К плесневым грибам (иногда их называют плесенями) относят микроскопические грибы-сапрофиты, питающиеся органическими веществами мертвых растительных остатков и образующие на их поверхности налеты разного цвета.
В виноделии нашли применение ферментные препараты, обладающие пектопротеолитическим, целлюлолитическим и гемицеллюлазным действием, готовящиеся из плесневых грибов рода Aspergillus, которые ускоряют отделение сусла от мезги и осветление сусла, увеличивают его выход, интенсифицируют биохимические процессы, протекающие при созревании вин [52].
В вине плесневые грибы не размножаются, но вино, налитое в тару с запахом плесени или укупоренное заплесневевшими пробками, приобретает плесневелый тон и становится испорченным. Обычно винодел принимает меры, предупреждающие появление плесени на различных объектах винодельческого производства, но мало знает о распространенных на винограде и винзаводах видах плесневых грибов и об особенностях их биологии.
КЛАССИФИКАЦИЯ ПЛЕСНЕВЫХ ГРИБОВ
В основу классификации плесневых грибов положены способы их размножения, строение мицелия и органов размножения. Большинство плесневых грибов размножается бесполым и половым путем. Их называют совершенными. Плесневые грибы, размножающиеся только бесполым путем, относят к несовершенным. Грибы, образующие несептированный мицелий, называют низшими, а образующие многоклеточный (септированный) мицелий – высшими грибами.
Плесневые грибы можно обнаружить среди представителей четырех больших классов грибов:
- фикомицетов (Phycomycetes), имеющих несептированный мицелий;
- аскомицетов (Ascomycetes), которые еще называют сумчатыми грибами, образующими септированный мицелий и эндогенные половые споры в сумках (асках);
- базидиомицетов (Basidiomycetes), образующих экзогенные половые споры на особых структурах базидиях;
- несовершенных грибов (Fungi imperfecti), которые обладают многоклеточным мицелием, но не образуют половых спор. Несовершенные грибы называют также дейтеромицетами (Deutero myces) [216, 223].
В группу несовершенных грибов объединены все грибные формы, которые по характеру своего многоклеточного мицелия должны быть отнесены к высшим грибам (сумчатым или базидиальным), но у которых не обнаружено полового спороношения, вследствие чего они и названы несовершенными.
Род Мисог
Плесневые грибы рода Mucor относятся к низшим грибам, к классу Phycomycetes. Мицелий у них обильно ветвящийся, но несептированный. Кверху от него отходят гифы-спорангиеносцы, несущие шарообразные спорангии, видимые простым глазом в виде головок, внутри которых находятся споры (рис. 41, а). Стенки спорангиев растрескиваются, освобождая споры, которые, попав в благоприятные условия, вновь прорастают в мицелий. Обычно у мукоровых грибов мицелий бывает вначале белый, позднее сероватый. Спорангии вначале желтоватые, позже темнеют, приобретая черную окраску.
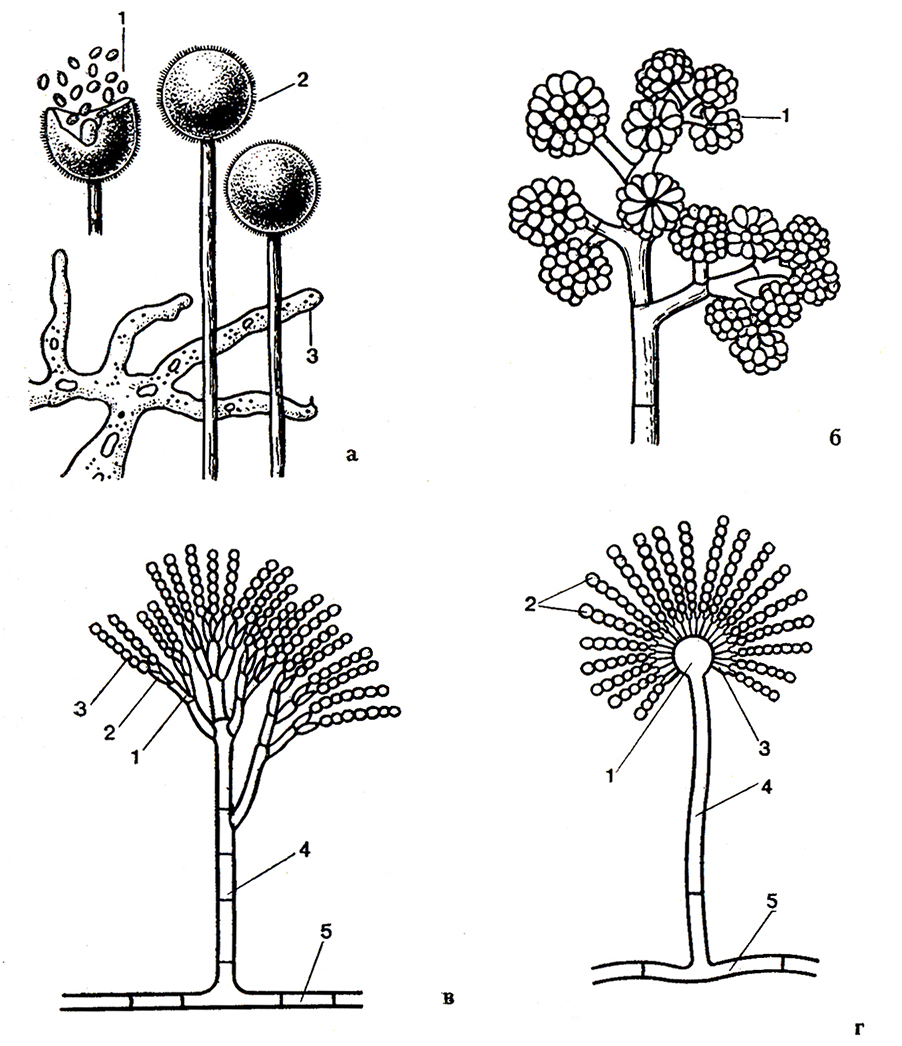
Рис. 41. Споры бесполого способа размножения:
а – Mucor: 1 – споры; 2 – спорангий; 3 – мицелий;
б – BotrytJs: 1 –конидии;
в – Penicillium: 1 – метулы; 2 – стеригмы; 3 – конидии; 4 – конидиофор; 5 – вегетативная гифа;
r – Aspergillus: 1 – булавовидное расширение конидиеносца;
2 – конидии; 3 – стеригмы; 4 – конидиофор; 5 – вегетативная гифа.
Бесполое размножение мукоровых грибов может происходить также оидиями – дрожжеподобными образованиями, называемыми мукоровыми дрожжами (рис. 42, в).
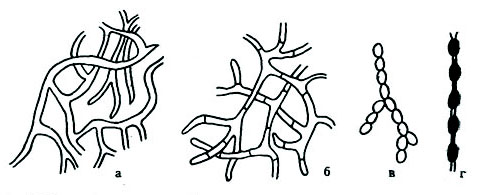
Рис. 42. Мицелий плесневых грибов и его видоизменения:
а — одноклеточный мицелий; б — многоклеточный мицелий; в — оидии; г — хламидо-споры
Они образуются путем распада гифов на отдельные короткие членики при попадании мицелия в жидкую питательную среду с недостаточным притоком воздуха. Мукоровые дрожжи обладают слабой сбраживающей способностью. При свободном доступе воздуха к среде они снова прорастают в мицелий.
Род Aspergillus
Грибы рода Aspergillus относятся к высшим грибам, так как имеют многоклеточный мицелий. Конидиеносцы плесневых грибов этого рода несептированы, на верхнем конце их находятся шаровидные или булавовидные вздутия. От вздутий отходят во все стороны по одному или по два ряда стеригм к цепочки конидий (см. рис. 41, г).
Общая картина конидиеносца с конидиями напоминает головку лейки, из которой струями выходят водяные брызги. Это сходство и привело к названию леечная плесень (по-латински аспергере – поливать, опрыскивать).
В зависимости от видовой принадлежности аспергиллов конидии их бывают окрашены в зеленый, желтовато-зеленый, бурый, темно-коричневый или черный цвет. Благодаря огромному их количеству, расположенному на приземистых конидиеносцах, весь налет плесени кажется окрашенным в цвет конидий.
Аспергиллы относятся к классу Ascomycetes. Половое размножение у них происходит спорами, образующимися в округлых сумках (асках), расположенных в замкнутых плодовых телах (клейстокарпиях), с диаметром от 80–350 мкм.
Род Penicillium
Грибы рода Peniciliium относятся к высшим грибам к классу Ascomycetes и являются наиболее распространенными среди плесневых грибов.
У пенициллов конидиеносцы большей частью имеют поперечные перегородки. В верхней части конидиеносцы разветвлены и образуют характерно построенную кисточку (отсюда и название пенициллиум, по-латински – кисть), несущую на конечных разветвлениях цепочки конидий (см. рис. 41, в). Различное строение кисточки у разных видов пенициллов положено в основу их систематики. Конидиеносцы часто прилегают плотно один к другому или склеены в коремии (от греч. корос – веник). Конидии у большинства видов пенициллов зеленого цвета, характерной для видов формы. Сверху они покрыты слоем воска, вследствие чего трудно смачиваются водой. Конидии могут размножаться даже после многолетнего пребывания в сухом состоянии. Они быстро прорастают в кислых средах. В нейтральных или щелочных средах прорастание их замедляется или не происходит.
Споры полового размножения образуются в асках (сумках), расположенных в плодовых телах (клейстокарпиях). Клейстокарпии обнаружены не у всех видов пенициллов, но по типичному строению конидиеносцев их относят к одному роду.
Род Sphaerulina
Грибы рода Sphaerulina относятся к высшим грибам, к классу Ascomycetes. На листьях, побегах, ягодах винограда, а также на бочках и стенах подвалов иногда можно наблюдать черный сажистый налет. Его образует гриб черни Sphaerulina intermixta, который раньше был известен под названиями Dematium pullulans и Pullularia pullulans.
Черный налет, по виду напоминающий сажу, состоит из огромного количества темноокрашенных устойчивых к неблагоприятным условиям конидий. При благоприятных условиях конидии прорастают и из них образуются белые гифы, в протоплазме которых много крупных вакуолей. От гиф, сплетающихся в мицелий, отчленяются дрожжеподобные почкующиеся клетки (бластоконидии) (рис. 43).
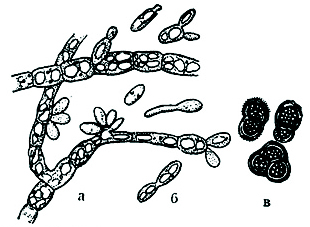
Рис. 43. Гифы, бластоконидии и устойчивые конидии Sph. intermixta:
а — гифы с бластоконидиями; б — дрож- жеподобные бластоконидии; в — устойчивые конидии.
Род Botrytis
Впервые возбудитель серой гнили винограда был описан в 1801 г. X. Г. Петерсоном, давшим ему название Botrytis cinerea (по-гречески ботритис – гроздь, цинереа – серый как зола). Вначале он был известен только в конидиальной форме и отнесен к классу несовершенных грибов. Позже Антон де Бари, подробно изучавший его биологию, обнаружил форму полового развития и включил его в род Sclerotinia, дав видовое название fuckeliana. Но старое название, относившееся к обычно наблюдаемой конидиальной стадии этого гриба, более распространено и им часто пользуются в научной литературе.
Мицелий Botrytis многоклеточный, конидиеносцы нитевидные, древовидно-разветвленные. На концах их образуются малозаметные вздутия, от которых отходят тонкие стеригмы с сероватыми конидиями, расположенными в виде гроздей (см. рис. 41,6). При исследовании микрофлоры виноградного сусла конидии ботритиса легко отличить от дрожжей по более крупным размерам (10–14 X 7–9 мкм).
Перезимовывает ботритис в форме склероциев – плотных мицелиальных образований размером 2–4X1–2,5 мм, окрашенных в темный цвет. Склероции образуются осенью на опавших листьях, на различных частях виноградного куста (рис. 45).
При благоприятных условиях прорастание склероциев происходит либо в конидиальную, либо в сумчатую стадию (половое размножение).
МОРФОЛОГИЯ ПЛЕСНЕВЫХ ГРИБОВ
Строение
Плесневые грибы при размножении на различных питательных субстратах образуют бархатистые, порошкообразные, войлочные, паутинообразные, моховидные налеты зеленого, белого, черного, желтого и других цветов различных оттенков.
Налет, образуемый плесневыми грибами, состоит из массы ветвящихся тонких нитей, называемых гифами. Скопление гифов образует мицелий. Гифы представляют собой нитевидные тяжи протоплазмы, сверху покрытые оболочкой (рис. 43).
В состав клеточной оболочки высших грибов (сумчатых и базидиальных) входит хитин, у некоторых плесневых грибов оболочка состоит из целлюлозы или соединений, близких к ней.
В клетках гифов грибов содержится по одному или несколько ядер. К особенностям ядерного аппарата грибов относится также наличие дикарионов, спаренных ядер в клетке после слияния их протоплазмы. Так же, как и в клетках дрожжей, имеются митохондрии, выполняющие функцию генераторов энергии, рибосомы, принимающие участие в синтезе белка, вакуоли, заполненные клеточным соком и содержащие гранулы полифосфатов (волютина). Основным запасным компонентом клеток плесневых грибов является гликоген, распределенный в виде мелких гранул по всей цитоплазме. Липиды и жировые вещества находятся в клетках в виде капелек, называемых липосомами. В старых клетках гифов грибов содержится много пигментов.
Многие виды плесневых грибов образуют гифы на поверхности и одновременно в толще используемого ими субстрата. Часть гифов служит для прикрепления к субстрату и потребления питательных веществ и называется субстратным мицелием, а часть образует воздушный мицелий, в котором формируются плодоносящие гифы со спорами. Гифы могут быть короткими или длинными. Толщина их колеблется от 1 до 15 мкм, длина – от 2065 до 50 мкм и более. Гифы могут не иметь поперечных перегородок (септ) и образовывать одноклеточный, несептированный мицелий или многоклеточный (септированный) (см. рис. 42, а, б).
У плесневых грибов известны такие видоизменения мицелиального роста, как оидии, хламидоспоры, мицелиальные тяжи (см. рис. 42, в, г) и склероции (рис. 44, а). Так, например, у гриба Mucor racemosus в сахаристых жидкостях грибница может распадаться на отдельные клетки, называемые оидиями, которые затем могут снова образовать мицелий.
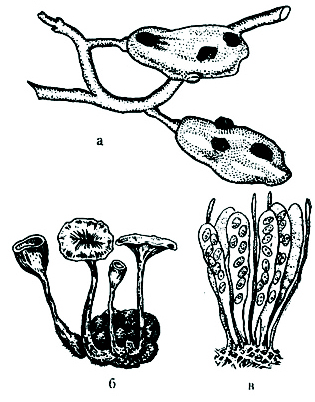
Рис. 44. Склероции, апотеции и сумки со спорами В. cinerea:
а — склероции иа ягодах; б — склероции с тецнями; в —сумки со спорами.
При формировании хламидоспор обособляются участки мицелия, которые покрываются толстой оболочкой, заполняются запасными питательными веществами и в некоторой степени обезвоживаются. В таком виде они могут оставаться длительное время в состоянии покоя, выдерживать высушивание и другие неблагоприятные воздействия.
Грибы родов Aspergillus, Botrytis могут при неблагоприятных условиях (высыхание, понижение температуры) образовывать склероции. Это округлые тела плотной консистенции, образующиеся в результате тесного переплетения гифов, богатые запасными питательными веществами.
На одном склероции вырастает один или несколько апотециев – плодовых тел (рис. 44, б). В апотеции правильными рядами располагаются сумки со спорами среди парафиз – бесплодных нитей, предохраняющих сумки от повреждения и высыхания (рис. 44, в). В нормальной развитой сумке формируется 8 аскоспор, которые, освободившись из асков, прорастают, образуя мицелий и конидиеносцы.
В годы с повышенной влажностью воздуха Botrytis cinerea быстро размножается на ягодах винограда в виде серого налета войлочной консистенции (рис. 45), который приводит к значительным потерям урожая и понижению его качества. Мицелий так называемого домового гриба Merulius domesticus, проникающего внутрь древесины и разрушающего ее, образует мицелиальные тяжи сложного строения. Длина их достигает иногда нескольких метров. По ним передается вода, питательные вещества, и поэтому гриб может распространяться на большие расстояния по поверхности, не способной поддерживать его существование. Встретив древесину и благоприятные условия существования, гриб снова образует мицелий, а затем мицелиальные тяжи.

Рис. 45. Виноградная гроздь, пораженная В. cinerea.
Слева — склероции на побеге.
Способы размножения плесневых грибов
В связи с тем что плесневые грибы можно обнаружить среди представителей четырех больших классов грибов, процесс размножения у них очень разнообразен. Типичный половой процесс – это слияние двух клеток-гамет и их ядер с последующим редукционным делением.
В цикле развития многих плесневых грибов гаплоидная и диплоидная фазы последовательно чередуются: от копуляции до редукции – диплоидная, от редукции до новой копуляции – гаплоидная.
Для грибов характерно явление гетерокариозиса – наличия в соматической клетке гетерокарионов (генетически неоднородных ядер), возникающих при мутации гомокарионов или при анастомозах (слиянии) гифов различных штаммов и переходе таким путем ядер из одной клетки в другую. Гетерокариозис обусловливает большую изменчивость грибов.
Распространен у плесневых грибов и парасексуальный процесс, во время которого происходит слияние двух гаплоидных ядер в гетерокариотических клетках, после чего происходит не редукционное, а митотическое деление. В ходе этого деления происходит кроссинговер (обмен гомологичными участками между гомологичными хромосомами) и возникают генетические рекомбинанты, т. е. особи, несущие новые комбинации родительских генов [23].
Органы полового размножения у плесневых грибов очень разнообразны. У плесневых грибов, принадлежащих к классу фикомицетов, гаметы обычно образуются на разных особях. Половые различия у этих грибов внешне не проявляются; их пол обычно обозначают знаками «плюс» ( + ) и «минус» (–). В результате слияния гамет (оплодотворения, которое у этих грибов называют изогамией) образуется зигоспора (рис. 46, а). При ее прорастании происходит мейотическое деление диплоидного ядра и образуются ядра с гаплоидным числом хромосом. Весь дальнейший цикл развития гриба – гаплоидная фаза.
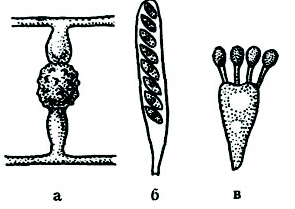
Рис. 46. Споры полового способа размножения:
а – зигоспора фикомицета; б – сумка с 8 спорам» аскомицета; в – 4 базидноспоры базндиомицета.
Половой процесс у типичных аскомицетов характеризуется оплодотворением дифференцированной женской половой клетки (архикарпа) мужской половой клеткой (антеридием). При этом сначала сливаются цитоплазмы (цитогамия), а ядра, сближаясь друг с другом, но не сливаясь, образуют дикарион. Из оплодотворенной клетки вырастают аскогенные гифы, в которые переходят и дикарионы.
Ядра несколько раз раздельно делятся и сливаются в развивающейся сумке, затем сразу же происходит редукционное деление. При этом образуется 4–8 спор с гаплоидными ядрами в сумке, называемой аском (рис. 46, б).
У некоторых сумчатых грибов аски со спорами образуются непосредственно на мицелии, у других они помещаются в плодовых телах разной формы и строения (рис. 47). В цикле развития аскомицетов преобладает гаплоидная фаза, весь мицелий и плодовые тела имеют гаплоидный набор хромосом, кроме аскогенных гиф.
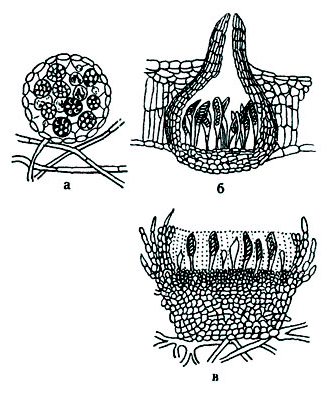
Рис. 47. Формы плодовых тел аскомицетов:
а – клейстокарпнй; б – перитеций; в - апотеций.
У дереворазрушающих плесневых грибов, принадлежащих к базидиальным грибам, неизвестны специальные половые клетки. Диплоидный мицелий образуется в результате анастомозов и слияния клеток одного или двух мицелиев. Через перетяжки сливающихся клеток ядро переходит из одной клетки в другую, получаются клетки с дикарионами, разрастающиеся в мощную систему мицелия. На нем образуются базидии, в которых ядра дикарионов сливаются, копуляционное ядро сразу вновь редукционно делится и образуемые на базидиях базидио-споры имеют гаплоидные ядра (см. рис. 46, в). Базидиоспоры вновь прорастают в первичный гаплоидный мицелий. В цикле развития преобладает диплоидная фаза.
Все способы формирования новой особи без полового процесса, т. е. слияния ядер, можно считать бесполыми, или вегетативными, способами размножения. У плесневых грибов бесполое размножение может происходить частями гифы или мицелия, оидиями, хламидоспорами, склероциями, частями мицелиальных тяжей. Однако наибольшее значение имеет бесполый способ размножения спорами (спорангиоспорами иликонидиоспорами), образующимися в огромных количествах. Так, на 1 см2 поверхности плесневого налета может находиться несколько тысяч спорангиеносцев или конидиеносцев, и на каждом из них может образоваться по несколько тысяч спор. Каждая спора при благоприятных условиях через 4–7 дней может дать новый такой же налет со спорами. У фикомицетов на плодоносящих гифах, называемых спорангиеносцами, формируются спорангии, внутри которых находятся спорангиоспоры (см. рис. 41, а).
У аскомицетов и многих несовершенных грибов споры бесполого размножения – конидиоспоры или конидии, образуются экзогенно на кончиках плодоносящих гифов, называемых конидиеносцами, или конидиофорами. Размеры, форма, расположение цепочек и цвет конидий очень разнообразны и являются характерными признаками различных родов и видов высших грибов (см. рис. 41 б, в, г).
ФИЗИОЛОГИЯ ПЛЕСНЕВЫХ ГРИБОВ
Питание
Плесневые грибы в качестве источников углерода могут использовать самые разнообразные соединения: сахара и полисахариды, спирты, органические кислоты, аминокислоты, белки, углеводороды, производные фенолов и многие др.
В качестве источников азота кроме органического азота большинство плесневых грибов способны усваивать нитратный и аммиачный азот. Органический азот используется главным образом в виде аминного азота, находящегося в составе NH2-rpyппы аминокислот. Наилучшими источниками азота аминокислот являются аспарагиновая и глютаминовая кислоты и их амиды.
Использование белков возможно только после их гидролиза протеолитическими ферментами. Описаны отдельные виды грибов, способные фиксировать атмосферный азот. К их числу относится, например, Aspergillus flavus [23].
Для роста и жизнедеятельности плесневых грибов необходимы минеральные вещества. Одни из них необходимы в относительно высокой концентрации, к ним относятся сера, фосфор, калий, кальций, магний, железо. Другие, их называют микроэлементами, необходимы в низких концентрациях (обычно в мкг/л). К ним относятся марганец, молибден, цинк, медь, кобальт, никель, бор и многие др.
Потребности плесневых грибов в витаминах обычно менее выражены, чем у дрожжей. Наиболее часто наблюдается потребность в тиамине, биотине, пиридоксине и мезоинозите, в то время как многие дрожжи нуждаются еще в никотиновой и пантотеновой кислотах.
Ферменты
Плесневые грибы имеют богатый комплекс ферментов, позволяющий им использовать в качестве питательных субстратов многие вещества. Ферменты плесневых грибов с древнейших времен использовались человеком для осахаривания крахмала риса в восточных странах при производстве рисовой водки – саке, известной более 2500 лет. В Японии для этой цели применяют культуру гриба Aspergillus oryzae, образующего активную амилазу. В нашей стране и большинстве западных стран осахаривание крахмала зерна проводили при помощи амилаз проросшего зерна (солода). В настоящее время на большинстве спиртовых заводов вместо солода применяют культуры плесневых грибов.
В специально созданных условиях плесневые грибы способны синтезировать большие количества разнообразных ферментов. При получении амилаз, пектиназ, протеаз, целлюлаз, глюкозо-оксидазы, каталазы используют различные виды плесневых грибов родов Aspergillus, Penicillium, Rhizopus, Mucor.
Штаммы плесневых грибов одного и того же рода и даже вида могут резко различаться по количеству синтезируемых ферментов. Поэтому для промышленного производства ферментов проводят селекцию наиболее активных штаммов продуцентов ферментов путем отбора из природных условий, воздействия ультрафиолетовыми лучами или химическими мутагенами, а также подбирают оптимальный состав среды и условия культивирования.
Ферменты плесневых грибов используются в качестве биокатализаторов в виде ферментных препаратов в различных отраслях пищевой и легкой промышленности, бытовой химии, медицине, сельском хозяйстве. Сведения о производстве и применении ферментных препаратов можно найти в ряде руководств [155, 213].
Антибиотики
Образование антибиотиков имеет эволюционно приспособительный характер. Большинство антибиотиков, образуемых грибами, являются вторичными продуктами метаболизма, которые активно не участвуют в процессах конструктивного и энергетического обмена. Антибиотикообразующая способность большинства природных штаммов плесневых грибов обычно низкая, и только штаммы, полученные путем селекции, обладают высокой антибиотической активностью.
Из грибов разных систематических групп выделено свыше 100 антибиотиков. Однако только немногие нашли практическое применение и изучены более детально. К ним относятся пенициллины, гризеофульвин, фумагиллин и некоторые др. Современная медицина и ветеринария используют антибиотики для лечения некоторых заболеваний человека и животных. Антибиотики находят применение и как стимуляторы роста сельскохозяйственных животных и птиц [23].
ФАКТОРЫ, ВЛИЯЮЩИЕ НА РОСТ
И ФИЗИОЛОГИЧЕСКУЮ АКТИВНОСТЬ ПЛЕСНЕВЫХ ГРИБОВ
Температура
Температура является одним из главных факторов регулирования роста и физиологической активности плесневых грибов. Оптимальная температура роста плесневых грибов большинства видов не всегда бывает оптимальной для образования репродуктивных органов и физиологической активности. Амплитуда оптимальной температуры роста у разных видов грибов неодинакова. Так, для многих видов пенициллов она находится в пределах 26–28°С, для некоторых видов аспергиллов составляет 25–40°С. Виды аспергиллов более теплолюбивы по сравнению с видами пенициллов. Они в значительном количестве выделяются из почв южных зон.
Температура оказывает фунгистатическое и фунгицидное действие на грибы. Фунгистатическое действие оказывает обычно низкая температура, она не вызывает гибели грибов. Фунгицидное действие повышенной и высокой температуры зависит от pH и состава среды, количества микроорганизмов, их физиологического состояния и других факторов.
Споры и конидии плесневых грибов, находящиеся в жидкостях, значительно чувствительней к высоким температурам, чем в обезвоженных средах. Обычно пастеризация виноградного сока при температуре 85°С в течение 15 мин вызывает гибель плесневых грибов. Однако описаны случаи порчи пастеризованного сока плесневым грибом Byssochlamys fulva. Этот гриб обладает удивительной теплоустойчивостью. Взвесь его конидий в плодовых соках выдерживает действие температуры 85°С – в течение 20 мин, а для гибели аскоспор при температуре 85°С требуется 65 мин [223].
Свет
Прямой солнечный свет ингибирует рост плесневых грибов. Чередование освещения и темноты, наоборот, стимулирует 212 рост и спорообразование многих из них.
На грибы, как и другие микроорганизмы, влияют радиоактивные излучения. Устойчивость к излучению у различных таксономических групп грибов неодинакова. Более устойчивы темно-окрашенные грибы, которые содержат пигменты в оболочках конидий, склероциев и других структур.
Ионизирующее облучение влияет на изменение генетических свойств плесневых грибов и обусловливает появление мутаций. Облучение УФ-лучами применяется при получении высокопродуктивных мутантов плесневых грибов – продуцентов антибиотиков, ферментов и для получения биохимических мутантов, которые затем используют для выяснения путей биосинтеза многих метаболитов.
Кислотность среды
Величина pH среды является важным фактором роста микроорганизмов. Если для большинства бактерий оптимальной величиной pH среды является нейтральная и слабощелочная, то для плесневых грибов – кислая реакция среды (pH 4–5). В виноградном сусле, имеющем величину pH 3–4, большинство плесневых грибов быстро размножается, однако в вине с такой же величиной pH спирт ингибирует их развитие. Некоторые плесневые грибы, например Aspergillus niger, могут размножаться при pH среды от 2 до 8 [113].
Аэрация
Для своего нормального развития плесневые грибы нуждаются в кислороде воздуха, особенно для образования конидий. Среди плесневых грибов неизвестны облигатные анаэробы, но многие виды могут расти при пониженном содержании кислорода в среде. Например, некоторые виды Mucor, Penicillium, Aspergillus могут размножаться на среде с глюкозой при содержании кислорода около 0,1–0,2%. Гриб Merulius lacrimans, разрушающий дерево, более чувствителен к недостатку кислорода, допустимый минимум его составляет около 3%.
В условиях пониженного содержания кислорода у многих видов грибов изменяется морфология и фазы развития, так, например, у грибов рода Mucor мицелиальный тип роста трансформируется в дрожжеподобный. Углекислота тормозит размножение плесневых грибов.
Влажность
От относительной влажности воздуха в период созревания винограда зависит степень его поражения плесневыми грибами. В сырую дождливую погоду степень поражения ягод винограда всегда сильнее, чем в сухую солнечную. Замечено, что в периоды очень частых дождей преобладают на ягодах представители рода Penicillium, а в сухие солнечные месяцы – рода Aspergillus. Наличие видов рода Mucor довольно постоянно. Размножение гриба Botrytis cinerea на созревающих ягодах винограда находится в прямой зависимости от влажности воздуха: при дождливой погоде ягоды покрываются серой плесенью (порошкообразный налет из конидий), особенно у сортов с плотными гроздями, а при сухой и солнечной погоде налет на ягодах бывает скудным и мало заметным.
РОЛЬ ОТДЕЛЬНЫХ ПРЕДСТАВИТЕЛЕЙ
ПЛЕСНЕВЫХ ГРИБОВ В ВИНОДЕЛИИ
Наиболее часто на винограде встречаются плесневые грибы родов Aspergillus, Penicillium, Botrytis, Mucor, Sphaerulina.
В годы с повышенной влажностью воздуха размножение плесневых грибов вызывает значительные потери урожая винограда и понижение его качества. Особенно сильно размножаются плесневые грибы на ягодах винограда, если после долгой засушливой погоды начинается дождливый период. При таком резком переходе на ягодах образуется большое количество трещин, через которые проникают споры грибов. Прорастая, споры образуют мицелий, и виноград плесневеет.
В некоторых винодельческих районах, где, как правило, осенние дожди выпадают редко, раза два-три в десятилетие, бывают благоприятные погодные условия для так называемого «благородного гниения» винограда. В такие годы виноград рано созревает при достаточной, но не чрезмерной влажности и происходит заизюмливание ягод при умеренном развитии гриба Botrytis cinerea. Мицелий гриба разрыхляет клетки кожицы, облегчает испарение воды. При дождливой погоде из поврежденных грибом ягод вымывается сахар и качество винограда снижается, а при солнечной и сухой осенней погоде Botrytis cinerea способствует испарению воды из ягод винограда и повышению концентрации сахара в соке до 40% и более.
В отличие от плесневых грибов других родов Botrytis потребляет в первую очередь органические кислоты, а не сахара. Поэтому при повышении концентрации сахара в ягодах, происходящей от испарения воды, в них не наблюдается увеличения кислотности. Содержание пектиновых и фенольных веществ, а также общего азота уменьшается в зависимости от степени размножения Botrytis cinerea.
В связи с тем что «благородное гниение» развивается постепенно, сбор гроздей и даже отдельных ягод в надлежащей степени готовности приходится производить несколько раз. Вследствие этого приготовленные вина называются выборочными (ауслезевейн). Сотернские вина, особенно Шато-Икем, пользуются заслуженной славой и являются одними из самых дорогих вин в мире. Состав их в зависимости от условий года очень непостоянен: содержание сахара от 1,34 до 44%; спирта от 6,4 до 17,5% об., глицерина от 0,7 до 2,4% [144]. Вина мягкие, маслянистые во вкусе, со своеобразным букетом.
В связи с высоким качеством вин типа Шато-Икем и невозможностью получения их в других винодельческих районах были предприняты попытки искусственного создания условий, необходимых для «благородного гниения» винограда в специальных камерах. У нас в стране такие опыты проводил А. А. Преображенский [144], в Калифорнии К. Е. Нельсон и М. А, Америн [286]. Были подобраны оптимальные температура и относительная влажность для искусственного воспроизведения «благородного гниения» винограда, из которого получали вина типа сотернских. Однако технические затруднения служат препятствием для внедрения этого приема в производство.
Е. М. Поповой в течение многих лет проводились опыты по внесению ферментных препаратов, полученных из мицелия Botrytis, в виноградное сусло и вино вначале с целью замены «благородного гниения» винограда, а затем для гидролиза пектиновых веществ и ускорения созревания вин. Изучался биохимизм процессов, протекающих в сусле и вине при внесении препаратов [143]. Позднее были отобраны штаммы Botrytis cinerea – активные продуценты комплексного ферментного препарата, обладающего пектолитической, целлюлолитической и протеолитической активностью [226].
Ботритис образует еше вещества, названные «ботритицином», обладающие антибиотической активностью в отношении дрожжей. В соке, полученном из винограда, пораженного грибом, образуется меньшая масса дрожжей и задерживается брожение. Ускорить брожение такого сусла можно сульфитацией (введением 50 мг/л SO2) или продолжительным нагреванием до 120ДС [154].
В винограде, пораженном грибом В. cinerea, образуется много оксидаз, вызывающих впоследствии побурение и оксидазный касс в белых столовых винах.
Чем больше виноград поражен плесневым грибом, тем больше связанной и меньше свободной сернистой кислоты получается при сульфитации сусла. Поэтому чем больше степень поражения винограда, тем большую дозу сернистого ангидрида нужно вводить в сусло. Так, если для сусла из винограда, пораженного плесневым грибом на 10%, для инактивации оксидаз достаточна доза сернистого ангидрида 150 мг/л, то для сусла из винограда, пораженного плесневым грибом на 40%, необходима уже доза сернистого ангидрида 250 мг/л.
Ж. Блоуэн считает, что уксуснокислые бактерии, находящиеся на винограде, пораженном плесневым грибом, являются в большей степени, чем плесневые грибы, причиной образования карбонильных соединений, наиболее легко связывающих сернистую кислоту [237].
Г. Дитрих, изучавший продукты брожения сусла, полученного из здоровых и пораженных грибом ягод, показал, что в последнем случае образовалось в 3,5 раза больше пировиноградной кислоты, в 2 раза больше кетоглютаровой кислоты и на 58% больше ацетальдегида [245].
Эти кислоты и ацетальдегид легко связывают SO2, и поэтому для инактивации окислительных ферментов и предотвращения побурения столового вина и оксидазного касса требуется сульфитация его повышенными дозами сернистой кислоты.
Окислительные ферменты из сусла, полученного из винограда, пораженного ботритисом, можно удалить путем обработки его бентонитом (3–5 г/л) с полиакриламидом (2 мг/л). Сусло и вино из винограда, пораженного на 20% плесневым грибом, после такой обработки имели хороший вкус и окраску (исчезали коричневые и бурые тона).
Для удаления плесневого тона и оксидазного касса в винах без применения сернистой кислоты рекомендуется нагревание до 55–60°С в течение 15 мин, затем охлаждение и обработка активным углем (1 г/л), диатомитом (1 г/л), желатином (0,1 г/л) и бентонитом (1 г/л).
Вина, имеющие в аромате и во вкусе сильные плесневые тона и пораженные оксидазным кассом, можно исправить, выдерживая их под хересной пленкой. Перед этим вина пастеризуют при температуре 70°С 5 мин [32].
Таким образом, размножение гриба Botrytis cinerea на винограде бывает полезным только в редкие годы в определенных винодельческих районах для выработки вин особого типа. Во всех остальных случаях размножение на винограде плесневых грибов всех родов снижает его качество.
Sphaerulina intermixta при размножении в виноградном соке может превратить сок в слизистую массу с содержанием спирта около 2% об. В вине этот гриб не размножается, так как при содержании спирта свыше 2% об. его жизнедеятельность прекращается. Гриб часто встречается в слизистом налете на стенах подвалов, на бочках.
* * *
На винодельческих заводах во избежание появления нежелательной микрофлоры – плесневых грибов – следует соблюдать чистоту.
В винподвалах размножение плесневых грибов так же вредно, как и на винограде. Споры плесневых грибов родов Mucor, Aspergillus, Penicillium, обычно всегда находящиеся в воздухе, легко прорастают на остатках сусла и вина, своевременно не удаленных из емкостей, с оборудования, из шлангов. При размножении плесневые грибы образуют летучие вещества, которые при контакте с вином в нем растворяются и придают винам очень неприятный трудно устранимый плесневый тон.
Для предотвращения размножения плесневых грибов необходимо тару и оборудование сразу после освобождения от сусла и вина тщательно промыть водой. Внутренняя поверхность деревянной и железобетонной тары без защитных покрытий обычно пористая, пропитанная остатками вина или сусла, трудно вымываемыми из пор. При достаточной влажности внутренние поверхности в таких емкостях быстро плесневеют. Поэтому для предотвращения размножения плесневых грибов деревянную тару, находящуюся без употребления, после тщательной мойки окуривают сернистым ангидридом или хранят заполненной 0,1%-ным раствором сернистой кислоты. Надежно устраняется пористость внутренних поверхностей железобетонных резервуаров при помощи синтетических защитных покрытий эполукс, эполор, эпросин, ХС-558 «В» и др.
Бродильные чаны протирают насыщенным раствором кальцинированной соды. Железобетонные емкости после тщательной мойки необходимо хранить с открытыми верхним и нижним люками. Приемы обработки заплесневевшей тары описаны в «Технологической инструкции по санитарной обработке винодельческих емкостей, оборудования винопроводов и помещени;» [184].
Одним из новых и перспективных направлений в современном виноделии является применение ферментных препаратов из плесневых грибов. Установлено, что обработка виноградной мезги ферментными препаратами грибов Aspergillus awamori (Пектаваморином П10Х, Пектаваморином Г10Х), Aspergillus foclidus (Пектофоетидином П10Х, Пектофоетидином Г10Х) и Botrytis cinerea (Пектоцинерином Г 10Х) ускоряет процесс отделения сусла, увеличивает его общий выход, ускоряет процесс осветления сусла и молодых виноматериалов, а также созревание вин.
В настоящее время изучается действие отдельных ферментных систем, входящих в состав препаратов, с целью оптимизации их состава для использования при производстве вин различных типов.
Ферментные препараты ускоряют процесс гидролиза белков и полисахаридов. Уменьшение количества полимеров в результате их гидролиза происходит в период осветления сусла, во время его брожения и дальнейшего хранения виноматериалов. Получаемые с применением ферментных препаратов виноматериалы быстрее осветляются, требуют меньше затрат при обработке и более стабильны к помутнениям.
