеллер.
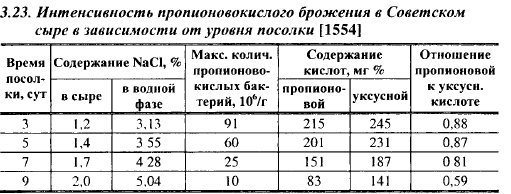
Ингибирующее действие соли на пропионовокислые бактерии снижается при повышении pH и увеличении содержания влаги в сыре. В молоке с 4% NaCl пропионовокислые бактерии растут, но начинают образовывать С02 значительно позднее, чем в молоке без добавления соли.
Оптимальный pH для пропионовокислых бактерий равен 6,8-7,2, что намного выше pH крупных сыров (5,2-5,6), однако и в крупных сырах pH выше минимального для роста пропионовокислых бактерий. Влияние pH на развитие в Советском сыре пропионовокислых бактерий показано в опытах, проведенных в Алтайском филиале ВНИИМС. В опыте pH сыров регулировали степенью разбавления водой сыворотки после отбора 30% ее количества. Результаты этого опыта приведены в табл. 3.24. Перед помещением в бродильную камеру pH сыров II и III вариантов были выше pH контрольного сыра, во время выработки которого сыворотку не разбавляли водой, на 0,06 и 0,13 ед. соответственно. Количество пропионовокислых бактерий за период после окончания посолки и до момента выхода из бродильной камеры возросло в контрольных сырах в 63,6 раза, в сырах II варианта - в 200 раз и III варианта - в 236 раз. Содержание этих бактерий в период максимума в сырах I, II и III вариантов находилось в отношении 1:2:3.
Недиссоциированная молочная кислота ингибирует рост пропионовокислых бактерий, степень этого ингибирования зависит от штамма и вида пропионовокислых бактерий. При исследовании Р. jensenii Р16 и P.freu- denreichii subsp. globosum P14, выделенных соответственно из вспученного и нормального сыра Грана, молочная кислота значительно сильнее ингибировала штамм Р14. Основная масса пропионовокислых бактерий погибает при пастеризации молока; уцелевших после пастеризации клеток недостаточно для обеспечения нужного уровня пропионовокислого брожения в сыре. Достаточным количеством пропионовокислых бактерий в Советском сыре при нормальном технологическом процессе будет наличие в нем после прессования (2-4)-103 КОЕ/г, что обеспечивается внесением в молоко после пастеризации 1-2 мл на 5000 л молока чистой культуры пропионовокислых бактерий или 1-2 тыс. КОЕ/мл. Следует, однако, отметить, что содержание пропионовокислых бактерий в Советском сыре в большей степени зависит не от их количества, вносимого в молоко, а от других факторов. В опытах Климовского с соавт. количество внесенных в молоко пропионовокислых бактерий для выработки сыров двух вариантов отличалось в 1800 раз, а максимальное их количество в сырах - только в 1,5 раза, хотя при использовании большей дозы максимум наступил намного раньше.
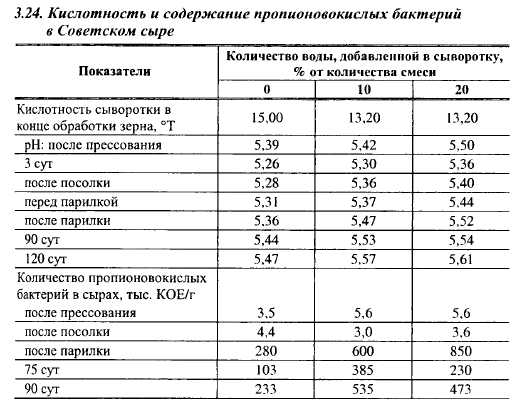
Приведенные выше данные о чувствительности пропионовокислых бактерий к содержанию в сырах соли и pH казалось бы свидетельствуют о том, что именно эти факторы должны играть главную роль в темпах развития пропионовокислых бактерий в Советском сыре. Однако в сырах должны быть и другие факторы, лимитирующие рост пропионовокислых бактерий. Если бы, например, концентрация соли в водной фазе сыра была единственным фактором, лимитирующим их рост, пропионовокислые бактерии прекращали бы размножаться в сырах в одно и то же время независимо от дозы их внесения, когда концентрация соли в центральных слоях превысит максимальную границу для их развития. В предыдущем опыте количество жизнеспособных клеток пропионовокислых бактерий в варианте с высокой посевной дозой перестало увеличиваться в сыре в конце его выдержки в теплой камере (через 40 сут), когда концентрация соли в центральных слоях не бывает высокой, а в варианте с низкой посевной дозой их биомасса увеличивалась до конца созревания (100 сут), когда соль более равномерно распределяется в пределах головки.
В 1-ый период созревания Советского сыра при температуре 10-12° С пропионовокислые бактерии практически не размножаются. Не размножаются они в этот период и в Эмментальском сыре, хотя температура его созревания на первом этапе (12-14° С) несколько выше, чем в производстве Советского сыра. Ацетаты и пропионаты в Эмментальском сыре не образуются до его помещения в теплую камеру (рис. 3.10).
Климовский с соавт. отсутствие размножения пропионовокислых бактерий на первом этапе созревания объясняют низкой температурой и высокой скоростью кислотообразования микрофлорой закваски в этот период. Повышение температуры созревания сыров в бродильной камере (парилке) до 22-24° С призвано прежде всего обеспечить условия для роста пропионовокислых бактерий. Этому не противоречит часто наблюдаемое размножение пропионовокислых бактерий во время созревания сыра при 10-12° С после его выхода из бродильной камеры, так как pH сыров после парилки существенно повышается (табл. 3.34), а чувствительность пропионовокислых бактерий к низким температурам уменьшается с повышением pH. Однако объяснение Климовского с соавт. причин отсутствия размножения пропионовокислых бактерий в первый период созревания в прохладной камере недостаточно.
По данным итальянских ученых, пропионовокислые бактерии, выделенные из вспученных в результате их жизнедеятельности крупных сыров Пармиджано Регано и Грана Падано, размножались в сыре Грана, хотя и очень медленно, при комбинации трех факторов: температуры ниже 22° С, содержания более 2% соли и pH ниже 5,5. Во время предварительного созревания сыра при 10-12° С одной из причин отсутствия их размножения могут быть сублетальные повреждения клеток, полученные во время обработки зерна при температурах II нагревания, для устранения которых клетке необходимо определенное время. Кроме того, в этот период в сырах достаточно интенсивно размножаются цитратферментирующие мезофильные лактобациллы, многие из которых образуют нестойкие антибиотические вещества, которые могут ингибировать рост пропионовокислых бактерий.
Некоторое увеличение продолжительности первого периода позволяет пропионовокислым бактериям полнее репарировать сублетальные повреждения клеток бактерий. К тому же, эта мера увеличивает биомассу мезофильных лактобацилл к моменту помещения сыров в теплую камеру, а следовательно, увеличивает количество энзимов в сырной массе, необходимых для созревания сыра. Увеличение количества этих энзимов в сырной массе повышает скорость созревания и сокращает его продолжительность в Советском сыре на 25%.
Кроме этого, в сырах в этот период немного повышается pH, что скорее всего обусловлено утилизацией цитратов лактобациллами. Таким образом, увеличение продолжительности первого периода созревания сыра в прохладной камере должно усилить влияние повышения температуры в бродильной камере на созревание сыра примерно в два раза.
В начальный период созревания в сыре недостаточно высоко содержание свободных аминокислот и низкомолекулярных продуктов протеолиза, стимулирующих рост пропионовокислых бактерий. Швейцарскими учеными было доказано, что внесение в молоко перед сычужным свертыванием микрококков оказывает сильнейшее стимулирующее действие на пропионовокислые бактерии: в опытном Эмментальском сыре в трехмесячном возрасте содержание пропионовокислых бактерий равнялось (373-679)- 106/г, в контрольном, вырабатываемом без применения микрококков, - (239-356)-106. О более интенсивном развитии пропионовокислых бактерий в опытных сырах свидетельствует и тот факт, что pH этих сыров повысился до нормального уровня на две недели раньше, чем в контрольных (в крупных сырах pH во время созревания сыра повышается за счет сбраживания лактатов и протеолиза).
Интенсивность развития пропионовокислых бактерий зависит от сезона года, что может быть связано с кормлением коров и изменениями состава и свойств молока на протяжении лактации. Так, повышение содержания ненасыщенных кислот в молочном жире в пастбищный период может ингибировать пропионовокислое брожение в крупных сырах.
В сыворотке, полученной при выработке сыров швейцарского типа, установлено наличие термолабильных, ингибирующих рост пропионовокислых бактерий веществ. Ингибирующее действие сыворотки нельзя было объяснить влиянием pH. Вполне возможно, что микрофлора закваски в период активного роста образует специфические по отношению к пропионовокислым бактериям антибиотические вещества, которые разрушаются в процессе дальнейшего созревания сыров. Увеличение посевной дозы болгарской палочки при выработке Швейцарского сыра с О до 2500 мл на 9080 л молока снизило содержание пропионовокислых бактерий в сыре при выходе из бродильной камеры в 3 раза, количество пропионовой кислоты в 1,1 раза, несмотря на увеличение продолжительности созревания в бродильной камере с 21 до 28 сут, однако содержание пролина увеличилось в 1,91 раза, повысилась и степень выраженности сырного вкуса. Дальнейшее увеличение посевной дозы болгарской палочки уменьшило концентрацию пролина и выраженность сырного вкуса и вызвало пороки сыра (горький и кислый вкус).
Таким образом, в сырах мог ут действовать стимулирующие и ингибирующие рост пропионовокислых бактерий факторы, отличные от pH и концентрации соли, совокупность которых может подавить рост пропионовокислых бактерий в первый период созревания при прохладной температуре. Однако отсутствие размножения пропионовокислых бактерий в этот период не обязательно: в опытах А. Гудкова с соавт. пропионовокислые бактерии, хотя и медленно, но размножались на первом этапе созревания в сырах одной из 9 выработок. Количество пропионовокислых бактерий в блочном Эмментальском сыре во время его созревания в первой прохладной камере систематически возрастало примерно на два порядка. Другими особенностями микробиологических и биохимических процессов в этом сыре, вырабатываемом на Пайденском молкомбинате в Эстонии, были размножение споровых аэробных палочек, интенсивный протеолиз в течение первых 10 сут, существенное повышение pH в период после прессования до помещения сыра в бродильную камеру (с 5,2 до 5,4). Авторы исследований этого сыра считают, что энергичный протеолиз создал благоприятные условия для размножения пропионовокислых бактерий на первом этапе созревания блочного Эмментальского сыра.
Протеолитическая активность пропионовокислых бактерий значительно меньше протеолитической активности микрофлоры закваски, тем не менее они образуют пептидазы, вызывающие освобождение пролина, который участвует в формировании характерного для крупных сыров сладковатого вкуса. Недостаточный протеолиз обусловливает формирование слабовыраженного сырного вкуса и излишне плотной консистенции, иногда - неравномерного рисунка, излишний протеолиз приводит к образованию перезрелого, слишком острого вкуса и крошливой консистенции. При образовании в крупных сырах большого количества низкомолекулярных продуктов протеолиза и слишком интенсивного развития пропионовокислых бактерий количество образуемого СО2 увеличивается за счет декарбоксилирования, что приводит к формированию излишне развитого рисунка, образованию трещин и снижению стойкости в хранении сыра.
Многие авторы считают, что излишнее развитие пропионовокислых бактерий может вызвать пороки в крупных сырах. Так, во вспученном сыре Сбринц с крошливой консистенцией, самоколом, повышенным pH были обнаружены пропионовокислые бактерии в количестве 108— 1 С)9 КОЕ/г. Кроме них, в дефектных сырах было 4,6Т08 КОЕ/г Е. faecalis и 9,8-108 КОЕ/г L. fermentum против 7,6-106 и 3-103 КОЕ/г в сырах без пороков. Авторы работы делают вывод о том, что энтерококки стимулировали рост пропионовокислых бактерий, которые вместе с L. fermentum вызвали порчу сыров.
Пропионовокислые бактерии обладают более высокой липолитической активностью, чем молочнокислые бактерии, применяемые в производстве крупных сыров [1030], однако среди них имеются виды и штаммы с высокой и низкой липолитической активностью (табл. 3.1).
По мнению швейцарских ученых, излишний протеолиз в сырах, в частности повышенная активность лейцинаминопептидазы, интенсифицирует развитие пропионовокислых бактерий и может привести к позднему вспучиванию сыра. Вспученные сыры Грюйер отличались от доброкачественных более интенсивным развитием пропионовокислых бактерий, более глубоким протеолизом, более высоким содержанием воды и пониженным содержанием соли. При этом остается неясным, сами ли пропионовокислые бактерии являются виновниками порчи сыра, или их излишне активное развитие коррелирует с развитием других бактерий и даже стимулирует их рост, в результате которого происходит порча сыра. Виновниками порчи могут быть, например, маслянокислые бактерии, условия для развития которых могут создавать пропионовокислые бактерии, снижая pH сыра. Маслянокислые бактерии, в отличие от пропионовокислых, кроме СО2 образуют водород, который плохо растворим в сырной массе и может вызвать вспучивание сыров на фоне высокого давления С02 даже при небольших концентрациях. Вспучивание сыров могут также вызвать гетероферментативные лактобациллы, цитратсбраживающие мезо- фильные лактобациллы, в частности L plantarum.
Для предотвращения этого Швейцарский исследовательский молочный институт отобрал из диких штаммов пропионовокислых бактерий штаммы с пониженной скоростью развития в сырах. В России подбором и производством бакпрепаратов и концентратов пропионовокислых бактерий для сыроделия занимаются лаборатория бактериальных препаратов и заквасок в Барнауле и, в меньшем объеме, Угличская экспериментальная биофабрика.
Развитие пропионовокислых бактерий в сырах с низкой температурой II нагревания до уровня более 107г вызовет в нем появление несвойственного вкуса, что расценивается как порок.
