Биохимические процессы играют основную роль в формировании качественных особенностей игристых вин, начиная от переработки винограда и кончая обработкой шампанизированного вина. Советские ученые А. И. Опарин, Н. М. Сисакян, А. М. Фролов-Багреев, Г. Г. Агабальянц, А. А. Мержаниан, А. К. Родопуло и др. внесли значительный вклад в установление механизма биохимических и физико-химических процессов, протекающих при формировании игристых вин. В последние годы появились работы по биохимии игристых вин за рубежом.
В биохимической теории процессов формирования игристых вин [3, 121, 122] установлены факторы, оказывающие влияние на формирование специфических свойств игристых вин, выяснены основные закономерности биохимических процессов, протекающих на отдельных стадиях получения этих вин, предложены биохимические способы регулирования процессов формирования игристых вин.
2.1. БИОХИМИЧЕСКИЕ ФАКТОРЫ,
ВЛИЯЮЩИЕ НА ФОРМИРОВАНИЕ ИГРИСТЫХ ВИН
Процесс формирования игристых вин включает комплекс сложных биохимических и физико-химических процессов, на направленность которых оказывают влияние три фактора: ферментативный и химический состав шампанизируемых виноматериалов, биохимические признаки используемых для шампанизации дрожжей, особенности вторичного брожения и выдержки шампанизируемого вина. Все эти факторы взаимосвязаны. Так, многие компоненты виноматериалов, переходя в шампанское, участвуют в сложении его вкуса и букета. Наряду с этим интенсивность и направленность большинства биохимических процессов формирования шампанского зависит от активности ферментов, содержания отдельных химических веществ, величины рН, уровня ОВ-потенциала в виноматериале. Такие вещества, как соли тяжелых металлов, диоксид серы, фенольные и другие соединения, присутствующие в купаже, могут ингибировать или активировать ферменты как при брожении, так и при выдержке. Кроме того, некоторые соединения виноматериалов при вторичном брожении вызывают индукцию синтеза ряда ферментов в дрожжевых клетках, другие компоненты, напротив. выступают в роли репрессоров синтеза внутриклеточных ферментов, т. е. углеводы, органические кислоты, азотистые вещества и другие компоненты виноматериалов участвуют в регуляции биосинтеза ферментов дрожжей и вина. В зависимости от того, присутствует ли кислород в вине, каков качественный и количественный состав отдельных аминокислот, спиртов и других веществ в виноматериале, в одних и тех же дрожжевых клетках будет синтезирован различный комплекс ферментов.
Активность внутриклеточных ферментов дрожжей в свою очередь определяет характер биохимической трансформации составных веществ виноматериалов при шампанизации. От индуцированных и конститутивных ферментов дрожжевых клеток зависит синтез высших спиртов, альдегидов, сложных эфиров и других компонентов, определяющих вкус и букет шампанского. Химический состав дрожжей также влияет на состав шампанского, так как многие компоненты переходят из клеток в вино при автолизе: физиологические и биохимические признаки дрожжей, например бродильная способность, холодостойкость, структура осадка и др., сказываются и на процессе шампанизации.
Особенности шампанизации в бутылках и резервуарах, условия контакта шампанизируемого вина с дрожжами, температурные режимы, продолжительность отдельных стадий, избыточное давление углекислоты и пр. существенно отражаются и на метаболизме дрожжей, и на биохимических реакциях в шампанизируемом материале. Одной из основных особенностей процесса шампанизации является проведение вторичного брожения и последующей выдержки шампанизируемого вина с дрожжами. Исключение выдержки при резервуарном способе приводит к тому, что в шампанизируемом вине не протекают ферментативные реакции с участием автолизирующихся дрожжевых клеток и другие процессы, формирующие специфические особенности шампанского и игристых вин.
Рассмотрим подробнее роль отдельных факторов в процессах формирования игристых вин.
Виноматериалы
Состав виноматериалов отличается большим многообразием. В них присутствуют азотистые вещества, ферменты, фенольные, фосфорные, ароматобразующие и другие соединения.
Ферменты, функционирующие в шампанских виноматериалах, относятся к двум классам: оксидоредуктазам и гидролазам. Обнаруженные в винах малат-, глютамат-, аланин-, сукцинат- и лактатдегидрогеназы, по-видимому, принимают участие в транспорте водорода (электронов) и восстановлении компонентов букета и вкуса. Гидролиз белковых веществ катализируется в вине протеиназой и пептидазой. Эти ферменты различаются по оптимальным условиям действия, причем протеиназная активность преобладает над пептидазной, вследствие чего протеолиз главным образом доходит до пептидов. Высока активность в виноматериалах и шампанском β-фруктофуранозидазы, которая помимо ускорения инверсии сахарозы синтезирует в шампанских винах β-этилфруктозид. В винах найдены гидролазы, катализирующие синтез этиловых эфиров капроновой, каприловой, молочной и других карбоновых кислот и, следовательно, принимающие непосредственное участие в формировании вкуса и букета виноматериалов и шампанского. Ускорение ферментами вина многих реакций подтверждает необходимость применения технологии, обеспечивающей накопление ферментов в виноматериалах и шампанском.
Азотистые вещества представлены белками, пептидами, аминокислотами, меланоидинами, пуриновыми и пиримидиновыми основаниями, аммиаком и другими соединениями. Лиофилизованные нативные белки вина – хлопьеобразное вещество белого цвета, включающее несколько фракций различной молекулярной массы. В белках преобладают аланин, треонин, глютаминовая и аспарагиновая кислоты, гликокол и серии. Пептиды вина состоят из аспарагиновой и глютаминовой кислот, треонина, аланина, лейцина и изолейцина, лизина, аргинина, гистидина и цистеина. На долю пептидов в виноматериалах приходится от 12 до 60 % общего содержания азотистых веществ. Выделение и определение строения индивидуальных пептидов показало, что в игристых винах присутствуют пептиды, содержащие от 5 до 30 аминокислот. В табл. 1 приведена первичная структура пептидов вина. Как видно, в наибольшем количестве в вине содержатся пептиды, состоящие из 5, б, 7 и 8 аминокислотных остатков.
Свободные аминокислоты составляют примерно 1/3 азотистых соединений шампанских вин. В их составе находится 22 аминокислоты, причем до 40 % приходится на пролин.
Таблица 1

Фенольные соединения участвуют в окислительно-восстановительных процессах, реакциях полимеризации и поликонденсации, взаимодействуют с азотистыми и другими веществами вина. Фенольные соединения влияют на вкус, цвет, букет и прозрачность вина. Согласно работам С. В. Дурмишидзе, П. Риберо-Гайона, Г. Г. Валуйко и др. в винограде и вине содержатся мономерные и полимерные полифенолы. Из мономерных выделяются производные бензойной кислоты (С6–С1), коричной кислоты (С6–С3) и флавоноиды (С6–С3–С6). В наибольшем количестве в вине содержатся флавоноиды, включающие катехины, антоцианы, лейкоантоцианы, флавонолы, флавоны. Из полимерных выделяются танины, полимеризованные флавоноиды, лигнин. Содержание в винограде и вине отдельных групп полифенолов приведено в табл. 2 [49].
Таблица 2

Каждая из групп фенольных соединений содержит несколько индивидуальных веществ различного строения. Группа катехинов включает помимо катехинов галлокатехины, их стереоизомеры, эфиры галловой кислоты. Катехины – наиболее восстановленные соединения из флавоноидов. Они легко окисляются и полимеризуются.
Антоцианы являются красящими веществами, в винограде они содержатся в виде гликозидов и соединений с ароматическими и алифатическими кислотами. Основным красящим веществом красных сортов винограда является моногликозид мальвидина. Антоцианы легко полимеризуются, вызывая появление коричневато-буроватых оттенков.
Лейкоантоцианы также окисляются и полимеризуются. Полимеры лейкоантоцианов вызывают потемнение белых вин.
Танины бывают гидролизуемыми и негидролизуемыми. Гидролизуемые танины являются полиэфирами фенолкарбоновых кислот и сахаров. Конденсированные танины включают продукты полимеризации в основном катехинов и лейкоантоцианов. Танины усиливают окраску антоцианов [19], играют важную роль в окраске выдержанных красных вин [103]. В молодых винах молекулярная масса танинов составляет 500–800, при созревании вин она возрастает до 3000–4000, а при старении вновь снижается из-за выпадения в осадок полимерных форм.
Фенольные соединения играют важную роль в процессах, проходящих при получении и обработке белых и красных виноматериалов для игристых вин и шампанизации. Они оказывают большое влияние на метаболизм дрожжей.
В шампанских виноматериалах содержатся фосфорные соединения, главным образом растворимые в трихлоруксусной кислоте. Основная масса их представлена ортофосфорными соединениями, количественно меньше лабильных и стабильных соединений фосфора и полифосфатов. В процессе шампанизации и выдержки изменяются соотношения отдельных форм фосфорных соединений. Содержание органических форм фосфорных соединений может в определенной мере служить характеристикой срока выдержки шампанского.
В составе липидов виноматериалов и шампанского идентифицировано 6 фракций: моноацил-, диацил- и триацилглицерины, фосфоглицериды, неэтерифицированные (свободные) жирные кислоты и стеролы [3].
Фракция фосфоглицеридов в липидах вина, которая является наибольшей, состоит из пяти субфракций: фосфатидные кислоты, полиглицерофосфатиды, фосфатидилхолин, фосфатидилэтаноламин и лецитины. Состав кислот в отдельных фракциях существенно различается, заметно колеблется и соотношение насыщенных и ненасыщенных кислот. Липиды шампанского построены из тех же кислот, что и липиды виноматериалов, однако в иных пропорциях: в шампанском доля ненасыщенных кислот с 20 углеродными атомами и бегеновой кислоты С22:0 в 2–3 раза выше, чем в купаже. Следует заметить также, что качественный состав кислот липидов вина и дрожжей идентичен, что свидетельствует о происхождении многих липидов вина из дрожжевых клеток. В виноматериалах и шампанском найдено липидов 60–220 мг/л.
Состав веществ, обусловливающих вкус и букет виноматериалов и шампанского, очень сложен. Среди легколетучих соединений наибольший удельный вес (20–500 мг/л) приходится на этиллактат, метил-3 бутанол-1, метил-2 бутанол-1, пропанол-1, метанол. Во фракции веществ с высокой температурой кипения преобладают фенилэтиловый спирт, этиловые эфиры капроновой, каприловой, лауриновой, миристиновой и пальмитиновой кислот, диэтилмалат, диэтилсукцинат, кислый этиловый эфир янтарной кислоты, жирные кислоты.
В составе газов из надвинного пространства – паров шампанского, непосредственно участвующих в формировании букета, обнаруживается около 30 соединений, из которых основными являются этанол, метанол, этилацетат, изоамиловый и изобутиловый спирты, ацетальдегид, н-пропанол, этилформиат, изоамилацетат. В композиционном составе летучих веществ виноматериалов и их паров имеются существенные различия.
Хромато-масс-спектрометрическое исследование букетистых веществ вин позволило установить, что для игристых и белых столовых вин в отличие от вин остальных типов характерно повышенное содержание сложных эфиров и углеводородов.
Сравнение состава летучих веществ различных типов вин показывает, что аромат определяется не одним специфическим веществом, а определенной композицией большого числа пахучих соединений, в том числе этилацетата, изоамилацетата, этилкапроната, этилкаприлата, изобутанола, изоамилового спирта, ацетальдегида и некоторых других, которые содержатся во всех образцах вин и таким образом составляют основу винного запаха, на фоне которого отдельные группы летучих соединений создают характерный аромат данного типа вина.
Оптимальный состав купажа до сих пор окончательно не установлен. Однако выяснено, что в шампанских виноматериалах не должно содержаться более 20 мг/л свободного ацетальдегида и более 0,8 мг/л диацетила, а повышение их концентрации вызывает появление тона окисленности [86].
Следует создавать условия для накопления в виноматериалах ряда ферментов: дегидрогеназ, гидролаз эфиров карбоновых кислот; букетистых веществ: этиллактата, 2-фенилэтилового спирта, высококипящих этиловых эфиров жирных кислот, терпенов. Отрицательное влияние на букет виноматериалов и шампанского оказывают изопентанол и изобутанол, количество которых в парах вин бывает достаточно велико. В шампанском усиленному переходу высших спиртов в пары способствует также выделяющийся диоксид углерода. Поэтому в случае повышения концентрации изоамилового и изобутилового спиртов в вине одновременно значительно увеличивается их содержание в парах, что резко ухудшает букет виноматериалов и шампанского. Именно по этим соображениям содержание указанных спиртов в шампанском должно ограничиваться. Для этого процесс брожения желательно проводить с помощью дрожжей, продуцирующих небольшие количества изобутанола и изопентанола.
В купаже необходимо ограничивать содержание ацетальдегида, этилацетата, изоамилового спирта, фенольных веществ, ухудшающих качество шампанского.
Следует также стремиться к снижению оптической плотности купажа при λ=400 и 280 нм, установить требования относительно величины ОВ-потенциала и восстановительной способности шампанских виноматериалов.
Шампанские виноматериалы различаются по активности отдельных ферментов, составу различных форм азотистых, фосфорных соединений, органических кислот, спиртов, альдегидов, эфиров и других компонентов. Многие из этих веществ переходят в шампанское в неизменном виде и являются как бы основой формирования его вкуса и букета. Другие трансформируются в процессе вторичного брожения и выдержки. Купаж служит той средой, в которой протекает жизнедеятельность дрожжевых клеток и возникают многообразные биохимические превращения. Вот почему в зависимости от состава купажа изменяются микробиологические и биохимические процессы при шампанизации.
Оптимизация состава виноматериалов – достаточно сложный вопрос, который должен решаться на научной основе. При этом необходимо принимать во внимание и то обстоятельство, что состав виноматериалов должен обеспечивать оптимальные условия для жизнедеятельности дрожжей и протекания биохимических процессов при шампанизации.
Дрожжи
Состав образующихся при шампанизации продуктов зависит от активности ферментов, катализирующих отдельные этапы спиртового брожения, обмена белков, жиров, и других биологических процессов дрожжевой клетки. При шампанизации виноматериалов штаммами дрожжей получаются шампанские вина с неодинаковым содержанием высших спиртов, эфиров, жирных кислот, альдегидов, с различными физико-химическими показателями и органолептическими свойствами [3]. Поэтому учет генетических особенностей дрожжевых клеток и научно обоснованная селекция дрожжей позволяют получать шампанское с высококачественными вкусом и букетом, хорошими игристыми и пенистыми свойствами. На это неоднократно указывали А. М. Фролов-Багреев и Г. Г. Агабальянц. При использовании тех или иных дрожжей следует принимать во внимание снижение ОВ-потенциала вина, образование высших спиртов и других побочных и вторичных продуктов брожения, скорость автолиза дрожжей и др. В этих целях необходимо более подробно исследовать роль конститутивных ферментов дрожжей.
Наряду с конститутивными в клетках дрожжей образуются индуцируемые ферменты, синтез которых определяется условиями культивирования. Наличие одних соединений в культуральной жидкости стимулирует, других – тормозит синтез отдельных внутриклеточных ферментов. Например, при росте на среде с глютаминовой кислотой в дрожжевых клетках индуцируется синтез НАД-глютаматдегидрогеназы, напротив, на среде с аммонием наблюдается репрессия синтеза этого фермента в клетках. Культивирование дрожжей на средах с различным содержанием сахара (2; 5; 10 %) и аэрация существенно сказываются на метаболизме шампанских дрожжей. Состав питательной среды и режимы производства оказывают влияние на активность митохондриальных ферментов и химический состав дрожжевых клеток. В связи с этим становится понятной необходимость установления оптимальных условий культивирования дрожжей для производства шампанского.
Вещества, содержащиеся в клетках дрожжей, принимают непосредственное участие в биохимических процессах при шампанизации. Кроме того, некоторые компоненты дрожжевых клеток переходят в шампанское при автолизе и изменяют его состав. На основании исследований биохимии процесса автолиза дрожжей в вине нами предложен механизм этого процесса, в основу которого положены представления о роли внутриклеточных изменений, возникающих при выдержке и термической обработке, и значении условий существования дрожжевых клеток в вине. Показано, что имеются существенные различия в характере биохимических превращений при автолизе дрожжей методами длительной выдержки при 15–20 °С, обработки холодом и тепловой обработки. Эти различия сказываются на интенсивности перехода отдельных компонентов дрожжей и их превращении. Так, при обработке холодом в вине значительно повышается активность многих ферментов. В случае нагревания вина с дрожжами наряду с выходом ферментов из дрожжей происходит их частичная инактивация. Вместе с тем при тепловой обработке более интенсивно протекают процессы распада дрожжевой клетки, перехода из нее азотистых и фосфорных соединений, одновременно возникают дальнейшие превращения компонентов дрожжей и вина, ускоряются химические реакции. Различия в процессах при выдержке, обработке холодом и теплом вина с дрожжами значительно сказываются на качественном и количественном составе компонентов букета.
Дрожжевые клетки при автолизе выделяют в вино около 40 компонентов, обусловливающих вкус и букет вина, в том числе дегидрогеназы, протеиназу, β-фруктофуранозидазу, эстеразы, около 20 аминокислот, другие азотистые вещества, кислоторастворимые фосфорные соединения, маннан, липиды. Таким образом, в результате жизнедеятельности дрожжей при вторичном брожении и связанной с этим трансформации компонентов виноматериалов формируются основные свойства шампанского.
Процесс шампанизации
Процесс шампанизации отличается сложными биохимическими и физико-химическими превращениями различных компонентов вина, причем в направленности биохимических процессов при бутылочной и резервуарной шампанизации имеются определенные различия (см. п. 2.2).
В табл. 3 приведены технологические параметры трех способов производства игристых вин. Они даются с определенными интервалами в связи с тем, что в отдельных странах, а нередко в одной и той же стране, приняты различные модификации способов, режимы, сроки. Несмотря на некоторую условность, приведенные данные позволяют с определенной достоверностью оценить технологические особенности различных способов. Наряду с известными в таблице фигурируют новые показатели, полученные расчетным путем.
Таблица 3

Из приведенных данных видно, что бутылочный, способ в 15–50 раз более продолжителен, чем резервуарный. При бутылочном и периодическом резервуарном способах происходит размножение дрожжей в том же сосуде, а при непрерывном способе размножение дрожжевых клеток осуществляется в отдельных аппаратах. Продолжительность вторичного брожения различается менее заметно: при резервуарных способах она сокращена в основном в 1,5–3,0 раза. Особенно велики различия в длительности 3-го и 4-го периодов. Так, 3-й период, характеризующийся интенсивными ферментативными процессами, в периодическом резервуарном способе в основном отсутствует; лишь в некоторых странах практикуют выдержку шампанизируемого в резервуарах вина с дрожжами. При непрерывной шампанизации в батарее аппаратов выдержка шампанизируемого вина с дрожжами в резервуаре с насадкой продолжается 2 сут, что в 100–150 раз меньше, чем при бутылочном Способе. Шампанизация в двухъемкостной системе предусматривает удлинение этой стадии до 8 сут. Что касается 4-го периода, то в некоторых странах он исключен, в других его продолжительность колеблется от 2 до 5 лет.
При бутылочном способе проводятся ремюаж и дегоржаж, связанные с механическими воздействиями, охлаждением, введением ликера и проникновением кислорода. При резервуарном способе интенсивное воздействие на вино оказывают обработка холодом, введение ликера, фильтрация и розлив. Эти приемы можно отнести к нежелательным, но необходимым.
Имеются некоторые различия между температурными режимами процесса шампанизации в бутылках и резервуарах, в основном при периодическом способе. Более заметна разница в концентрации дрожжевых клеток в шампанизируемом вине: при непрерывном способе она примерно в 3 раза ниже, чем при бутылочном и резервуарном периодическом, но имеется тенденция к ее повышению; самая высокая концентрация клеток дрожжей – в резервуарах с насадками в установках непрерывной шампанизации.
Большой интерес представляет показатель ферментирования дрожжами (ФД), получаемый произведением концентрации клеток дрожжей на продолжительность процесса. Наиболее высокие значения ФД – при бутылочном способе, особенно в 3-й и 4-й периоды. Близки к ним абсолютные величины ФД при непрерывном способе, тогда как при резервуарном периодическом способе величины ФД в несколько раз ниже. Еще более велика разница в величинах показателя ВФД (возрастного ферментирования дрожжами), представляющего собой произведение показателя ФД на средний возраст дрожжевых клеток при шампанизации. По этому показателю резервуарный периодический способ в несколько десятков раз уступает бутылочному и непрерывному резервуарному. Указанное свидетельствует о том, что ферментативные процессы наиболее выражены при бутылочной шампанизации, несколько меньше при непрерывной и минимальны при периодической резервуарной шампанизации. Предложенные показатели показывают возможные пути моделирования процессов, характерных для бутылочного способа, в условиях резервуарных способов производства игристых вин.
Следует отметить, что продолжительность процесса имеет не только физический смысл, но и определяет степень протекания химических и биохимических процессов при шампанизации. Из химических процессов при длительной выдержке шампанизированного вина в бутылках проходят окислительно-восстановительные, этерификации, карбониламинные, полимеризации и поликонденсации. Очевидно, что сокращение длительности выдержки или исключение этого этапа при резервуарных методах приводит к тому, что указанные реакции протекают не в полной мере, вследствие чего не заканчиваются процессы формирования выдержанных тонов в игристых винах. В определенной мере этот недостаток может быть исправлен использованием в купаже выдержанных виноматериалов.
Из медленно протекающих биохимических процессов необходимо отметить автолиз дрожжей, реакции, катализируемые оксидоредуктазами и гидролазами. При кратковременной выдержке указанные процессы окажутся незавершенными.
Таким образом, технологические особенности процесса шампанизации оказывают решающее влияние на биохимические процессы формирования игристых вин.
2.2. ОСНОВНЫЕ ЗАКОНОМЕРНОСТИ БИОХИМИЧЕСКИХ ПРОЦЕССОВ
ПРИ ПОЛУЧЕНИИ ИГРИСТЫХ ВИН
Получение игристых вин, начиная от дробления винограда и кончая получением шампанизированного вина, включает ряд стадий, различающихся характером и интенсивностью биохимических процессов.
Биохимические процессы, протекающие при мацерации винограда
Процессы, происходящие при дроблении винограда и мацерации мезги, имеют различную направленность в зависимости от технологии. При получении белых виноматериалов для игристых вин виноград дробят, быстро отделяют сок с помощью прессов и мезгоотсекателей.
Прессование целых гроздей винограда по «шампанскому» способу исключает измельчение составных частей грозди, переход из гребней и кожицы танина, полифенолов, оксидаз и других веществ, в результате чего снижаются окислительные процессы и образуется меньше взвешенных частиц.
При производстве мускатных виноматериалов мезгу настаивают при определенной температуре. В этом случае происходит мацерация сока вследствие автолиза клеток ягод, диффузия и экстракция эфирных масел.
Получение красных виноматериалов предусматривает брожение на мезге, в результате чего бродящее сусло обогащается дубильными, красящими и другими веществами кожицы. При тепловой обработке мезги при температуре 40–45 °С происходит тепловой автолиз клеток стенок и диффузия красящих веществ в сок.
В технологии крепленых виноматериалов используется прием спиртования мезги. В этом случае добавление спирта к мезге обусловливает быстрое отмирание клеток и улучшает экстракцию эфирных масел винограда.
В винограде активны протеиназы [3], эндополиметилгалак- туроназа, Сх-фермент [29] и другие гидролитические ферменты, которые ускоряют распад внутриклеточных структур ягод, вследствие чего экстрактивные вещества и эфирные масла переходят в сок. Часть эфирных масел, в частности терпеноиды, находятся в «связанном» состоянии в виде гликозидов [126] и при действии гликозидаз в процессе мацерации мезги переходят в свободное состояние и диффундируют в сок. Благодаря такому способу переработки винограда сусло обогащается эфирными маслами, которые придают ему сортовой аромат.
В различных частях виноградной ягоды сорта Мускат содержится следующее количество терпенов (в %) [127]:

Таким образом, при мацерации мезги сок обогащается терпенами, но одновременно изменяется их соотношение.
Процессы, протекающие при отстаивании сусла
При отстаивании сусла протекают физические, физико-химические и биохимические процессы. К физическим процессам относятся оседание обрывков ягоды, различных взвешенных частиц, растворение кислорода в соке. Среди физико-химических процессов следует выделить взаимодействие дубильных веществ с белками, коагуляцию белков, адсорбцию оседающими частицами различных компонентов, диффузию дубильных, красящих веществ, эфирных масел, азотистых веществ из взвешенных частиц в сок. К химическим и биохимическим процессам, протекающим при отстаивании сусла, относятся окисление полифенолов под действием полифенолоксидазы, окисление аскорбиновой кислоты под действием аскорбинатоксидазы, окисление различных органических веществ (кислот, полифенолов, спиртов) под действием пероксидазы, окисление кислородом воздуха ненасыщенных компонентов вина (редуктонов) с образованием органических перекисей, гидролиз белковых веществ под действием протеиназы, гидролиз пектиновых веществ, ускоряемый пектинолитическими ферментами, гидролиз гликозидов под действием гликозидаз, взаимопревращение органических кислот и др.
Учитывая, что ферменты виноградной ягоды расположены главным образом в твердых частях, активность ферментов в соке невелика, а во взвешенных частицах, в мякоти и кожице повышена, ферментативные процессы при отстаивании сусла протекают медленнее, чем при мацерации мезги. Чем больше в сусле взвешенных частиц, тем интенсивнее протекают ферментативные процессы.
В процессе отстаивания сусла проводят его сульфитацию, в результате чего интенсивность ферментативных процессов снижается. Диоксид серы является ингибитором ферментов (так, активность полифенолоксидазы снижается на 80 % пои введении до 100 мг/л SО2 [129]), антиоксидантом (предотвращает окислительные процессы) и антисептиком (предотвращает развитие микрофлоры).
В последние годы при отстаивании сусла практикуют обработку бентонитом, который вызывает осаждение белковых веществ, а следовательно, и ферментов виноградного сусла, вследствие чего снижается интенсивность ферментативных процессов.
Если не проводить сульфитацию или проводить ее в незначительных дозах, активные пероксидаза и полифенолоксидаза ускоряют окислительные процессы, происходит окисление полифенолов в хиноны, окисление ненасыщенных соединений в перекиси, образование пероксида водорода и окисление им с участием пероксидазы органических кислот, аминокислот, полифенолов и других компонентов, т. е. происходят интенсивные окислительные процессы. В начальный период в сусле содержится много редуктонов. По мере окисления редуктонов появляется все больше окисленных форм и все меньше восстановленных, в сусле накапливаются хиноны, которые начинают полимеризоваться и давать темноокрашенные конденсированные продукты. Эти продукты придают суслу бурую или темно-коричневую окраску, а шампанский виноматериал из такого сусла получается пониженного качества.
Вместе с тем сусло, полученное в анаэробных условиях в атмосфере СО2, дает вино, склонное к оксидазному кассу [129]. Это вызвано тем, что в данном случае не инактивируется фермент полифенолоксидаза. Инактивирование его происходит лишь при размножении дрожжевых клеток, которые способны его адсорбировать и гидролизовать до аминокислот.
В процессе отстаивания с сульфитацией происходят следующие изменения активности ферментов и содержания азотистых веществ:

Виноматериал, полученный из сусла, осветленного отстаиванием, отличается более высоким качеством, имеет более тонкий и интенсивный аромат, менее окрашен, содержит меньше полифенолов, ионов железа, оксидаз, меньше высших спиртов и больше эфиров, чем виноматериал, полученный из сусла, не подвергавшегося отстаиванию [136]. Удаление при отстаивании взвешенных частиц, служащих центрами десорбции СО2, и дрожжевых клеток способствует замедлению брожения.
Таким образом, отстаивание сусла, при котором происходят важные физико-химические и биохимические процессы, является обязательным при производстве шампанских виноматериалов.
В Шампани применяют невысокие дозировки SO2, так как при дозах более 80 мг/л виноматериалы получаются грубее во вкусе, развитие букета задерживается, тормозится яблочно-молочное брожение; повышенная сульфитация приводит к большому накоплению в вине уксусного альдегида, обусловливающего тона окисленности.
Процессы, протекающие при брожении сусла
Брожение вызывает интенсивные биохимические процессы превращения многих компонентов сусла. Размножающиеся дрожжи потребляют азотистые вещества, ассимилируют многие аминокислоты, витамины, адсорбируют белки и ферменты винограда. В первый период брожения концентрация большинства компонентов сусла снижается. Во второй половине брожения содержание ряда компонентов в бродящем сусле возрастает. Накапливающиеся вещества не являются составными частями винограда, они синтезируются и выделяются в вино дрожжами. Это относится и к ферментам. Так, при брожении активность большинства ферментов сусла снижается (табл. 4) вследствие адсорбции их размножающимися дрожжами. К концу брожения из клеток в среду переходят многие ферменты. Исключение составляет о-ДФО, которая не обнаруживается в вине. Учитывая, что при адсорбции ферментов и белков дрожжевыми клетками одновременно происходит их гидролиз протеиназами, была высказана мысль о необратимой инактивации о-ДФО дрожжами и о необходимости их размножения. При незначительном доступе воздуха в сусло или при переработке винограда в атмосфере СО2 торможение размножения может привести к сохранению активности о-ДФО и, как следствие, к потемнению виноматериалов.
Таблица 4
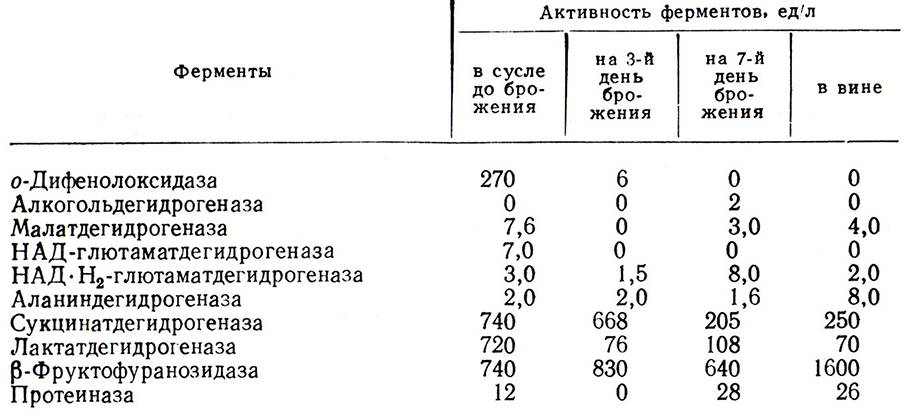
Немаловажное значение при брожении имеют эстеразы, под действием которых синтезируются этиловые и другие эфиры жирных кислот, причем в концентрациях, намного превышающих их равновесные. Из других процессов следует отметить дезаминирование аминокислот дрожжами, вследствие чего синтезируются высшие спирты и жирные кислоты. Образование основных, вторичных и побочных продуктов брожения существенно трансформирует вкус и аромат бродящего сусла. Важнейшую роль при этом играют вид дрожжей, условия брожения и состав сырья.
Особенно интенсивно ассимилируются компоненты сусла при получении мускатных виноматериалов для Асти Спуманте путем многократно повторяющихся операций забраживания сусла и отделения дрожжей. Этот прием, называемый биологическим азотопонижением, приводит к потреблению 58–69 % азотистых веществ сусла [138]. Одновременно дрожжами ассимилируются витамины, микроэлементы и другие вещества сусла, являющиеся факторами роста дрожжевых клеток. Очевидно, что происходит не только ассимиляция компонентов сусла, синтез из них веществ, необходимых для построения новых клеток, но и образование и выделение дрожжами различных продуктов метаболизма, многие из которых (спирты, кислоты, альдегиды, аминокислоты и др.) изменяют состав сусла и угнетают дрожжи.
При получении сухих виноматериалов также заметно падает содержание азотистых веществ – на 40–50 %. В крепленых виноматериалах, получаемых остановкой брожения спиртованием, содержание азотистых веществ снижается на 20–25 %.
Содержание фенольных веществ существенно зависит от режима брожения. Так, при получении красных виноматериалов брожением мезги оптимальной является температура 28–32 °С [19]. Испытание термовинификации при получении виноматериалов для красных игристых вин показало, что нагревание мезги следует проводить при 50–55 °С в течение 8 ч [63].
В процессе брожения сусла ряд компонентов эфирных масел винограда улетучивается вместе с выделяющимся СО2, некоторые трансформируются. В ходе брожения исчезают гексенол, гексаналь, ацетоксибутаналь. Такие альдегиды, как метилфурфурол, бензальдегид, коричный, фенилуксусный, изомасляный, масляный, изовалериановый, гептиловый, превращаются в соответствующие спирты.
В шампанских виноматериалах общее число ароматобра- зующих соединений снижается в процессе брожения с 42 до 30, в то же время суммарная масса компонентов эфирных масел увеличивается в 20–30 раз [88]. Такие соединения, как цис-гексен-З-ол-1, цис-гексен-З-илацетат, цис-гексен-3-илкаприлат, терпеновые углеводороды, восстанавливаются в соответствующие насыщенные соединения. Например, цис-гексен-З-ол-1 восстанавливается в гексанол. При брожении значительно уменьшается содержание гераниола, α-терпинеола, нерола и линалоола. Одновременно появляются фарнезол, этиллактат, этилсукцинат, этилкаприлат, этиллаурат и этилмиристат, сильно увеличивается количество этилацетата, изоамилового спирта, 2,3-бутандиола, ацетата 2-фенилэтилового спирта и 2-фенилэтилового спирта. Механизм образования глицерина, уксусного альдегида, пировиноградной кислоты, уксусной, янтарной, лимонной и молочной кислот хорошо известен. Б вопросе же о путях образования ацетоина, 2,3-бутандиола, диацетила, высших спиртов и эфиров единого мнения нет. Образование высококипящих эфиров при выдержке вина катализируется главным образом ферментными системами винных дрожжей.
Качественный и количественный состав летучих продуктов брожения зависит от используемых штаммов дрожжей. Дикие дрожжи синтезируют больше эфиров и меньше высших спиртов, чем винные дрожжи. Большой интерес вызывает явление различного соотношения н-пропанола, изобутанола и изоамилового спирта в винах, полученных брожением на различных штаммах [3]. Указанное, вероятно, обусловлено неодинаковой активностью ферментных систем клетки и может быть использовано как биохимический тест для характеристики отдельных таксономических групп дрожжей.
Процессы, протекающие при обработке виноматериалов
Основным требованием, предъявляемым к процессам обработки виноматериалов, является предотвращение их аэрации. Газохроматографические исследования показали [2], что аэрация вина вызывает глубокие и необратимые изменения состава компонентов букета. В процессе аэрации и окисления наблюдается значительное снижение концентрации этилацетата, этиллактата, этилпеларгоната и полное исчезновение этилизовалерианата, изоамил ацетата, этилкапроната, этилкаприлата. Одновременно накапливаются ацетальдегид и диэтиловый спирт. Присутствующие в парах сложные эфиры в определенной концентрации придают мягкость и гармоничность букету и вкусу вина. Отсутствие их в аэрированных и окисленных винах и увеличение в композиционном составе букета высших спиртов и ацетальдегид а вызывает грубость и разлаженность букета и вкуса. При окислении в виноматериале почти в 3 раза увеличивается содержание уксусного, изомасляного и изовалерианового альдегидов [2], несколько уменьшается концентрация изобутилового, изоамилового и фенилэтилового спиртов, этилкапроната, изоамилкапроната и других эфиров. Исчезновение эфиров объясняется их улетучиванием при аэрации вина. Увеличение содержания альдегидов и уменьшение содержания спиртов связано с химическим окислением последних. При окислении вина альдегиды образуются в основном из спиртов и в меньшей мере из аминокислот. Реакция окисления спиртов в вине ускоряется полифенолами, ионами Fe2+, Fe3+, Cu1+, Cu2+. При выдержке виноматериала в течение 1,5 мес с периодическим введением О2 через 3–4 сут также отмечается окисление спиртов, накопление альдегидов, исчезновение многих сложных эфиров.
Анаэробная выдержка виноматериалов после открытой переливки характеризуется двумя стадиями. На 1-й происходят ассимиляция растворенного в вине кислорода, окисление спиртов, образование альдегидов. Бескислородный режим 2-й стадии способствует превращению альдегидов, этерификации высших спиртов, накоплению сложных эфиров, благодаря чему качество виноматериалов улучшается. Продолжительность выдержки виноматериалов, содержащих кислород, должна быть не менее 1 мес.
Одним из основных требований, предъявляемых к хранению и обработке мускатных виноматериалов, является сохранение в букете и вкусе специфических веществ мускатного аромата. Ухудшение вкуса мускатных игристых вин связано в основном с окислением виноматериалов и внесением избыточных доз SO2. Так, при общем содержании сернистой кислоты в мускатных мистельных виноматериалах 200–300 мг/л в игристом вине накапливается 100 мг/л и более ацетальдегида, затрудняется проведение вторичного брожения. С другой стороны, SO2, будучи антиоксидантом, защищает вещества, дающие мускатный аромат, от окисления, н его присутствие необходимо. Однако при свободном доступе воздуха даже завышенные дозы SO2 не предотвращают окисления вина, потерю аромата и преждевременное забраживание. При анаэробном хранении снижается ОВ-потенциал, сохраняется мускатный аромат, вино становится гармоничным, более зрелым и не забраживает даже при 20 °С. Анаэробное хранение биологически нестойких виноматериалов при пониженных температурах позволяет на 30–40 % снизить дозы вносимого в них SO2.
Мускатные виноматериалы, получаемые на поточных линиях, имеют несколько повышенное содержание солей металлов (от 7 до 20 мг/л), катализирующих процессы окисления. Поэтому при поступлении виноматериалов на завод игристых вин их необходимо сразу же обработать ЖКС, а при необходимости удалить часть протеинов бентонитом.
После приемки, обработки ЖКС, оклейки и фильтрации мускатных виноматериалов содержание в них растворенного кислорода повышается до 6–8 мг/л. Однако при дальнейшем анаэробном хранении кислород ассимилируется и последующее хранение виноматериалов происходит в бескислородных условиях при низком уровне ОВ-потенциала.
В отличие от производства Советского шампанского, где предусматривается резервное накопление виноматериалов (обработанных купажей), мускатные виноматериалы для игристых вин должны купажироваться незадолго (не ранее чем за 5–10 дней) перед вторичным брожением.
Подготовка к шампанизации виноматериалов включает 2 этапа: получение купажа с гармоничным вкусом и букетом и получение бродильной смеси (удаление кислорода из купажа, снижение ОВ-потенциала, термообработка купажа, обогащение его компонентами дрожжевых клеток, растворение ликера).
При биологической ассимиляции кислорода происходит быстрое потребление кислорода, значительное падение ОВ-потенциала, увеличение восстановительной способности и скорости образования свободных и связанных альдегидов. При деаэрации путем выдержки кислород медленно расходуется на окисление компонентов вина, заметно падает ОВ-потенциал, несколько повышается концентрация альдегидов. При выдержке виноматериалов с добавкой 2 % дрожжей кислород удаляется быстрее, величина падения ОВ-потенциала больше, а свободного ацетальдегида накапливается меньше. Расходование кислорода при выдержке купажа с ферментным концентратом также продолжается длительное время и приводит к заметному снижению ОВ-потенциала. Обработка вина водородом не оказывает влияния на величину ОВ-потенциала.
Термообработка купажа после биологической ассимиляции О2 приводит к дальнейшему падению ОВ-потенциала, увеличению восстановительной способности и накоплению альдегидов. Пастеризация выдержанного в потоке купажа с ликером вызывает образование альдегидов. Нагревание бродильной смеси, обработанной водородом, обусловливает уменьшение концентрации восстанавливающих веществ.
В процессе биологического удаления О2 наблюдается падение активности β-фруктофуранозидазы (БФФ). Если нагревание до 40 °С способствует тепловому автолизу дрожжей и переходу БФФ из дрожжей в вино, то пастеризация при 65 °С инактивирует ферменты вина. Выдержка купажа с ферментным концентратом, обработка его теплом оказывают незначительное влияние на активность БФФ.
Биологическое удаление кислорода сопровождается потреблением азотистых веществ, в частности аминокислот и. аммиака. Последующая термообработка приводит к обогащению бродильной смеси азотистыми веществами. Пастеризация ускоряет реакции меланоидинообразования. При выдержке купажа в потоке без добавления дрожжей содержание большинства аминокислот уменьшается. Количество пролина, глицина, гистидина и аргинина увеличивается. После термообработки накапливаются глютаминовая кислота, пролин, аспарагиновая кислота, а расходуются аланин, валин, аргинин, лейцин и изолейцин. Накопление аминокислот связано с гидролизом белковых веществ, а исчезновение – с процессами дезаминирования.
При выдержке купажа с ферментными концентратами уменьшается содержание аммиака и аминокислот, особенно аланина, валина и аргинина. Обработка купажа только водородом не влияет на содержание азотистых веществ, но в комплексе с нагреванием приводит к снижению содержания некоторых их форм и накоплению меланоидинов. Содержание глицина, лейцина и изолейцина, увеличивающееся в процессе выдержки, уменьшается при термообработке.
С точки зрения создания оптимальных условий для протекания окислительно-восстановительных процессов лучшими способами обескислороживания являются биологический (быстрое воздействие) и выдержка в потоке (медленный эффект). Наибольшее обогащение виноматериала ферментами происходит при нагревании до 40 °С и выдержке при этой температуре в течение 2 сут; в процессе пастеризации это преимущество исчезает. При ассимиляции кислорода вина дрожжами активность сукцинатдегидрогеназы, лактатдегидрогеназы и β-фруктофуранозидазы снижается вследствие адсорбционных явлений. Ряд ферментов (глютамат-, аланин-, малатдегидрогеназы) синтезируется дрожжами и выделяется в вино. Дрожжевые клетки ассимилируют многие аминокислоты. В процессе подбраживания в купаже увеличивается содержание ацетальдегида, н-пропанола, изобутанола, изопентанола, этилацетата, масляной, изовалериановой, капроновой и каприловой кислот. При удалении кислорода путем выдержки концентрация этих компонентов повышается незначительно. Синтезу этилацетата, высших спиртов и жирных кислот способствует наличие в среде сахара. При последующей выдержке на дрожжах в течение 1 мес образовавшиеся спирты и кислоты этерифицируются. В результате тон сивушных масел, усиливающийся после ассимиляции кислорода, исчезает, а накопление сложных эфиров способствует развитию букета выдержанных вин.
В процессе выдержки на дрожжах в купаж переходят ферменты, которые ускоряют биохимические процессы. После деаэрации и выдержки с дрожжами купаж мягче, ароматичнее и значительно лучше, чем при деаэрации без дрожжей.
В связи с тем, что из сахара дрожжи синтезируют нежелательные компоненты, добавлять ликер при биологическом способе не следует. В целях обогащения купажа ферментами, создания условий для протекания реакций этерификации целесообразно биологическую ассимиляцию кислорода проводить сразу после приготовления купажа и далее выдерживать купаж в потоке в течение 1–2 мес.
Качество цимлянских виноматериалов и купажей зависит от способа их деаэрации и последующей обработки теплом в условиях низкого уровня ОВ-потенциала [57]. Биологическую деаэрацию в потоке следует проводить при концентрации дрожжей (в пересчете на сухие) порядка 48 мг/л и удельном расходе вина 0,23 дал/ч на 1 дал рабочего объема колонки. При этом резко снижается содержание растворенного кислорода, восстанавливаются наиболее активные перекиси; интенсивность и качество окраски виноматериалов практически не изменяются; заметно улучшаются их физико-химические свойства (коэффициент сопротивления выделению СО2 К и показатель устойчивости двухсторонних пленок 𝓣), несколько снижается содержание общего и белкового азота (табл. 5).
Таблица 5
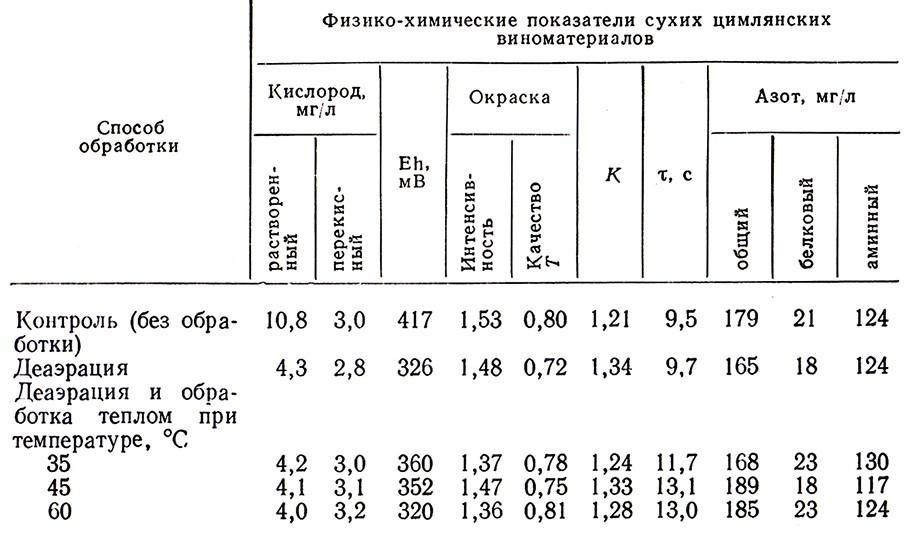
Для деаэрации цимлянских виноматериалов, содержащих сахар, биологический способ неприемлем. Кислород целесообразно удалять из них методами барботирования инертными газами или выдержкой в анаэробных условиях.
Способ выдержки красных крепленых виноматериалов (спирт 12% об., сахар 18%) в анаэробных условиях при температуре 2°С в течение 20–24 сут обеспечивает значительное улучшение технологических свойств. При этом их физико-химические показатели изменяются следующим образом:

Дегустационная оценка виноматериалов, выдержанных до 24 сут в анаэробных условиях, на 0,2 балла выше, чем контрольных виноматериалов (соответственно 8,4 и 8,2 балла).
Обработка резервуарных смесей теплом (при 40 °С в течение 3 сут) или холодом (при –2°С в течение I–2 сут) улучшает органолептические показатели красных игристых вин, сброженных на брют, особенно если в них содержатся 2–4 млн./мл жизнедеятельных клеток дрожжей или 0,25 % ферментного концентрата (табл. 6). Как при нагревании, так и при обработке холодом увеличиваются выход ферментов и других азотистых веществ из дрожжей и обогащение ими вина.
Таблица 6
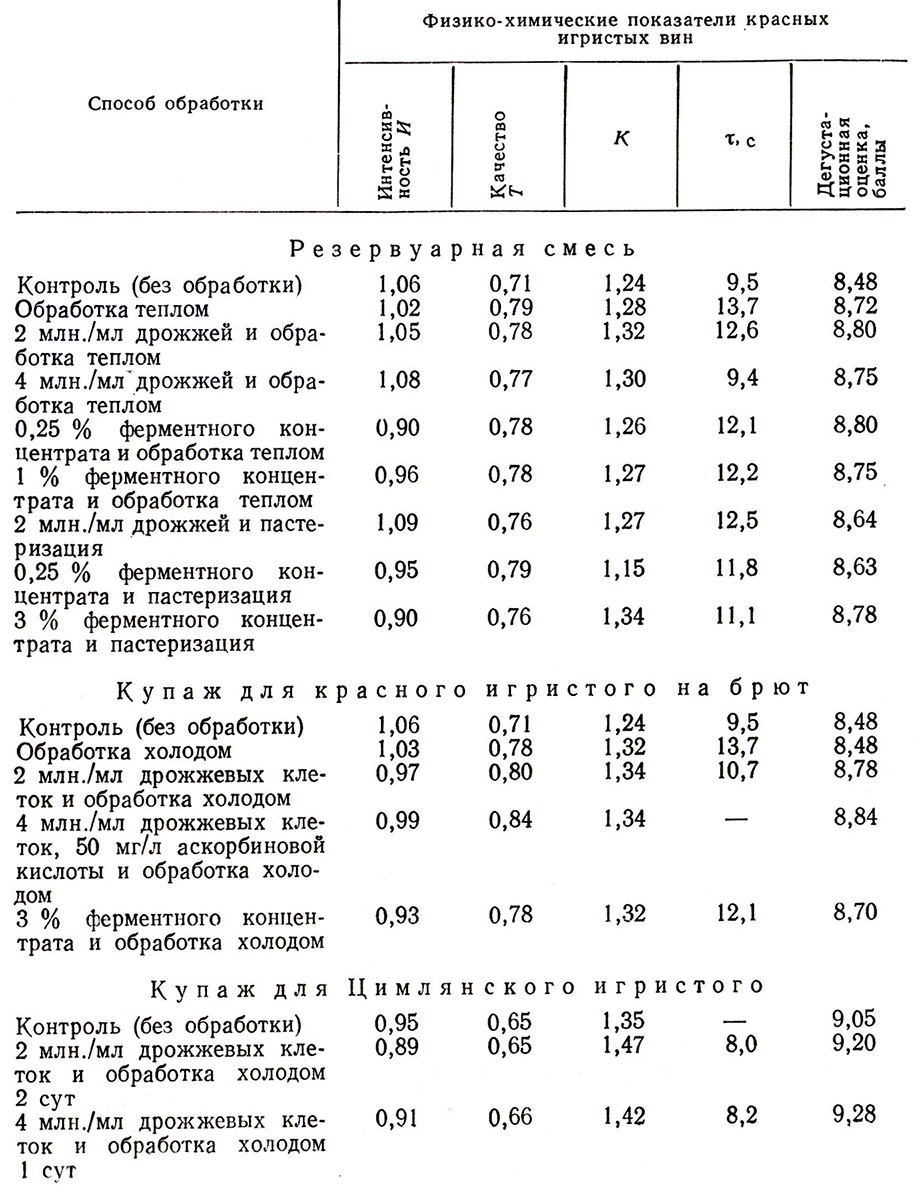
Биохимические процессы, протекающие при шампанизации
Процесс бутылочной шампанизации может быть разделен на 4 периода. В первый период (0–7 сут) дрожжевые клетки интенсивно размножаются, адсорбируя ферменты (рис. 1), потребляя азотистые вещества, ассимилируя кислород, продуцируя диоксид углерода, альдегиды, высшие спирты и другие продукты брожения. В течение следующего периода (7–30 сут) заканчивается вторичное брожение. Потребив весь сахар и накопив значительное количество диоксида углерода, дрожжевые клетки начинают угнетаться и выделять в вино ферменты (табл. 7), азотистые вещества, фосфорные соединения, окислительно-восстановительные системы с низким уровнем ОВ-потенциала. В процессе вторичного брожения, как это следует из теории Г. Г. Агабальянца, образуются химически и физико-химически связанные формы диоксида углерода, которые определяют качественные отличия шампанских вин от искусственно газированных. В третий период, продолжающийся до конца первого года выдержки, в вине протекают интенсивные биохимические превращения (рис. 2). После окончания вторичного брожения концентрация угнетенных дрожжевых клеток увеличивается. Согласно предлагаемому нами механизму процесса автолиза дрожжей при длительной выдержке в клетках дрожжей изменяется клеточный метаболизм, активируются протеолитические ферменты. Протеазы вызывают распад белков и важных ферментов клетки, вследствие чего нарушается координационная связь и регуляция ферментов в дрожжевых клетках и дрожжи начинают автолизоваться. Это наглядно видно на электронно- микроскопических снимках (см. п. 7.5, рис. 13–25).
Таблица 7
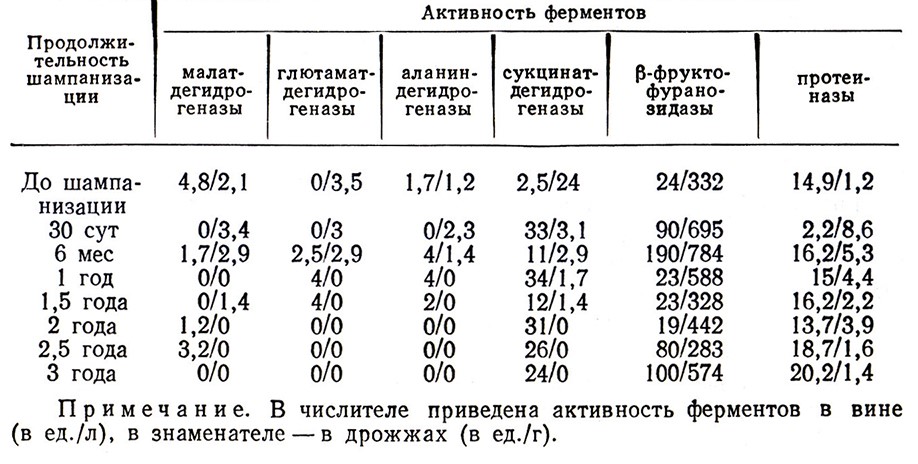
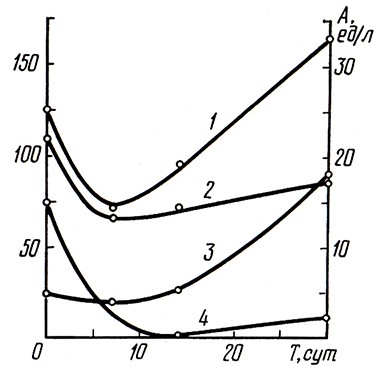
Рис. 1. Изменение активности ферментов вина при вторичном брожении:
1 – сукцинатдегидрогеназа; 2 – лактатдегидрогеназа; 3 – β-фруктофуранозидаза; 4 – протеиназа
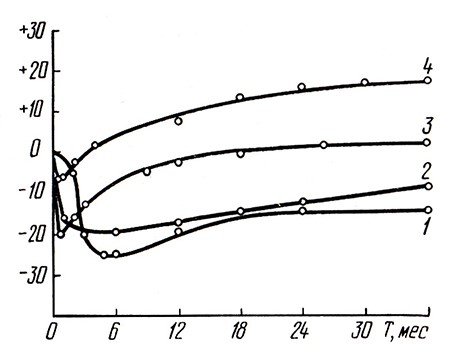
Рис. 2. Биохимические изменения различных показателей (в %) при бутылочной шампанизации:
1 – восстановительная способность; 2 – ОВ-потенциал; 3 – общий азот; 4 – аминный азот
Активирующиеся при автолизе гидролитические ферменты ускоряют распад внутриклеточных макромолекул и переход в вино отдельных компонентов дрожжей: азотистых веществ, главным образом аминокислот, фосфорных соединений, жирных кислот, альдегидов, терпенов, сложных эфиров этилового, изобутилового, изоамилового спиртов. В процессе длительной выдержки в органолептически активные вещества превращаются аминокислоты и другие компоненты вина.
Выделяемые дрожжами ферменты (протеазы, эстеразы, дегидрогеназы) интенсифицируют реакции, обусловливающие формирование качественных особенностей шампанского. Биохимические процессы, а также ферментативные превращения, улучшающие качество шампанского, по всей вероятности, катализируются ферментами дрожжей непосредственно в цитоплазме и отдельных органоидах клетки. Внутриклеточные ферменты служат своеобразными центрами биохимических превращений компонентов шампанизируемого вина. Ферментативные реакции в цитоплазме и органоидах автолизующихся дрожжей, по- видимому, формируют специфические особенности выдержанного шампанского.
Начинающийся после годичной выдержки четвертый период (2–3 года) характеризуется медленно идущими биохимическими превращениями. Относительно низкая величина рН и некоторые компоненты вина вызывают замедление процесса автолиза дрожжей. Вследствие этого большинство дрожжевых клеток после 2–3 лет послетиражной выдержки теряют около 1/3 составных веществ, сильно уменьшаются в размере, но полностью не разлагаются, а становятся похожими на грибы сморчки. Этому способствует невысокая температура выдержки (10–15 °С), при которой внутриклеточные процессы в дрожжевой клетке прекращаются постепенно и некоторые из ее функций сохраняются в течение длительного времени.
В клетках дрожжей, выдержанных 2–3 года, инактивнруются многие оксидоредуктазы, но сохраняют высокую активность гидролитические ферменты. В шампанизируемом вине активны дегидрогеназы, β-фруктофуранозидаза, протеазы (см. табл. 7), которые способствуют протеканию биохимических процессов формирования шампанского.
Вследствие усиливающегося в дрожжевых клетках разрушения субклеточных структур, распада составляющих основу цитоплазмы комплексов белков с липидами, углеводородами и неорганическими солями клетки дрожжей пропитываются липидными гранулами, часть которых переходит в вино. По-видимому, выделяемые дрожжевыми клетками липиды, жирные кислоты, высококипящие сложные эфиры, терпеноиды совместно с вновь образующимися и содержащимися в шампанском букетистыми соединениями обусловливают появление специфических «подсолнечных» тонов выдержанного шампанского. По мнению А. К. Родопуло и сотр., образование этих тонов связано с накоплением этиллинолеата. Накопление важных в органолептическом отношении соединений и протекание биохимических превращений при низком уровне ОВ-потенциала способствуют формированию гармоничного и тонкого букета и вкуса бутылочного шампанского.
Технология резервуарной шампанизации, предложенная А. М- Фроловым-Багреевым, имеет существенные отличия от шампанизации вин в бутылках. По этой технологии процесс шампанизации вина до марки брют протекает в две стадии [3] (рис. 3): на первой (0–12 сут) происходят адсорбция ферментов, ассимиляция аминокислот, синтез альдегидов вследствие размножения дрожжевых клеток; на второй (12–21 сут) в вино выделяются ферменты, азотистые вещества, восстанавливаются альдегиды, что связано с началом автолиза дрожжей.
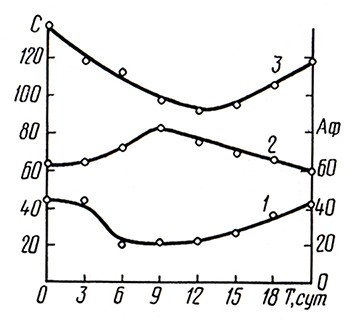
Рис. 3. Биохимические превращения в вине при вторичном брожении в резервуаре:
1 – активность β-фруктофуранозидазы; 2 – альдегиды; 3 – аминокислоты
Таким образом, при периодической шампанизации в резервуаре протекают те же процессы, что и на первых двух стадиях бутылочной шампанизации. Вместе с тем резервуарный периодический способ не имеет 3-й и 4-й стадий, на которых формируется высококачественное бутылочное шампанское.
Совершенствование резервуарного способа привело к созданию способа шампанизации вина в непрерывном потоке [А. с. 122467 (СССР).– Б. И., 1959, № 18; А. с. 236400 (СССР). – Б. И., 1969, № 7]. В процессе шампанизации в потоке происходит сбраживание сахара в условиях постоянного повышенного давления и насыщение вина СО2. Вследствие отсутствия стадии размножения дрожжей при непрерывной шампанизации в отличие от бутылочной нет периода адсорбции ферментов. К концу процесса шампанизации начинающийся автолиз дрожжевых клеток вызывает некоторое увеличение активности протеаз, β-фруктофуранозидазы, эстераз. Отсутствие кислорода и обогащение шампанского восстанавливающими веществами способствуют снижению ОВ-потенциала. Автолитические процессы интенсифицируются в установленных в конце потока резервуарах с насадками (табл. 8). Задерживающиеся в зоне насадок дрожжевые клетки при длительной эксплуатации установки отмирают и обогащают шампанское продуктами автолиза. Протекающие при низком уровне ОВ-потенциала биохимические процессы обусловливают формирование шампанского высокого качества.
Таблица 8

Вместе с тем при шампанизации бродильной смеси с 3,5–5 % сахара наличие остаточного сахара тормозит скорость процесса автолиза дрожжей. При шампанизации вина до марки брют скорость автолитических процессов выше. Обращает на себя внимание тот факт, что при непрерывной шампанизации происходит в основном вторичное брожение и почти отсутствуют наиболее важные с биохимической точки зрения 3-й и 4-й периоды, характерные для бутылочной шампанизации.
С целью воспроизведения условий, существующих при длительной выдержке бутылочного шампанского на дрожжах, нами в 1965 г. предложено сброженное насухо шампанизируемое в потоке вино с дрожжами направлять в резервуары с насадками – автолизеры, в которых задержанные дрожжевые клетки автолизуются и обогащают шампанское важными компонентами цитоплазмы клетки. Указанный прием, внедренный на всех заводах страны, позволил за сравнительно короткий срок повысить качество резервуарного шампанского.
Предложено также проводить шампанизацию в потоке в резервуарах с насадками [91, 96]. Этот метод характеризуется задержкой дрожжевых клеток на насадке, неуклонным увеличением дрожжевой биомассы и концентрации автолизуемых дрожжей, активным сбраживанием сахара при низкой температуре (см. п. 10.3). Заметных колебаний концентраций азотистых веществ не происходит, но улучшаются окислительно-восстановительные показатели: снижается содержание альдегидов, диацетила, повышается восстановительная способность вина, в вине накапливаются ферменты. Шампанизация при повышенной концентрации иммобилизованных дрожжей оказывает существенное влияние на вкус и букет игристых вин.
Биохимические процессы протекают и при обработке шампанизированного вина (см. п. 10.4).
2.3. МЕХАНИЗМ ПРОЦЕССОВ СОЗРЕВАНИЯ
ВИНОМАТЕРИАЛОВ И ИГРИСТЫХ ВИН
Процесс созревания вина включает совокупность сложных физико-химических, химических и биохимических реакций, приводящих к развитию тонов выдержки во вкусе и образованию букета. Исследованию механизма реакций, происходящих при созревании вина, посвятили свои труды Ж. Риберо-Гайон, Э. Пейно, А. М. Фролов-Багреев, М. А. Герасимов, Г. Г. Агабальянц и др. Тем не менее многие вопросы остаются неясными [49].
Согласно современным представлениям при созревании вина протекают следующие реакции: окислительно-восстановительные, карбониламинные, этерификации, полимеризации, поликонденсации, гидролиза и др., которые при длительной выдержке виноматериалов (используемых в купаже), послетиражной и длительной выдержке шампанизируемого вина в бутылках или резервуарах приводят к созреванию шампанского.
Окислительно-восстановительные процессы
Окислительно-восстановительные процессы имеют первостепенное значение в производстве игристых вин. Согласно классификации вин по степени окисленности, предложенной Г. Г. Агабальянцем, белые столовые и шампанские вина относятся к неокисленным винам и не должны содержать свыше 20 мг/л свободного уксусного альдегида. По мнению А. К. Родопуло и сотр., показателем окисленности вин может служить содержание диацетила, причем для игристых вин его концентрация не должна превышать 1 мг/л. Для созревания вин определенного типа необходимо строго определенное количество кислорода, поэтому для белых столовых и шампанских виноматериалов скорость ассимиляции вином кислорода не должна превышать 0,02–0,05 мг О2 на 1 л вина в сутки [6].
Существуют различные точки зрения на роль и механизм окислительно-восстановительных процессов в биологических средах, к которым, в частности, относится и вино.
В биологическом окислении различают два основных направления. Одно рассматривает механизм активирования молекулярного и перекисного кислорода при действии соответственно оксидаз и пероксидаз. Другое направление – это активирование непосредственно водорода органического вещества, причем перенос активированного водорода к активированному кислороду осуществляется в порядке сочетания ряда биокатализаторов, действующих между дегидрогеназами и оксидазами. Представителем первого направления был А. Н. Бах, по мнению которого молекула кислорода непосредственно с водородом не может связываться. Вначале он должен вступить в реакцию с ненасыщенными веществами (–СН = СН–; – С = С–), которые А. Н. Бах назвал «оксигеназами». В результате реакции образуются вещества типа пероксидаз, в которых кислород находится в активном состоянии:
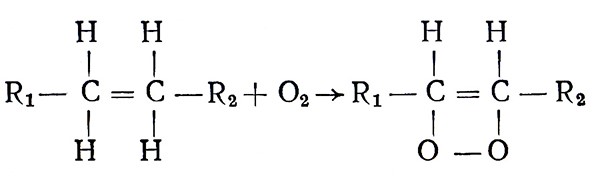
Далее пероксид передает кислород какому-то трудноокисляемому веществу – образуются оксид и вода, оксигеназа регенерируется и может вновь вступать в реакцию:
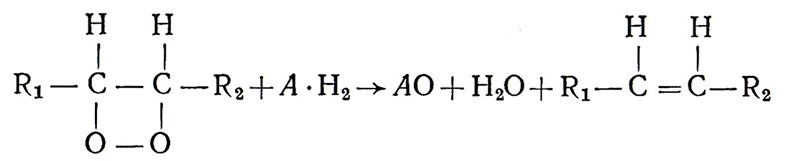
Перекисная теория А. Н. Баха нашла подтверждение после открытия нового класса ферментов – оксигеназ, активирующих молекулярный кислород и катализирующих его внедрение в субстрат окисления.
Согласно теории В. И. Палладина – сторонника второго направления, в окислении органического вещества участвуют особые соединения, так называемые пигменты-хромогены (дыхательные хромогены). По этой концепции процессы окисления органических веществ могут протекать в отсутствие кислорода воздуха. В. И. Палладии впервые показал роль процессов дегидрирования в окислении биологических субстратов и создал оригинальную теорию биологического окисления, которая лежит в основе всех современных представлений о тканевом дыхании.
По современным представлениям биологическое окисление рассматривается как процесс переноса атомов водорода или электронов окисляемого органического вещества на молекулярный или перекисный кислород. В цепи ОВ-реакций водород вначале отщепляется от субстрата и переносится через ряд дегидрогеназ и коферментов – промежуточных переносчиков – на активированный оксидазами кислород, образуя воду.
Кроме биологических катализаторов в винах большую роль играют катализаторы неорганические, в основном ионы меди и железа. Причем комплексно связанные ионы железа являются более сильными катализаторами, чем неорганические соли железа.
В последнее время в химических и биохимических процессах, и в частности окислительно-восстановительных, придают особое значение анион-радикалу О2‾ (часто его называют супер- оксидом), который образуется при одноэлектронном восстановлении молекулярного кислорода и при энзиматическом аэробном метаболизме. Механизм образования анион-радикала О2‾ следующий:
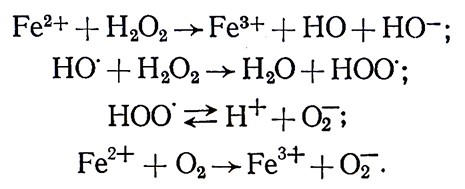
Кроме того, возможно взаимодействие О2‾ непосредственно с Н2О2, в результате чего образуется гидроксильный анион-радикал:
![]()
Это взаимодействие наиболее вероятно в присутствии комплексов железа или меди:
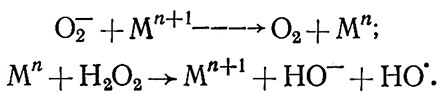
Интенсивность окислительно-восстановительных процессов характеризует редокс-потенциал (ОВ-потенциал), который зависит от соотношения окисленных и восстановленных систем. Часть ОВ-систем являются обратимыми, другие – необратимыми. При доступе кислорода равновесие реакций сдвигается в сторону окисленных форм, при анаэробной выдержке – в сторону восстановленных форм. Наличие дрожжей в вине способствует усилению восстановительных процессов. По мнению ряда ученых, это связано с переходом из дрожжевых клеток восстановителей (глютатиона, цистеина и др.). Важное значение имеют ферменты дрожжей, особенно дегидрогеназы, которые ускоряют реакции восстановления окисленных соединений. Формирование игристых вин высокого качества должно протекать при низком уровне ОВ-потенциала.
При хранении, выдержке и обработке виноматериалов попадание в них кислорода вызывает окислительные процессы. На начальных стадиях производства виноматериалов наличие небольшого количества О2 и невысокая температура выдержки обусловливают небольшую скорость ассимиляции вином кислорода, что благоприятно сказывается на созревании виноматериалов. Многое при этом зависит от химического состава виноматериалов, запаса редуктонов. Для белых и мускатных виноматериалов эти величины минимальны.
Как показали наши исследования, при выдержке виноматериалов и шампанизируемого вина происходят превращения органических кислот и терпеноидов. Эти превращения интенсифицируются при доступе кислорода воздуха и повышенной температуре (40–45 °С). Введение в вино меченых по 14С яблочной и винной кислот показало, что метка 14С яблочной кислоты появилась в щавелево-уксусной, пировиноградной, гликолевой, глиоксиловой, щавелевой кислотах, ацетальдегиде, а также предположительно в фумаровой и двух неидентифицированных кислотах. На основе исследований механизм окисления яблочной кислоты в вине можно представить так:
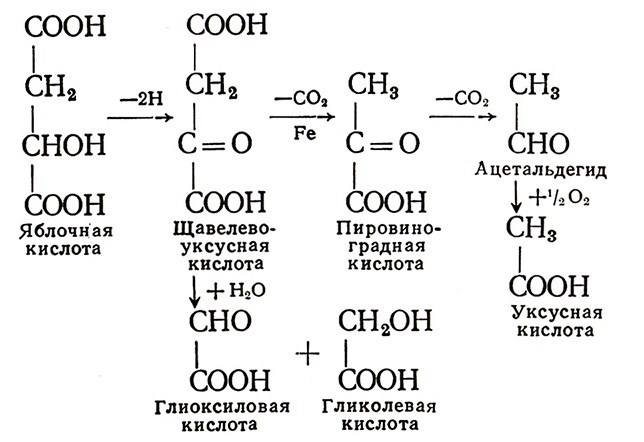
Окисление винной кислоты происходит следующим образом:
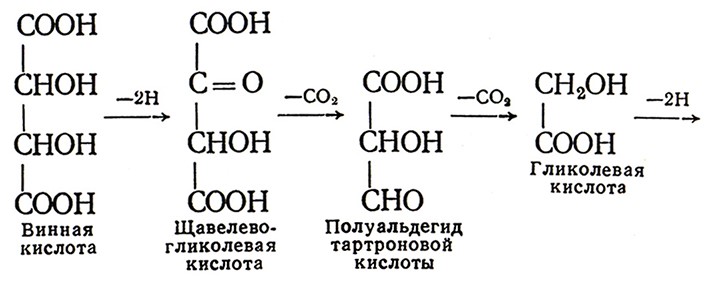
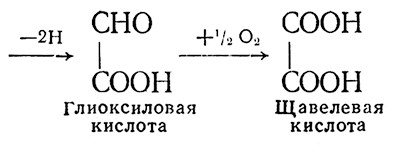
По мнению А. К. Родопуло, образование в шампанских виноматериалах щавелевой кислоты ухудшает их качество.
Превращения терпеновых спиртов – гераниола и линалоола протекает интенсивнее при температуре 40 °С. Наибольшим изменениям они подвергаются в присутствии дрожжей. Состав терпенов, образующихся из указанных спиртов, зависит в наибольшей степени от температуры и длительности выдержки. а-Терпинеол является очень стабильным соединением и в процессе созревания вина изменяется незначительно.
Методом газовой хроматографии установлено, что продуктами превращения линалоола являются гераниол, α-терпинеол, нерол, линалилацетат и геранилацетат, а гераниола – линалоол, α-терпинеол, нерол, линалилацетат, геранилацетат и цитраль. Гераниол и линалоол способны к взаимопревращениям. В процессе тепловой выдержки интенсивно протекают реакции этерификации гераниола и линалоола с образованием соответствующих эфиров уксусной кислоты. Образование α-терпинеола, нерола, цитраля связано, вероятно, с окислением гераниола и линалоола.
В образцах с дрожжами, особенно при температуре 40 °С, в среде накапливается сесквитерпеновый спирт фарнезол. И. А. Егоровым было установлено, что дрожжевые клетки выделяют фарнезол в вино. Имеются сведения, что фарнезол образуется непосредственно в дрожжевой клетке в результате пирофосфорного окисления из гераниола. В процессе выдержки и тепловой обработки фарнезол накапливается в вине в результате автолиза дрожжевой клетки.
Учитывая, что терпены гераниол и линалоол обусловливают аромат мускатов, а при анаэробной выдержке и в присутствии дрожжей их количество уменьшается, особенно при повышенных температурах, нецелесообразно выдерживать виноматериалы для мускатных игристых вин при температуре выше 20–25 °С и осуществлять длительный контакт с дрожжами. Ввиду того что в процессе выдержки на дрожжах образуется фарнезол, выдержка виноматериалов и шампанизированных вин с дрожжами улучшает органолептические качества вина. По современным представлениям терпеноиды обусловливают сортовой аромат виноматериалов. Для его сохранения в мускатных игристых винах, некоторых марках белых и красных игристых вин следует создавать условия, исключающие окисление терпенов, выдержку проводить при пониженной температуре (лучше при 0–5 °С), сокращать срок выдержки шампанизируемого вина, исключать тепловую обработку.
Приведенные данные свидетельствуют о том, что регулирование окислительно-восстановительных реакций при производстве игристых вин имеет крайне важное значение, так как от интенсивности и направленности этих процессов в решающей степени зависят цвет, вкус и аромат вин этого типа.
Реакции этерификации
Сложные эфиры в винах образуются главным образом на стадии брожения в результате синтеза их клетками дрожжей и взаимодействия спиртов с ацильными производными кислот, накапливающимися в вине при биосинтезе высших жирных кислот. Концентрация синтезируемого при брожении эфира может в несколько раз превышать равновесную для соответствующих спирта и кислоты. Преобладание в вине эфиров кислот жирного ряда с четным числом углеродных атомов объясняется механизмом синтеза их кислотных предшественников, согласно которому прогрессивное удлинение ацильных производных происходит за счет конденсации молекулы с ацетил-КоА, т. е. добавлением к молекуле С2. Повышенное содержание эфиров жирных кислот и уксусной кислоты (за счет присутствия в вине при брожении высокой концентрации ацетил-КоА), имеющих приятный фруктовый запах, оказывает непосредственное влияние на аромат молодых вин.
О важной роли эфиров уксусной и высших жирных кислот в формировании качества вин с завершенным брожением свидетельствует высокая корреляционная взаимосвязь между ароматом молодого вина Ркацители и содержанием в нем этих групп эфиров. Коэффициенты парной корреляции между оценкой аромата вина и содержанием в нем изоамилацетата, этилкапроната, гексилацетата, этиллаурата, этилкаприлата, 2-фенилэтилацетата варьируют в пределах от 0,51 до 0,73.
Изучение механизма этерификации кислот, содержащих меченый кислород 18О, показало, что процесс идет путем замены водорода в спиртовом гидроксиле на ацильную группу кислоты. Реакция катализируется ионом водорода или недиссоциированным протоном органической кислоты и является обратимой:
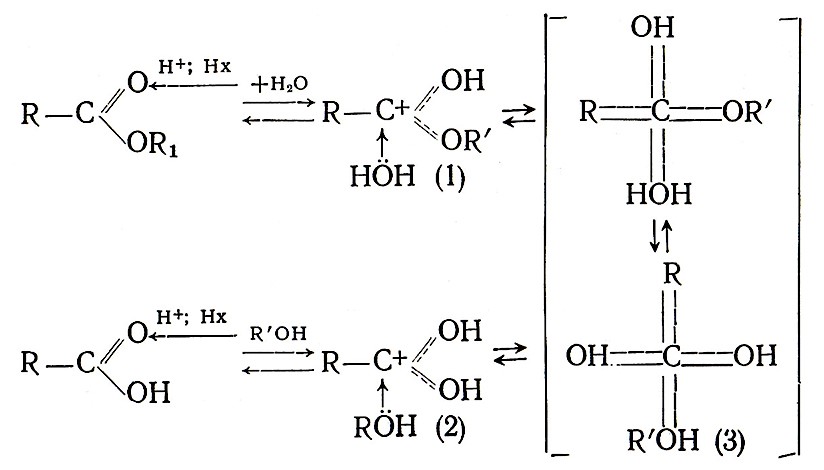
При изучении кинетических закономерностей реакций и изотопного обмена кислорода [123] установлено, что процесс синтеза–гидролиза сложных эфиров протекает через образование неустойчивого промежуточного продукта (3).
Расчет энтропии активации промежуточного комплекса для ряда сложных эфиров уксусной и высших жирных кислот в белых столовых винах показал, что величина ее колеблется от 51 до –81 кал/(моль•К). Отрицательные значения энтропии активации переходного комплекса для синтеза – гидролиза сложных эфиров свидетельствуют о том, что эти реакции в вине протекают по кислотному бимолекулярному механизму через стадию образования тетраэдрического интермедиата (3) за счет присоединения соответственно к протонированному эфиру (1) или кислоте (2) воды или спирта.
Процессы трансформации сложных эфиров особенно интенсифицируются в вине с понижением рН и увеличением температуры выдержки. Наиболее значительные изменения под влиянием указанных факторов претерпевают в вине эфиры уксусной кислоты (этилацетат, изоамилацетат, гексилацетат, 2-фенилэтилацетат), менее существенные – эфиры жирных кислот С6–С14. В процессе выдержки столовых и шампанских виноматериалов отмечается неодинаковая тенденция изменения в них концентраций отдельных эфиров. Выдержка при повышенных температурах (особенно высококислотных вин) приводит к быстрой потере ими сортового аромата. Для сохранения молодого аромата вин хранение их необходимо осуществлять при низких температурах.
Таким образом, главная роль в формировании букета вина в процессе созревания принадлежит обратимым процессам синтеза–гидролиза эфиров легко этерифицируемых кислот, так как концентрации их в вине варьируют в пределах, близких к пороговым величинам сенсорного восприятия. Эфиры многоосновных кислот вследствие присутствия их в винах в концентрациях, значительно ниже пороговых, по-видимому, могут влиять на качество вина главным образом за счет сенсибилизирующего действия, т. е. снижения или усиления в их присутствии восприятия других компонентов.
Карбониламинные реакции
Сахароаминная реакция имеет большое значение в технологии игристых вин. Изучение превращения различных Сахаров и аминокислот, добавленных в вино, показало, что образцы, содержащие фруктозу, при нагревании до 40, 50 и 70 °С очень быстро темнеют [2]. Растворы с фруктозой имеют в несколько раз больший максимум поглощения при 280–290 нм, чем растворы с глюкозой. В связи с тем что оптическая плотность чистого раствора фруктозы примерно равна оптической плотности растворов фруктозы с добавкой аминокислоты, можно считать, что при нагревании модельных сред и вина с фруктозой происходит термический распад фруктозы. В опытах с глюкозой он составляет небольшой процент и основное потемнение связано с накоплением продуктов реакции меланоидинообразования. Из веществ, образующихся при нагревании вина и модельных растворов с аминокислотой и фруктозой, были идентифицированы оксиметилфурфурол, ацетальдегид, пропионовый альдегид, пировиноградная кислота и т. д. Снижение скорости автолиза дрожжей в присутствии сахара, усиление распада фруктозы и неферментативных реакций потемнения вина показывают, что при производстве шампанского целесообразно нагревать купаж без сахара, а ликер вводить после тепловой обработки.
Реакции меланоидинообразования оказывают влияние на фенольный состав, вкус и цвет красного вина [63]. При тепловой обработке красных виноматериалов содержание аминокислот снижается в большинстве случаев до следов, что свидетельствует об их полном распаде. Виноматериалы, обработанные теплом, отличаются своеобразным ароматом различной интенсивности, что может служить косвенным свидетельством реакций меланоидинообразования, сопровождающихся распадом аминокислот с образованием соединений, ответственных за появление тех или иных запахов. При этом наибольшие превращения претерпевают мономерные и малоконденсированные формы фенольных соединений – лейкоантоцианы, антоцианы и фенольные вещества, дающие окрашивание с ванилином.
Действие аминокислот на антоцианы и другие полифенолы активнее при жесткой термической обработке [70]. В таких условиях образуются многие активные побочные продукты (перекиси, альдегиды и пр.), которые обусловливают ускорение процесса разрушения фенольных веществ.
Карбониламинные реакции, называемые также сахароаминными и реакциями неферментативного потемнения или меланоидинообразования, – это сложные реакции, протекающие через несколько стадий по кислотно-основному и радикально-цепному механизму [7, 102]. На первых стадиях происходит дегидратация Сахаров и образующиеся продукты циклизуются, давая гетеро- и карбоциклические кислородсодержащие соединения, или конденсируются с азотистыми веществами, образуя гетероциклические азотсодержащие соединения (см. схему).
При тепловой обработке вина образуется 7 кислородсодержащих соединений: фурфурол, 5-метилфурфурол, 5-этоксиметилфурфурол, 2-ацетилфуран, фурфурилметилкетон, 3,5-диокси-2-метил-4Н-пиран-4-он, 4-окси-2-метокси-5-метил-3 (2Н) -фуранон, которые обусловливают карамельные тона, и 4 производных пиразина: метилпиразин, 2-5-диметилпиразин, 2-этил-З-метилпиразин, 2,5-диэтил-З-метилпиразин, которые обладают тонами хлебной корочки и «жжеными» тонами. Перечисленные компоненты отличаются низкими порогами аромата и совместно с другими продуктами распада Сахаров и аминокислот формируют в вине специфические тона тепловой обработки. Промежуточные продукты: карбонильные, карбоциклические и гетероциклические соединения – при нагревании участвуют в реакциях полимеризации и поликонденсации, которые в конечном итоге приводят к образованию темноокрашенных веществ – меланоидинов.
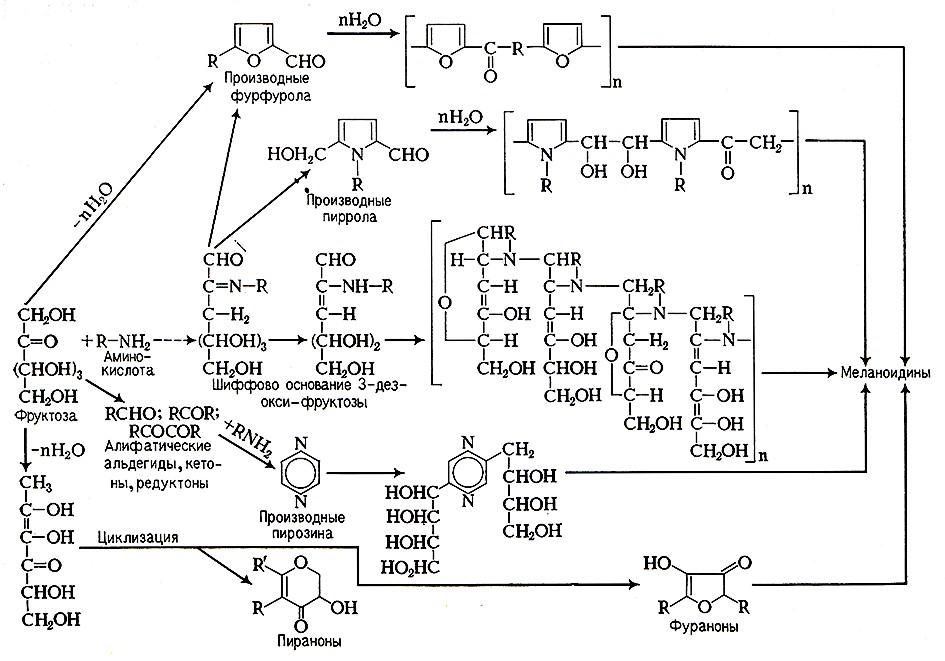
Схема образования меланоидинов в вине при тепловой обработке
На интенсивность и направленность карбониламинных реакций оказывают влияние состав вина, кислородный режим, наличие катализаторов. Органические кислоты (винная, фумаровая, пировиноградная) ускоряют превращения углеводов. Аскорбиновая кислота, дегидрируясь и взаимодействуя с аминокислотами, интенсифицирует потемнение вина. Экстрактивные вещества (особенно полифенолы и азотистые вещества), ионы железа, кислород, перекиси ускоряют формирование тонов тепловой обработки. Введение SО2 в дозе 20–80 мг/л ингибирует эти процессы. Особенно ускоряется меланоидинообразование при повышении температуры до 50–60 °С. Карбониламинные реакции проходят и при длительной выдержке вина при низких температурах (10–20 °С), особенно в присутствии дрожжей, что имеет место при послетиражной выдержке. И хотя структура продуктов реакций в этих условиях иная, их влияние на органолептические качества игристых вин нельзя исключать.
2.4. СПОСОБЫ РЕГУЛИРОВАНИЯ БИОХИМИЧЕСКИХ ПРОЦЕССОВ
ФОРМИРОВАНИЯ ИГРИСТЫХ ВИН
Направленность биохимических процессов, влияющих на формирование игристых вин, можно изменять путем регулирования процессов при получении и обработке виноматериалов, направленного изменения биохимических особенностей дрожжевых клеток и селекции штаммов с ценными для производства признаками, регулирования биохимических процессов при шампанизации.
Регулирование процессов при получении виноматериалов включает выбор определенных сортов винограда, условий их культивирования, способов сбора и переработки в зависимости от типа игристых вин. Способы мацерации винограда, извлечения сока, ферментации сусла существенно сказываются на составе сусла. Регулирование условий брожения, размножения дрожжей, изменение температуры, применение определенных штаммов дает возможность направленно менять состав виноматериалов. По окончании брожения выдержка на дрожжах, переливки, обработка холодом и теплом, выдержка в анаэробных условиях, барботирование газами, деаэрация биологическим методом и другие технологические приемы способны устранить определенные недостатки виноматериалов, ускорить их созревание, обогатить определенными веществами, улучшить качество.
Биохимические особенности дрожжевых клеток можно изменять путем подбора определенных штаммов, воздействием на генный аппарат клетки, регулированием условий культивирования. Селекция штаммов с денными для производства биохимическими признаками, как показывают результаты многочисленных отечественных и зарубежных исследований последних лет, – это весьма перспективное направление существенного улучшения качества игристых вин.
Регулирование процессов при шампанизации – достаточно сложная задача. Среди факторов, оказывающих влияние на ход процессов при бутылочном способе, следует отметить продолжительность и температуру выдержки, способ укупорки, дозирование определенных веществ в тиражную смесь, изменение числа перекладок, интенсивности перемешивания. При резервуарной шампанизации помимо этого важное значение имеют способ сбраживания, концентрация вводимых и задерживаемых дрожжей, улучшение условий контакта шампанизируемого вина с дрожжами, исключение явлений дешампанизации и др.
Более подробно способы регулирования биохимических процессов описаны при рассмотрении специальных вопросов.
В заключение следует отметить, что биохимическая теория процессов формирования игристых вин позволяет определить направления научных исследований в данной области. На перспективу намечено три таких направления:
1 – установление оптимального состава виноматериалов для различных типов игристых вин, разработка объективных критериев для оценки их качества и пригодности использования для шампанизации, совершенствование технологических приемов их производства и обработки;
2 – определение важнейших биохимических признаков дрожжей, селекция перспективных штаммов и разработка технологии их культивирования. Из биологических признаков особый интерес представляют такие, как возможность биосинтеза ценных для букета и вкуса шампанского компонентов, образование связанных форм диоксида углерода, высокая восстановительная способность, ферментативная активность. При селекции дрожжей перспективно использование методов генной инженерии. Подвергнутся пересмотру способы культивирования и использования дрожжей;
3 – регулирование биохимических процессов при шампанизации. Предстоит уточнить сроки и условия шампанизации, состав и количество вводимых добавок и дрожжей, условия контакта шампанизируемого вина с дрожжами с целью интенсификации биохимических процессов при шампанизации, оптимальные режимы выдержки и обработки шампанизированного вина; смоделировать в условиях резервуарной непрерывной шампанизации процессы, протекающие при шампанизации в бутылках, разрешить возникающие при этом технические проблемы.
