Маслянокислые бактерии - возбудители позднего вспучивания сыров, что подразумевает их размножение во второй половине созревания сыров. Это не совсем так, о чем свидетельствует динамика размножения маслянокислых бактерий, показанная в табл. 6.19 и 6.20 на примере сыров Костромской и Российский. Молоко для выработки сыров после па- стризации инокулировали коллекционными штаммами маслянокислых бактерий из расчета примерно 250 спор/мл. В сырах после прессования концентрация спор маслянокислых бактерий во всех вариантах уменьшилась по сравнению с их концентрацией в смеси, в 2,3-19,2 раза, хотя она должна была увеличиться примерно в 10 раз даже без размножения маслянокислых бактерий, так как почти все споры из смеси переходят в сгусток. Условия выработки сыра не могут вызвать гибель спор, они благоприятны для их прорастания, но проросшие споры теряют устойчивость к нагреванию и не выявляются при определении содержания спор в исследуемом субстрате. В это же время начинается вегетативный рост маслянокислых бактерий, поскольку в молоке после пастеризации вегетативные клетки маслянокислых бактерий отсутствовали, а в сырах после прессования насчитывалось в 1 г (3-14)-103 клеток Cl. tyrobutyricum, 9-103 Cl. butyricum и 2,5-103 Cl. pasteurianum. Как и следовало ожидать, наиболее быстро размножался Cl. tyrobutyricum. Неожиданным оказалось более быстрое размножение маслянокислых бактерий в Российском сыре во время выработки, что, возможно, обусловлено более быстрым развитием в нем молочнокислых бактерий, которые на первом этапе своего развития стимулируют рост маслянокислых бактерий. Таким образом, во время выработки сыра маслянокислые бактерии, как и в лабораторных опытах, размножаются параллельно с молочнокислыми бактериями.

В Российском сыре Сl, tyrobutyricum размножался до 10-суточного возраста, потом количество его жизнеспособных клеток начало уменьшаться, вначале медленно, затем быстро. Количество жизнеспособных клеток в этом сыре в период максимума было ниже, чем максимальное их количество в Костромском сыре, примерно в 16 раз, а в зрелом сыре - в 600 раз, что свидетельствует о более медленном развитии маслянокислых бактерий в Российском сыре во время созревания, и это закономерно, учитывая более высокую активную кислотность этого сыра.
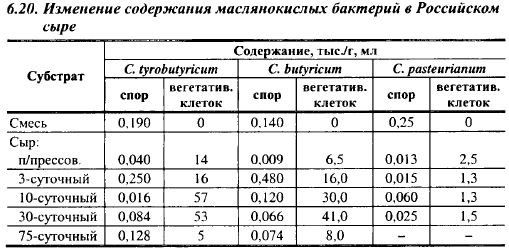
Cl. butyricum размножался в Российском сыре только до 30-суточного возраста, после чего количество его клеток стало уменьшаться. В период максимума содержание жизнеспособных клеток этого вида было несколько ниже, чем максимальное количество клеток Cl. tyrobutyricum. Cl. pasteurianum размножался только во время выработки сыра. Никаких видимых признаков маслянокислого брожения в Российском сыре не было.
В Костромском сыре размножение маслянокислых бактерий прослеживалось до 5-суточного возраста, в сырах 10-15-суточного возраста содержание вегетативных клеток Cl. tyrobutyricum было ниже, чем в 3-5-суточных сырах. Оно снова начало возрастать в сырах, начиная с 25-суточного возраста. В 45-суточных сырах количество клеток Сl. tyrobutyricum было достаточно, чтобы вызвать дефекты в органолептических показателях Костромского сыра: в нем появился прогорклый, слегка салистый вкус, рваные глазки, белесый цвет теста, но вспучивания не было.
Результаты этого опыта и многочисленные последующие наблюдения за развитием маслянокислых бактерий свидетельствуют о трех- фазности маслянокислого брожения в сырах: первая фаза - прорастание спор и вегетативный рост во время выработки и на первом этапе созревания сыра; вторая фаза - отсутствие размножения и даже снижение количества жизнеспособных клеток; третья фаза - вознобновление роста. Продожительность и время наступления каждой фазы вариабельны, что обусловлено особенностями микробиологических и физико-химических процессов в различных сырах.
Наличие первой фазы вполне объяснимо, так как во время выработки и в начале созревания условия в сырной массе наиболее благоприятны для развития маслянокислых бактерий. Труднее объяснить существование второй фазы. Прекращение видимого размножения маслянокислых бактерий, означающее окончание первой фазы, может быть вызвано снижением pH сыра до близкого к нижней границе для их роста. В пользу этого говорит то, что первая фаза кончается, когда pH сыров достигает минимального уровня. После этого pH сыров начинает медленно повышаться за счет протеолиза. Кроме этого, метаболизм маслянокислых бактерий, в частности сбраживание лактатов, может очень медленно продолжаться без деления клеток, в результате чего pH в ближайшем окружении клеток или колоний маслянокислых бактерий будет также медленно повышаться. Возникнет цепная реакция: маслянокислые бактерии медленно повышают pH, сбраживая лактаты, а повышение pH ускоряет сбраживание лактатов. При повышении pH до определенного уровня маслянокислые бактерии начнут размножаться в сыре с достаточно высокой скоростью, и начнется третья фаза. Одной из причин начала активного роста маслянокислых бактерий на третьем этапе может быть увеличение в сыре низкомолекулярных продуктов протеолиза, стимулирующих рост маслянокислых бактерий. Обычно интенсивность протеолиза во вспученных сырах выше, чем в доброкачественных, но это может быть и причиной, и следствием развития возбудителей вспучивания. Третья фаза заканчивается после достижения А в в центре головки сыра нижней границы для роста маслянокислых бактерий или продолжается до конца созревания.
Одной из причин прекращения роста маслянокислых бактерий на первом этапе созревания сыра может быть накопление лактококками в сырной массе специфических антибиотических веществ, но это маловероятно, учитывая очень слабое антибиотическое действие лактококков на маслянокислое брожение, за исключением низинообразующих штаммов, которые в составе заквасок не используются.
Первая фаза маслянокислого брожения всегда будет возникать в сырах, если в молоке есть споры маслянокислых бактерий. Об этом свидетельствует наличие небольших количеств масляной кислоты во всех сырах, даже при отсутствии видимых признаков маслянокислого брожения.
Второй фазы может не быть, если молочнокислое брожение идет слишком медленно и pH сыров не опускается до уровня ниже 5,4-5,5, а молоко содержит много спор Сl. tyrobutyricum или Сl. butyricum. В этом случае маслянокислые бактерии могут вызвать вспучивание сыров даже в 10-15-суточном возрасте за счет сбраживания лактатов и лактозы.
Третьей фазы маслянокислого брожения при наличии второй может не быть, если в молоке содержится небольшое количество спор маслянокислых бактерий (не более 10 спор/г), а pH сыров быстро снижается до 5,2-5,3, или ниже (в соответствии с видом вырабатываемого сыра), и содержание соли в водной фазе центральных слоев головки сыра через 25-30 сут превысит 4,1%, т. е. предельный уровень для роста маслянокислых бактерий при pH сыра (табл. 6.17). В этом случае метаболизм маслянокислых бактерий может быть полностью подавлен или же находиться на низком уровне, недостаточном для повышения pH сыра до требуемой для активного размножения маслянокислых бактерий величины. Чем быстрее pH сыра снижается до 5,3, чем ниже степень загрязнения сыра маслянокислыми бактериями и короче срок созревания сыра, тем больше вероятность того, что третья фаза развития маслянокислых бактерий не наступит и сыр не приобретет дефектов, вызываемых маслянокислыми бактериями.
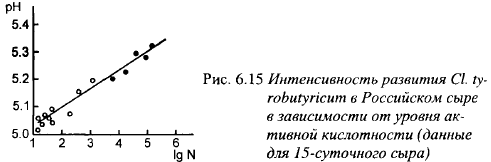
Данные, приведенные в табл. 6.19 и 6.20, свидетельствуют о том, что в сырах маслянокислые бактерии или не образовывали спор, или образовывали их в незначительных количествах, что может быть обусловлено низким pH сырной массы. Следовательно, оценивать размножение маслянокислых бактерий в сыре по содержанию только спор маслянокислых бактерий, как это обычно делают, нельзя. Более точным критерием в этом случае является содержание масляной кислоты в сыре. О решающем влиянии скорости молочнокислого брожения на развитие маслянокислых бактерий в Российском сыре свидетельствуют результаты опыта, представленные на рис. 6.15. Молоко в опыте после пастеризации инокулировали спорами маслянокислых бактерий в количестве 25-250 спор/мл. В этом сыре pH после прессования должен быть равен 5,15-5,25; pH сыров в 15-сугочном возрасте выше 5,3 свидетельствует о низкой скорости молочнокислого брожения. В сырах с таким pH количество клеток маслянокислых бактерий приблизилось к критическому уровню (-2,5-105 г-1) или его превысило. Таким образом, даже при небольшом снижении активности молочнокислого брожения и высоком обсеменении молока спорами маслянокислые бактерии могут вызвать порчу достаточно стойкого к маслянокислому брожению Российского сыра.
Развитие маслянокислых бактерий в сыре в значительной степени зависит от состава микрофлоры закваски. На рис. 6.16 показано содержание маслянокислых бактерий в Костромском сыре, выработанном с традиционной (К) и содержащей штаммы молочнокислых бактерий со специфическим антагонистическим действием на клостридии (О), заквасками.
В К-сыре содержание маслянокислых бактерий равнялось примерно 106 г-1, т. е. превысило критический уровень, в О-сыре оно равнялось З-103 г-1 или было ниже этого уровня. Результаты органолептической оценки (средние по 6 повторностям) приведены в табл. 6.21. Степень выраженности пороков сыра зависела от количества вегетативных клеток маслянокислых бактерий в период максимума. Они выражаются в появлении прогоркло-слащавого вкуса, мажущейся консистенции, белесой окраски сырного теста, рваного, броженого рисунка, переходящего во вспучивание головок. Признаки маслянокислого брожения появляются при содержании в сыре примерно 2-105 г-1 клеток маслянокислых бактерий, вспучивание наступает, когда их количество достигнет нескольких млн./г. Выше указывалось, что размножение маслянокислых бактерий может происходить без видимого газообразования. Таким образом, спектр пороков сыра, вызываемых маслянокислыми бактериями, достаточно широк и не ограничивается поздним вспучиванием.

