Физиолого-биохимические свойства маслянокислых бактерий были подробно изучены в отделе микробиологии ВНИИМС (А. Гудков, Перфильев и др.).
Кислотность. Минимальные значения pH, при которых может происходить рост маслянокислых бактерий, при прочих оптимальных условиях в зависимости от штамма и вида равны 4,2-4,7. На средах с глюкозой маслянокислые бактерии наиболее энергично размножались при pH 6,6-6,0, в средах с лактатом Са в присутствии ацетатов при pH 5,5-6,0. Минимальный pH, при котором Сl. tyrobutyricum сбраживал лактаты, равняется 5,0, Cl. butyricum - 5,1-5,3; остальные виды маслянокислых бактерий - 5,4-5,5. Оптимальный pH для образования и прорастания спор маслянокислых бактерий выше, чем для роста.
Нетоксигенный штамм Cl. butyricum рос при pH 4,2, токсигенный - при pH не ниже 5,2.
На развитие Сl, tyrobutyricum оказывает влияние не только pH, но и содержание не- диссоциированной молочной кислоты: в модифицированной сыворотке из-под сыра Гауда рост этого вида при 30° С был задержан на 40 сут при pH не выше 4,8 и концентрации молочной кислоты 2%, или при pH 5,1-5,2 и содержании молочной кислоты 4%.
При низких pH меняется характер метаболизма маслянокислых бактерий: они перестают образовывать или резко снижают объем образуемых бутирата и водорода, ограничиваясь образованием ацетата и С02. Так, в опытах с выработкой сыра Чеддер в асептической ванне Сl. tyrobutyricum начал расти в некоторых опытных сырах, имеющих pH около 5,0, без следов газообразования. В процессе роста этого вида pH сыров повысился до 5,3 (маслянокислые бактерии при сбраживании лактатов образуют одну молекулу бутирата из двух молекул лактата), а в опытных сырах, в которых не было признаков роста маслянокислых бактерий он остался без изменения. При ухудшении условий для роста маслянокислых бактерий, прежде всего подавляется их газообразующая способность. При снижении активности воды минимальный pH для размножения маслянокислых бактерий повышается.
Активность воды и содержание соли. Минимальные значения Ав и максимальные концентрации соли, при которых происходит рост маслянокислых бактерий в средах с глюкозой, показаны в табл. 6.16. Они зависят от штамма и вида маслянокислых бактерий.

Чаще всего штаммы с высокой устойчивостью к Ав встречались среди Сl, tyrobutyricum. Это очень важно для сыроделия, поскольку, в отличие от большинства возбудителей пищевых отравлений, маслянокислые бактерии размножаются в сырах после того, как соль начала распространяться по всей массе головки сыра, что снижает Ав в глубинных слоях. Большинство штаммов Cl. tyrobutyricum прекращало рост в среде с 5%, штаммов другого вида - с 4% соли.
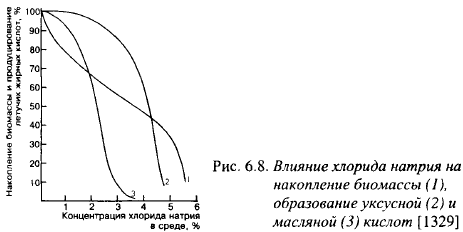
Продукты метаболизма Cl. tyrobutyricum в большой степени зависят от Ав среды (рис. 6.8): по мере увеличения концентрации соли снижается общее количество образуемых кислот, но количество масляной кислоты снижается намного быстрее, чем уксусной. Масляная кислота перестала образовываться в среде с 3,7% NaCl.
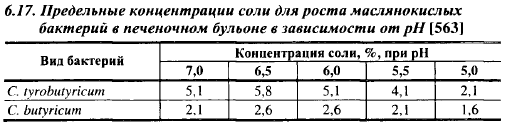
Чувствительность к соли зависит от pH, состава среды. В табл. 6.17 приведены предельные концентрации NaCl в печеночном бульоне для роста маслянокислых бактерий при 37° С в течение 4 сут. Наибольшей устойчивостью к соли они обладали при pH 6,5, при pH 5,1-5,5, характерном для твердых сыров, устойчивость к NaCl снижается. Добавление в печеночный бульон 1% лактата натрия несколько увеличило чувствительность к соли Cl. butyricum, а у Cl. tyrobutyricum она возросла только при pH 5,0. При выдержке в течение 14 и 60 сут максимальные концентрации соли для Cl. tyrobutyricum и Cl. butyricum при 24° С и pH равнялись соответственно 5,6 и 3,1%, при 37° С - 5,1 и 2,1%.
Температура. Влияние температуры на развитие маслянокислых бактерий в среде с глюкозой показано на рис 6.9. Оптимальные температуры для Сl, butyricum и Сl, pasteurianum лежат в интервале 26-34° С, Cl. tyrobutyricum - 26-37° С, максимальные лежат в пределах 46° С. Для сыроделия большее значение имеет нижняя граница для роста маслянокислых бактерий: для Сl, tyrobutyricum она равнялась около 8° С, для двух других видов - (11-12)° С.
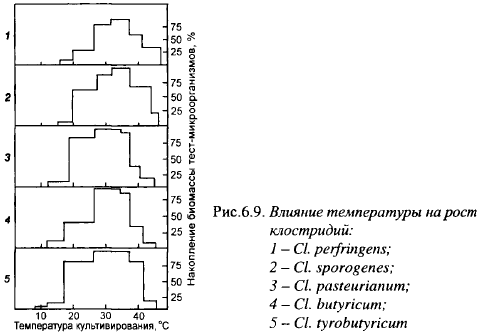
Температура влияет на выход и скорость накопления биомассы: при 20° С Сl, tyrobutyricum достигала стационарной фазы в зависимости от штамма за 150-190 ч, а при температурах 30-37° С - за 95 ч (70-120 ч). Выход биомассы и скорость размножения маслянокислых бактерий резко снижались при температурах ниже 16° С, а для Cl. tyrobutyricum также наблюдается подобный скачок при переходе температуры через 12° С. При сбраживании лактата кальция Сl, tyrobutyricum быстрее растет при 22-24° С (А. Гудков, неопубликованные материалы). Споры Сl, tyrobutyricum отличаются невысокой термоустойчивостью: часть из них погибает при температурах выше 80° С. Для полного уничтожения их спор необходима выдержка при 120° С в течение 20 с, при 130° С - 4 с.
Взаимоотношения с другими бактериями. Для сыроделия особенно важны взаимоотношения между маслянокислыми и молочнокислыми бактериями, поскольку в сырах маслянокислое брожение проходит на фоне молочнокислого брожения. При совместном развитии маслянокислых и молочнокислых бактерий на первом этапе молочнокислые бактерии, снижая Eh среды, создают условия для роста маслянокислых. Это продолжается до тех пор, пока pH в совместной культуре не снизится до уровня, при котором рост маслянокислых бактерий полностью подавляется. Поскольку молочнокислые бактерии при достаточно большом начальном количестве и наличии в среде необходимых количеств углеводов быстро смещают pH в незабуференных средах до уровня ниже предельного для роста маслянокислых бактерий, последние в кисломолочных продуктах не размножаются. Иное дело в твердых сырах, pH которых из-за ограниченного содержания лактозы и высокой буферности сырной массы, хотя и приближается к нижней границе для роста маслянокислых бактерий, но не пересекает ее (исключением являются сыры с высоким уровнем молочнокислого брожения, в которых иногда он опускается ниже этой границы - до 4,9).
Попадание в сыр спор маслянокислых бактерий (даже Cl. tyrobu- tyricum) еще не означает его неизбежную порчу в результате маслянокислого брожения, даже если сыр будет выработан по оптимальной с точки зрения формирования органолептических показателей технологии (при выработке сыров по такой технологии создаются наиболее благоприятные условия для развития маслянокислых бактерий, поэтому ее применяют при отсутствии в молоке опасных количеств спор этих микроорганизмов). Наиболее вероятной причиной этого является наличие в сырах среди микрофлоры закваски или «дикой» молочнокислой микрофлоры видов или штаммов, обладающих специфическим антагонизмом по отношению к маслянокислым бактериям.
Две группы лактобактерий обладают специфическим антагонизмом к клостридиям. Первую представляют штаммы молочного лактококка, образующие антибиотик низин. Низин - полипептид, оказывает бактерицидное действие на маслянокислые бактерии преимущественно в период после прорастания спор до начала активного размножения, а также на многие грамположительные бактерии, включая стафилококки и листерии. Пока его не используют в производстве натуральных сыров, поскольку он угнетает развитие кислотообразующих бактерий закваски и тем самым создает условия для развития грамотрицательных бактерий, на которые не действует. Не нашли применения и закваски из образующих низин лактококков, поскольку они обладают низкой кислотообразующей активностью и высокой чувствительностью к бактериофагу.
Пока в сыроделии низин используют только в производстве плавленых сыров. Однако ведутся работы по конструированию штаммов лактококков, продуцирующих низин и обладающих требуемой в сыроделии скоростью развития и кислотообразования в молоке и фагорезистентностью.
Специфический антагонизм других видов молочнокислых бактерий к маслянокислым бактериям обусловлен способностью связывать растворенный в среде кислород с образованием определенных количеств перекиси водорода и, возможно, других радикалов кислорода, токсичных для живых клеток. По-видимому, биологическая целесообразность ассимиляции кислорода молочнокислыми бактериями заключается в снижении Eh среды до требуемого для их активного роста уровня. Молочнокислые бактерии или связывают кислород с минимальным образованием токсичных веществ, или имеют системы защиты, позволяющие им успешно размножаться в присутствии в среде определенного количества перекиси водорода. Маслянокислые бактерии эффективной защиты от токсичных радикалов кислорода не имеют, что и определяет их высокую чувствительность к кислороду. Штаммы молочнокислых бактерий, образующие достаточно высокие концентрации токсичных продуктов ассимиляции 02, ингибируют маслянокислые бактерии при совместном росте. Как правило, специфический антагонизм этого рода присущ видам и штаммам с невысокой кислотообразующей активностью. Наиболее часто специфические антагонистические свойства этого типа встречаются у ме- зофильных лактобацилл, в частности среди штаммов Lbc. plantarum. На рис 6.10 показано образование кислоты и Н202 штаммами Lbc. plantarum (средние данные для нескольких штаммов).
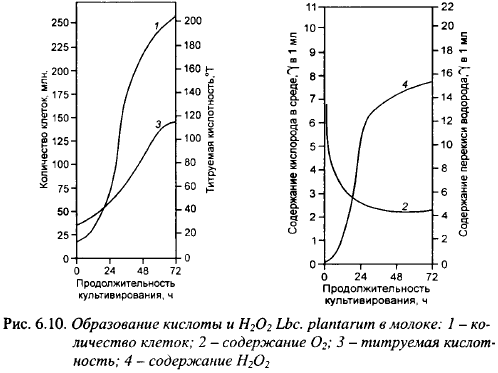
Особенно энергично перекись образовывалась в первые сутки роста. Все штаммы Lbc. plantarum образуют Н202, но в разных количествах: к 48 ч культивирования, когда содержание перекиси в среде достигает максимума, 46,6% исследованных культур образовывали в среднем 2,01 мг/л (0,53-3,6) перекиси, 33,3% - около 12,3 мг/л (8,03-16,9), 13,8% - 28,6 мг/л и 6,8% - более 50 мг/л. Образование перекиси прекращается к началу стационарной фазы развития, что зависит от температуры культивирования: при 11-12° С оно продолжается 12 сут. После достижения максимума содержание перекиси начинает снижаться.
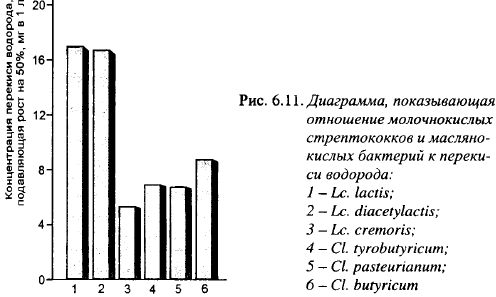
По влиянию на органолептические показатели сыра и скорости кислотообразования в молоке Lbc. plantarum может быть только дополнительным компонентом заквасок для сыра, основу которых должны составлять лактококки. Для того чтобы закваски, в состав которых включают штаммы Lbc. plantarum - антагонисты маслянокислых бактерий, образующие Н202, - сохраняли требуемую активность, лактококки не должны ингибироваться теми концентрациями перекиси, которые образуют лактобациллы. На рис. 6.11 показана чувствительность к Н202 лактококков и маслянокислых бактерий. Из рисунка видно, что концентрация Н202, на 50% подавляющая рост маслянокислых бактерий, почти в два раза ниже ее концентрации, подавляющей на 50% рост молочного и диацетильного лактококков. Чувствительность к перекиси сливочного лактококка почти такая же, как маслянокислых бактерий, поэтому его нецелесообразно вводить в закваски вместе с лактобациллами - антагонистами маслянокислых бактерий. Таким образом, существует диапазон концентраций перекиси водорода, которые в большой степени ингибируют рост маслянокислых бактерий, оказывая незначительное действие на молочный и диацетильный лактококки.
На рис. 6.12 показан совместный рост маслянокислых бактерий и штаммов Lbc. plantarum, образующих в молоке различные количества Н202. Из рисунка видно, что все три проверенные штамма Lbc. plantarum оказывали ингибирующее действие на накопление биомассы маслянокислыми бактериями, степень ингибирования была различной - пропорционально количеству образуемой в среде штаммами лактобацилл Н202 . Наибольшей антагонистической активностью обладал Lbc. plantarum 28, образующий в молоке около 9 мг/л перекиси. Он был использован в заквасках, обладающих антагонистическим действием на маслянокислые бактерии. Рост Cl. tyrobutyricum в зависимости от штамма снижался на 50% в присутствии 2,7-8,8 мг/л Н202.

Максимальные концентрации H202, образуемые в молоке лактококками, Lbc. plantarum, Lbc. acidophilus, Lbc. lactis и Lbc. bulgaricus в зависимости от штамма равнялись соответственно 1,02-6,46; 2,04- 70,04; 13,6-54,4; 14,96 и 10,88 мг/л.
Некоторые штаммы молочного лактококка, Lbc. casei, Lbc. plantarum, Lbc. fermentum, Lbc. lactis, Lbc. helveticus, Lbc. bulgaricus и Lbc. acidophilus, термофильного стрептококка образуют бактериоцины, ингбирующие рост маслянокислых бактерий. Обычно антагонистическое действие этих веществ слишком низкое, чтобы подавить развитие клостридий в сырах с высокой температурой II нагревания.
Имеются материалы, свидетельствующие о стимулирующем действии на маслянокислые бактерии БГКП. В условиях, лимитирующих снижение pH, многие молочнокислые бактерии не подавляют, а стимулируют рост маслянокислых бактерий. Так, из числа проверенных штаммов термофильных молочнокислых палочек в среде, содержание энергетического материала в которой было рассчитано так, чтобы конечный pH ее равнялся 5,2-5,3, рост Cl. tyrobutyricum стимулировали 74,95% штаммов Lbc. helveticus, 62,5% Lbc. bulgaricus, 60% Lbc. lactis; ни один из проверенных штаммов не обладал антагонизмом по отношению к этим микроорганизмам. В этих же условиях при изменении pH среды от 6,7 до 5,2 все испытанные штаммы лактококков, за исключением низинобразующих, стимулировали рост маслянокислых бактерий. Особенно сильное стимулирующее действие оказывали штаммы молочного лактококка. В совместных с лактококками культурах маслянокислые бактерии начинали вегетативный рост с самого начала культивирования. Рост маслянокислых бактерий стимулируют термофильный стрептококк и пропионовокислые бактерии.
