Иммобилизованные ферменты и клетки – это всегда гранула того или иного размера (или плоский слой иммобилизованного агента).
Размер гранулы определяется ее радиусом R.
Субстрат (в том числе растворенный кислород) проходит внутрь гранулы через поры или через гель и одновременно на своем пути потребляется клетками.
Если клетки свободно витают в питательной среде, можно принять, что скорость реакции Кзависит от концентрации субстрата S или кислорода С по уравнению Михаэлиса–Ментен: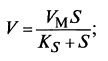 (12·30)
(12·30) ![]() (12·31)
(12·31)
Но при наличии гранулы концентрация субстрата в грануле разная в зависимости от расстояния до центра гранулы (рис. 12.12).
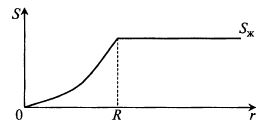
Рис. 12.12. Профиль изменения концентрации субстрата
по глубине гранулы иммобилизованного фермента:
Sж– концентрация субстрата в жидкости;
r – текущее расстояние от центра гранулы;
R – радиус гранулы
В середине гранулы может образовываться зона, куда субстрат не доходит, т. е. S = 0. В жидкости S = Sж на поверхности гранулы S = SR = Sж.
В зависимости от размеров гранулы, скорости потребления субстрата, концентрации фермента (или биомассы) в грануле профиль субстрата по ее глубине будет разный (рис. 12.13).
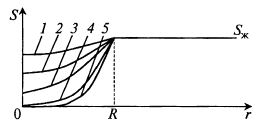
Рис. 12.13. Варианты профиля концентрации субстрата
по глубине гранулы в зависимости от концентрации фермента в грануле:
1–5 – различные концентрации фермента в грануле в порядке возрастания
В центре гранулы может быть, а может и не быть зона голодания. Ясно, что и общая средняя скорость потребления субстрата гранулой при этом будет разная.
Можно найти теоретическое решение для профиля распределения концентрации субстрата по грануле и для средней скорости потребления субстрата гранулой V̅.
Эти расчеты приводят к новой зависимости:![]() (12.32)
(12.32)
Здесь вместо Vстоит средняя скорость V̅, вместо текущей концентрации S около каждой клетки по всему объему гранулы стоит концентрация на ее поверхности, принятая равной концентрации в жидкости Sж.
Показано, что максимальная скорость потребления субстрата Vm такая же, как для одиночной клетки.
А вот константа KS, по смыслу аналогичная субстратной константе Михаэлиса, которую часто называют «кажущейся» константой, отличается от величины KS «истинной», т. е. от субстратной константы, определяемой закономерностью V(S), которой подчиняется каждая отдельная клетка в грануле. Связь K’S(KS) выражается соотношением
K’S = KS( 1+0,0175 Da). (12.33)
Здесь Da – критерий Дамкелера для гранулы радиуса R, определяемый следующим выражением:![]() (12·34)
(12·34)
где Vm – максимальная скорость потребления субстрата на единицу объема; R – радиус гранулы; KS – «истинное» значение константы Михаэлиса; DS – коэффициент диффузии субстрата внутри гранулы.
Физически критерий Дамкелера представляет собой соотношение максимально возможных скоростей потребления субстрата и его диффузии внутрь гранулы.
Если речь идет об иммобилизованных клетках микроорганизмов, то вместо Vm можно записать:![]() (12.35)
(12.35)
где Ʋm – удельная скорость потребления субстрата на единицу массы клеток; X – концентрация клеток в грануле на единицу объема.
Мы здесь использовали обозначения К и υ для общей и удельной скоростей, принятые в ферментативной кинетике. Поскольку иммобилизованные клетки подчиняются уже закономерностям микробной кинетики, логично перейти к обозначениям Q и q. В результате получаем: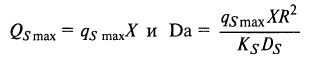 (12.36)
(12.36)
или для кислорода![]() (12.37)
(12.37)
Обычно величина второго члена в скобках уравнения (12.33) значительно больше 1. Поэтому кажущаяся константа Михаэлиса возрастает при увеличении размера гранулы R, или максимальной потребления субстрата, или концентрации клеток микроорганизмов в грануле (плотности заполнения гранулы клетками).
Функция Q̅ S(Sж) (зависимость среднего по грануле потребления субстрата от концентрации субстрата в жидкости на поверхности гранулы) для разных гранул будет иметь разный вид в зависимости от величины критерия Дамкелера (рис. 12.14).

Рис. 12.14. Искажение зависимости Qs(Sж)
при изменении критерия Дамкелера для гранулы:
1 – неискаженная кинетическая зависимость для свободных клеток;
2–5 – зависимости для гранул с различными (возрастающими)
значениями критерия Дамкелера
Были проведены расчеты для разных субстратов. И для одного и того же процесса оказалось, например, что для медленно потребляемых субстратов (глюкоза) кажущееся значение K’S мало отличается от KS, а для кислорода, например при биосинтезе пенициллина, отличие K’S от KS – в тысячи раз.
А раз отличается K’S, то и критическая концентрация кислорода, которую нужно поддерживать в среде, оказывается значительно больше, чем для свободных клеток. В табл. 12.1 представлены сравнительные данные по значениям qO2max, X и qO2max, формирующим величину критерия Дамкелера, для разных культур.
Таблица 12.1. Сравнительные данные по дыханию клеток
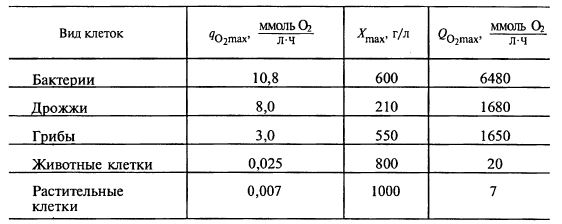
Сравнительные данные по дыханию клеток показывают, что наиболее критична ситуация с иммобилизацией бактерий, в то время как животные и растительные клетки самой природой предназначено иммобилизовать – они ведь обычно практически и существуют в виде «тканей», т. е. как раз прикрепленных клеток. Наоборот, они меньше привыкли к «свободному витанию».
Анализ зависимости показывает, что диффузия кислорода лучше вдет в более мелких гранулах. Крупные, которые, казалось бы, лучше отделяются, имеют зоны кислородного голодания, а также больше подвержены механическому воздействию мешалки и могут разрушаться. Компромисс достигается при размере гранул 200–500 мкм, которые не вызывают проблем.
