Чистые культуры лактобактерий в молоке, инфицированном вирулентными для них фагами, вначале развиваются без видимых отклонений от нормы. Через 2-4 ч большинство бактериальных клеток лизируется, и нарастание кислотности приостанавливается. После определенной выдержки кислотность снова начинает расти (так называемый «вторичный рост») за счет фагорезистентных мутантов, которые практически всегда есть в популяции чувствительных к фагу штаммов. Свертывание молока лактококком при 30° С и 5 %-ном инокулировании в присутствии фага-гомолога происходит не через 5-7 ч, а через 20-36 ч, причем клетки в сгустке будут представлять главным образом не исходный штамм, а его спонтанные фагорезистентные мутанты.
При приготовлении многовидовых и многоштаммовых заквасок полная остановка нарастания кислотности молока может произойти при одновременном заражении закваски фагами, способными лизировать все входящие в ее состав виды и штаммы лактобактерий с достаточно высокой скоростью кислотообразования в молоке, что бывает редко. Чаще всего поражается один или несколько таких штаммов, что ведет к снижению кислотности закваски в конце ее культивирования в течение обычного промежутка времени. Такая закваска должна сниматься с производства даже при небольшом снижении ее кислотности, так как она содержит большое количество вирионов, среди которых могут появиться или уже появились мутанты, способные лизировать другие штаммы, входящие в состав закваски. Для того чтобы не остановить производство сыра, предприятие должно иметь бактериальные концентраты, пригодные для непосредственного внесения в смесь для выработки сыра или для приготовления производственной закваски ускоренными способами.
При загрязнении фагами, лизирующими газообразующую микрофлору, закваски перестают образовывать диацетил и С02 и становятся непригодными для выработки сыров с правильным рисунком. Это маловероятно для заквасок, в состав которых входят лейконостоки, поскольку фаги лейконостоков редко обнаруживаются на сыродельных предприятиях, а активное размножение лейконостоков во время выработки сыра начинается уже в сформованном сыре, когда фагам труднее вступить в контакт с клетками возможного хозяина.
Инфицирование фагами может произойти в сырной ванне. В этом случае фаг не окажет влияния на свертывание молока в производстве сычужных сыров, в котором микрофлора участия не принимает.
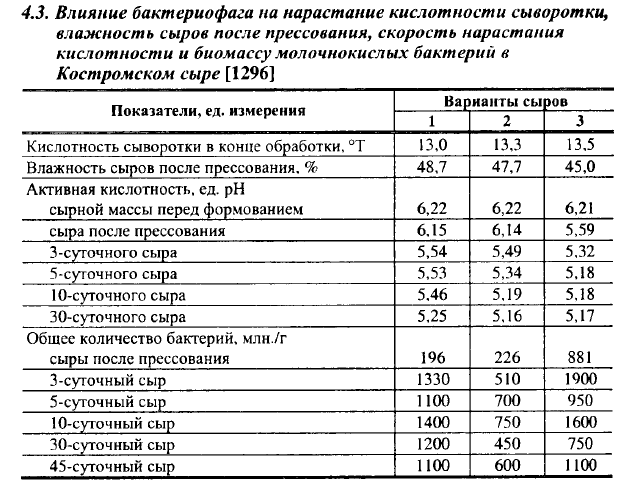
Для выявления действия фага на микробиологические процессы и качество сыров были проведены выработки Костромского сыра на закваске, в состав которой было включено по одному штамму каждого подвида лактококков. В первой серии в смесь для выработки сыра в каждом варианте вносили фаги только к одному штамму, различия в вариантах были небольшие. Во второй серии в смесь I варианта вносили фагигомологи к штаммам молочного и сливочного лактококков, II варианта - ко всем трем подвидам, в смесь для III варианта (контроль) фаги не вносили. Некоторые показатели опытных сыров приведены в табл. 4.3. Фаговая инфекция в опытных сырах (3-150 тыс. вирионов/мл) начала проявляться к концу обработки зерна, что выражалось в несколько меньшей кислотности сыворотки в опытных вариантах. Однако pH сырной массы перед формованием был во всех вариантах одинаковым. Резкое падение скорости кислотообразования в опытных сырах началось во время прессования сыров; pH опытных сыров был на 0,55 ед. выше, чем в контрольных. Существенные различия в pH опытных и контрольных сыров сохранялись вплоть до 10-суточного возраста. Лактоза в контрольных сырах была сброжена к 5-суточному возрасту, тогда как в опытных сырах в этом возрасте ее оставалось 0,115-0,150 г%. Остается неясным, почему pH в опытных сырах II варианта, в смесь для выработки которых добавляли фаги ко всем штаммам закваски, снижался несколько быстрее, чем в сырах I варианта, в смеси которого не было фагов к диацетильному лактококку. Возможно, в сырах II варианта больше осталось несброженной лактозы после прессования сыра, в результате чего при ее сбраживании в более поздние сроки вся образующаяся из этой лактозы кислота осталась в сыре, тогда как при сбраживании лактозы во время выработки сыра часть образующейся кислоты уходит с сывороткой.
По количеству бактерий в сырах после прессования и 3-суточного возраста контрольные сыры превосходили опытные, особенно сыры II варианта, но при последующем созревании высокое количество жизнеспособных клеток сохранялось в опытных сырах I варианта.
Содержание БГКП в опытных сырах после прессования было в 450 раз выше, чем в контрольных сырах. Контрольные сыры были оценены высшим сортом, с оценкой 91,4 балла, опытные - первым, с оценкой 88,3 балла, в т. ч. 36,6 за вкус и запах. В сырах II варианта рисунка не было или был рваный рисунок, образованный в результате продуцирования Н2 посторонней микрофлорой.
Опыт показывает, что массивное загрязнение фагами смеси для выработки сыров в сырной ванне может оказать мощное воздействие на микробиологические процессы и качество сыра, но это влияние начинает проявляться не в сырной ванне, а во время прессования, и оно может быть обнаружено по pH сыров после прессования. Вследствие этого необходимо обязательно включать определение pH сыров после прессования в схему контроля производственных процессов. Умеренное развитие фагов во время выработки не оказывает существенного влияния на технологический процесс и качество сыра.
Анализ большого количества сыров с низкими температурами II нагревания, выработанных с Угличскими заквасками, показал, что в них после прессования обычно содержится около 1,4-109 КОЕ/г молочнокислых бактерий, а выработанных в условиях фаговой инфекции (0,4-0,6) 109 КОЕ/г. Численность лактококков в пораженных фагами сырах достигает нормы в 5-10-суточном возрасте, а может ее не достигнуть до конца созревания. В последнем случае в сырах обнаруживали несброженную лактозу даже в середине периода созревания.
Интересны результаты опыта по выработке Чеддера в условиях фаговой инфекции. Сыр вырабатывали с молочным лактококком ML8 как одноштаммовой закваской, двумя дозами сычужного энзима и тремя дозами фагов-гомологов. Степень лизиса клеток в сыре зависела от дозы фагов-гомологов. Повышение дозы фагов увеличивало содержание свободных аминокислот и NH3 в сырах (за счет высвобождения внутриклеточных бактериальных протеиназ), но снижало скорость ферментации лактозы. Повышение дозы сычужного энзима увеличило содержание пептидов и горечь, последняя исчезала при повышении дозы фага. Таким образом, лизис части клеток микрофлоры закваски фагами может уменьшить горечь в сырах. Снижение горечи в сырах в результате лизиса части клеток лактококков в конце выработки сыра также отмечал Lowrie. Этот метод, конечно, нельзя рекомендовать для борьбы с горечью, так как снижение скорости сбраживания лактозы может привести к пищевому отравлению.
