Лактококки (по старой номенклатуре мезофильные молочнокислые стрептококки) являются основными представителями необходимой микрофлоры для большинства сыров за исключением твердых сыров с высокими и средними температурами II нагревания. Опыты по выработке сыров из асептически полученного молока в асептических ваннах, предохраняющих сыр во время выработки от загрязнения посторонней микрофлорой, показали, что лактококки являются необходимой и достаточной микрофлорой для производства сыров с низкими температурами II нагревания. Они являются единственной микрофлорой, участвующей в производстве большинства кисломолочных сыров.
В 9-ом издании определителя бактерий Берджи род Lactococcus входит в группу 17 «Грамположительные кокки». Дифференциальные признаки рода: факультативные анаэробы, метаболизм бродильного типа, сбраживают углеводы с образованием в основном L(+)-молочной кислоты без выделения газа, растут при 10° С, не растут при 45° С и в присутствии 6,5% NaCl. Лактококки являются мезофильными гомоферментативными бактериями. Типовой вид Lactococcus lactis имеет три подвида: Lactococcus lactis subsp. lactis; Lactococcus lactis subsp. cremoris и Lactococcus lactis subsp. diacetylactis (для краткости будем их называть молочный, сливочный и диацетильный лактококки, либо Lc. lactis, Lc. cremoris и Lc. diacetylactis). В 9-ом издании определителя бактерий Берджи приведен Lactococcus lactis subsp. hordinae, не сбраживающий лактозу, о роли которого в сыроделии ничего неизвестно, но исключен Lc. lactis subsp. diacetylactis, имеющий исключительно важное значение в производстве сыра. Исключение диацетильного лактококка из классификации следует рассматривать как ошибку. Кроме молочного лактококка с подвидами в группу лактококков включены четыре вида Lc. garviese, Lc. piscium, Lc. rqffinolactis и L plantarum. Значение для сыроделия первых трех видов не выявлено, клетки L plantarum обычно имеют форму коротких палочек и этот вид рассматривается в группе лактобацилл.
Лактококки в среде с достаточной концентрацией глюкозы или лактозы при pH 7,0 и 30° С в анаэробных условиях образуют из сброженного сахара до 95% молочной кислоты; в средах с ограниченным содержанием сахаров они сбраживают их по гетероферментативному пути, образуя, кроме молочной, уксусную и муравьиную кислоты, этанол.
Лактококки - факультативные анаэробы, получают энергию только путем брожения. Большинство микроорганизмов могут получать энергию разными путями, что обеспечивает им возможность существования в различных условиях. Поэтому более правильно говорить не о гомо- или гетероферментативных бактериях, а о том, каким путем они получают энергию. Лактококки в оптимальных условиях получают энергию из глюкозы или лактозы гомоферментативным путем, в неблагоприятных - гетероферметативным путем. В опытах Collins в среде, содержащей 0,18% глюкозы, лактококки сбраживали в уксусную кислоту до 14% сахара. При низкой степени разбавления среды в хемостате, приводящей к дефициту углеводов, лактококки могут трансформировать в молочную кислоту только 1% сброженной глюкозы. Пути сбраживания глюкозы и лактозы лактококками зависят от температуры: при 30-37° С глюкоза сбраживается по гомоферментативному, при 15-20° С - по гетероферментативному пути. Таким образом, во время созревания сыров лактоза сбраживается по гетероферментативному пути, что объясняет постоянное наличие уксусной кислоты в твердых сычужных сырах. По гетероферментативному пути большинство штаммов лактококков сбраживает галактозу, мальтозу, рибозу, глюконат.
Диагностические характеристики лактококков приведены в табл. 3.3. Филогенетически все три подвида близко родственны и образуют четко очерченный таксон. Однако между подвидами четкого разделения не существует. Из заквасок и ферментированных молочных продуктов часто выделяют штаммы лактококков с промежуточными свойствами. Это происходит потому, что такие свойства лактококков, как способность ферментировать лактозу, цитраты, сахарозу, галактозу, протеиназная активность, способность синтезировать антибиотик низин и, возможно, гидролизовать аргинин, закодированы на плазмидной ДНК, репликация которой происходит независимо от репликации хромосомной ДНК. При потере клеткой плазмиды дочерние клетки не получают ее копии и теряют способность выполнять ту функцию, которая была закодирована на данной плазмиде. Лактококки, выделенные из сырого молока, по способности сбраживать различные источники углерода можно разделить на 15, 35 и 13 физиологических групп соответственно Lc. lactis subsp. cremoris, Lc. lactis subsp. lactis и Lc. lactis subsp. diacetylactis. Встречаются штаммы молочного лактококка, неспособные гидролизовать аргинин. Потеря этого свойства часто сопровождается приобретением способности утилизировать цитраты. Из заквасок выделяли штаммы сливочного лактококка (по старой номенклатуре Str. cremoris), которые росли в бульоне с 4% NaCl, сбраживали мальтозу и декстрин, образовывали небольшие количества диацетила. Некоторые авторы считают, что лактококки образуют фенотипически и генотипически непрерывный спектр с концентрацией около трех точек, которые условно назвали подвидами lactis, diacetylactis и cremoris. Показательно, что около 30% бактериофагов, выделенных на сыродельных заводах, способны лизировать штаммы всех трех подвидов лактококков.
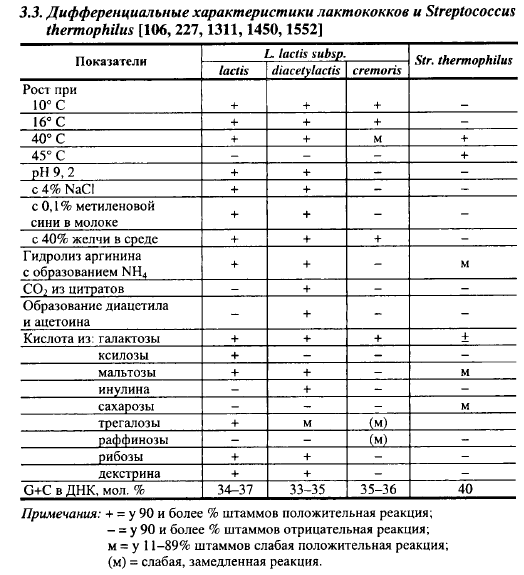
Главным отличием Lc. lactis subsp. diacetylactis от двух других подвидов является его способность утилизировать цитраты с образованием С02, диацетила и ацетоина. Это отличие имеет принципиальное значение, так как образуемый этим подвидом С02 обусловливает формирование рисунка в наиболее распространенных твердых сырах, а рисунок является важным и легко распознаваемым потребителем показателем качества продукта. Диацетил (но не ацетоин) принимает участие в формировании аромата сыров. Благодаря образованию диацетила диацетильный лактококк и лейконостоки часто называют ароматообразующими, а два других вида - кислотообразующими микроорганизмами. Это справедливо для кислосливочного масла, аромат которого зависит в первую очередь от содержания в нем диацетила, в сырах же роль диацетила значительно скромнее. В производстве Чеддера, например, ароматообразующие молочнокислые бактерии не применяют, потому что образование ими С02 приводит к появлению «открытой» структуры (появлению трещин в сырной массе), которая считается большим пороком этого сыра. Диацетил в Чеддере часто вообще не обнаруживается после первых двух недель созревания. Несмотря на это, Чеддер обладает хорошо выраженными ароматом и сырным вкусом. В то же время активные по кислотообразующей способности штаммы диацетильного лактококка вносят в образование кислоты при производстве сыров такой же вклад, какой вносят два других подвида лактококков. Диацетильный лактококк и лейконостоки лучше называть не ароматообразующей, а газообразующей микрофлорой заквасок.
В процессе культивирования, особенно на молочных средах, диацетильный лактококк может потерять способность синтезировать цитрат пермеазу, осуществляющую транспорт цитратов через клеточную мембрану, и тем самым перестать утилизировать цитраты. Однако и в этом случае у него обнаруживается цитратаза, отсутствующая у молочного лактококка. Нестабильность способности диацетильного лактококка утилизировать цитраты обусловлена тем, что она закодирована на плазмидах.
Несмотря на способность образовывать С02 из цитратов, диацетильный лактококк является гомоферментативным микроорганизмом, так как гомоферментативность и гетероферментативность являются характеристиками основного пути получения бактерией энергии из глюкозы, а глюкозу он сбраживает подобно другим подвидам лактококков в молочную кислоту (до 95% от общего количества сброженной лактозы) без образования С02. Для сбраживания цитратов в молоке диацетильный лактококк нуждается в Мп2+.
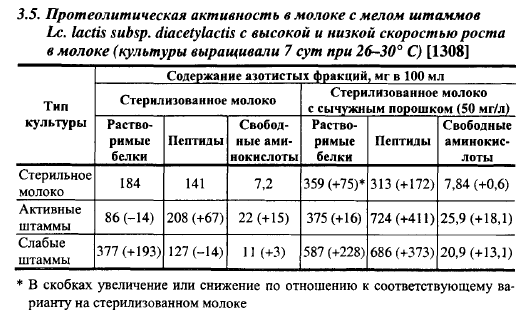
Диацетильный лактококк отличается от Lc. lactis subsp. lactis по протеолитической активности, скорости роста в молоке (табл. 3.5). По скорости роста в молоке диацетильный лактококк разделяется на две группы: свертывающие молоко при посевной дозе 3% и оптимальной температуре за 4-9 ч («активные» штаммы), т. е. с такой же скоростью, как активные штаммы молочного лактококка, и свертывающие молоко в этих условиях за 14-36 ч («слабые» штаммы). При посевной дозе 1% и температуре 30° С активные штаммы повышали кислотность молока за 5 ч до 40-54° Т, слабые - до 32-34° Т. По количеству образуемого в молоке диацетила слабые штаммы можно разбить на две группы: представители первой группы образовывали в молоке 0,5-0,75 мг% диацетила, второй - 0,139-0,233 мг%. Слабые и активные штаммы отличаются по протеолитической активности в молоке (табл. 3.5). Активные по росту в молоке штаммы диацетильного лактококка подобно молочному и сливочному лактококкам в процессе размножения в молоке снижали содержание растворимых белков и увеличивали содержание пептидов и свободных аминокислот, а слабые штаммы, наоборот, увеличивали содержание растворимых белков и снижали содержание пептидов при незначительном увеличении содержания свободных аминокислот. Эта особенность штаммов диацетильного лактококка с низкой скоростью кислотообразования в молоке имеет большое значение для сыроделия, так как горечь в сырах чаще всего связана с образованием пептидов, обладающих горьким вкусом. Включение в состав заквасок для производства твердых сыров с низкими температурами II нагревания слабых штаммов диацетильного лактококка уменьшает опасность появления в них горького вкуса.
Штаммы диацетильного стрептококка с низкой скоростью роста в молоке отличаются от активных штаммов не только протеолитической активностью, но и более высоким уровнем образования диацетила в молоке по отношению к образованию ацетоина, хотя среди них имеются штаммы, также образующие только небольшие количества диацетила.
Сливочный лактококк в дополнение к диагностическим признакам, приведенным в табл. 3.3, отличается от двух других подвидов морфологически: 19,7% клеток сливочного лактококка объединены в короткие цепочки (по 3-5 клеток), 51,1% - в более длинные цепочки, а у молочного и диацетильного лактококков только 10,2 и 6,3% соответственно объединены в короткие цепочки (до 5 клеток), а более длинных цепочек в их культурах вообще нет.
Сливочный лактококк отличается от молочного и диацетильного меньшей метаболической активностью и более низкой устойчивостью к внешним факторам, особенно к содержанию в среде NaCl и активности воды, а также к температуре (табл. 3.4). Его можно рассматривать как вариант молочного лактококка, наиболее адаптированный к молоку. Показательно, что в сыром молоке, отобранном на фермах, чаще встречается молочный, на заводе - сливочный лактококки. Это доказывает, что сливочный лактококк нашел свое место в нише молокоперерабатывающих предприятий.
Многие штаммы сливочного лактококка имеют максимальную температуру для роста в молоке ниже 40° С (35-39° С). Так, из 9 штаммов сливочного лактококка ни один не дал заметного роста при 40° С и только один размножался при 39° С . Повышение температуры инкубации с 28-31° С до 35,6° С увеличило продолжительность времени генерации у трех штаммов сливочного лактококка в 5,1-5,3 раза, у двух - в 2,6-2,9 раза, еще у двух - в 1,7-1,9 раза, в то время как у штаммов молочного лактококка - только в 1,2 раза. Штаммы сливочного лактококка более вариабельны по отношению к максимальной температуре для роста, чем штаммы молочного лактококка: некоторые из них хорошо растут при 38° С, другие растут очень медленно при температурах выше 36° С. В молоке с добавлением 0,5-1,0% дрожжевого экстракта многие штаммы сливочного лактококка росли при 37,5° С с такой же скоростью, как при 31,5° С, но только в течение первых двух часов, после чего их рост прекращался или шел с очень низкой скоростью. Это свидетельствует о потере сливочным лактококком при повышенных температурах способности к синтезу какого-то фактора роста, содержащегося в небольших количествах в дрожжевом экстракте.
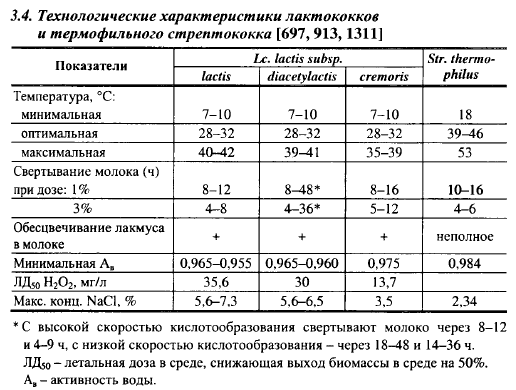
Максимальная температура для роста лактококков, по существу, определяет температуру II нагревания при производстве мелких сыров. За рубежом II нагревание при выработке этих сыров проводят при 35° С, т. е. температуре, достаточно благоприятной для кислотообразования и накопления биомассы всех лактококков, в т. ч. Lx. cremoris. В России такая температура недостаточна для достижения необходимой степени синерезиса сычужного сгустка в производстве сыров в связи с невысоким содержанием казеина и высоким содержанием соматических клеток в молоке (гл. 2 и 7), поэтому ее повышают до 38-42° С. За рубежом такие температуры II нагревания применяют только при производстве Чеддера ускоренным способом. Они слишком высоки для Lc. cremoris и поэтому основу большинства российских заквасок для сыров с низкими температурами II нагревания составляют два других подвида лактококков: Lc. lactis и Lc. diacetylactis, хотя и для многих штаммов этих подвидов температуры 41-42° С выше максимальных для роста.
Австралийские ученые нашли, что выдержка микрофлоры мезо- фильных заквасок во время II нагревания при температурах до 43° С не отражается на ее последующей способности сбраживать углеводы в сыре. В другом опыте стерилизованное молоко инокулировали мезо- фильными заквасками и выдерживали 30 мин при 45, 47 и 50° С, затем выдерживали при 30° С с измерением кислотности после 2, 4, 9 и 24 ч выдержки. Кислотность в конце опыта во всех вариантах сравнялась, но скорость нарастания кислотности уменьшалась при повышении температуры тепловой обработки. Отсутствия длительного торможения развития лактококков в сыре в результате применения повышенных температур II нагревания можно было ожидать, так как повышение температур по отношению к максимальным для роста лактококков невелико, а микроорганизмы обладают способностью репарировать сублетальные повреждения. Однако для репарации повреждений требуется время, пропорциональное их тяжести, поэтому замедление развития лактококков при высоких температурах II нагревания на какое-то время вполне естественно, а любое замедление роста лактококков во время выработки сыра оказывает крайне негативное влияние на показатели качества и безопасности сыров. Рамазанов с соавт. получили такую зависимость содержания молочнокислых бактерий (в основном молочного и диаце- тильного лактококков) в Голландском круглом сыре после прессования от температуры II нагревания:

В опытах Раманаускаса с соавт. свертывание молока при производстве твердых сыров с низкими температурами II нагревания проводили при 30, 32, 36, 40, 42 и 44° С с использованием мезофильных заквасок, в состав которых входили все три подвида Lc. lactis. Скорость накопления общего количества и количества ароматообразующих молочнокислых бактерий с увеличением температуры снижалась, а при 42-44° С отмечалось резкое торможение их развития. При температуре свертывания 42° С количество ароматообразующих бактерий в зерне в конце обработки было в 9, при 44° С - в 25 раз меньше, чем при 32° С.
Следует сказать, что только по скорости кислотообразования нельзя судить о влиянии субоптимальных факторов на развитие лактококков. При 32° С рост биомассы сливочного лактококка соответствует количеству образуемого им лактата. Повышение температуры, высокие концентрации соли и Н+ разобщают эти процессы и могут полностью подавить размножение, не оказывая столь сильного влияния на кислотообразование. Полное прекращение размножения сливочного лактококка при довольно высокой скорости кислотообразования наблюдается при концентрации соли больше 3%, температуре 38-40° С. Явление разобщенности процессов размножения и кислотообразования при субмаксимальных температурах наблюдается и в культурах Lc. lactis (А. Гудков, Г. Д. Перфильев, неопубликованные материалы). Некоторые считают, что при супероптимальных для лактококков температурах II нагревания часть клеток отдельных штаммов сливочного лактококка погибает. По-видимому, при повышенных температурах происходят прогрессирующие повреждения клеток, приводящие к расстройству их катаболитических и анаболитических функций, относительно независимых друг от друга, причем скорость нарушения анаболизма значительно выше. Возможно, при повышенных температурах у чувствительных штаммов инактивируются пермеазы, контролирующие транспорт аминокислот через клеточную мембрану.
Таким образом, анализ влияния близких к максимальным или немного превышающих максимальные температур на рост и кислотообразующую активность лактококков показывает, что лучше всего температуру во время выработки сыров с низкими температурами II нагревания поддерживать на уровне, не превышающем 36° С. При необходимости интенсифицировать синерезис температуру II нагревания можно повысить до 40° С, применив при этом закваску, в которой доминирующую роль играют штаммы молочного и диацетильного лактококков, максимальная температура для роста которых выше, чем для сливочного лаклактококка.
Методами экспериментальной селекции, например, часовой обработкой этилметансульфонатом при 25° С, из штаммов лактококков, неспособных расти при 38^Ю° С, удается получить мутанты лактококков, способные образовывать кислоту при этих температурах. Особый интерес вызывают мутанты, быстро лизирующиеся при температуре 40° С, и таким образом высвобождающие внутриклеточные энзимы в самом начале созревания, когда условия для биохимических процессов в сыре наиболее благоприятны.
В твердых сырах с высокими и средними температурами II нагревания (48-57° С) большинство клеток мезофильных лактококков погибает во время II нагревания. В опытах Каган и Чистяковой при производстве сыра Советский во время II нагревания (52-55° С, 20-30 мин) количество лактококков снизилось с 3-107до 3,7-103/г, или в 811 раз. Однако, гибель большей части клеток лактококков во время II нагревания при производстве крупных сычужных сыров - еще недостаточный повод для их исключения из состава заквасок для производства сыров этой группы, так как они производят значительное количество L(+)-молочной кислоты до своей гибели. Исключение их из состава заквасок может изменить соотношение между L(+)- и DL-mo- лочной кислотой, образуемой некоторыми термофильными лактобациллами, что, в свою очередь, может отразиться на развитии пропионово- кислых бактерий, которые утилизируют L(+)лактат быстрее, чем Б(-)-лактат. Термофильный стрептококк, который образует правовращающую молочную кислоту, не может служить равноценной заменой лактококкам, так как обладает высокой чувствительностью к наличию в молоке ингибирующих веществ и к бактериофагам, а также несколько медленнее растет при температурах свертывания молока сычужным энзимом. Замена лактококков закваски для крупных сыров термофильным стрептококком приводит к увеличению содержания галактозы в сыре (лактококки сбраживают лактозу без накопления в среде галактозы, а термофильный стрептококк ферментацию лактозы начинает с ее гидролиза в среде на глюкозу и галактозу, утилизация галактозы идет после сбраживания глюкозы). Большое количество галактозы, остающейся в сыре, может изменить соотношение между мезофильными и термофильными молочнокислыми бактериями в сыре и повлиять на его качество. В то же время слишком интенсивное развитие лактококков может привести к появлению горечи в крупных сырах, которая не встречается при правильном соотношении между лактококками и термофильным стрептококком.
На супероптимальные для лактококков температуры по-разному реагируют штаммы, образующие и не образующие горечь в сырах: «горькие» штаммы сливочного лактококка хорошо росли и быстро образовывали кислоту при 31,5 и 37,5° С, а «негорькие» штаммы при 37,5° С прекращали рост через 2 ч, продолжая образовывать молочную кислоту. Добавление в молоко 1% дрожжевого экстракта не стимулировало размножения негорьких штаммов при повышенных температурах. Явление разобщенности процессов роста и кислотообразования следует использовать для отбора в состав мезофильных заквасок для производства сыров штаммов лактококков, образующих кислоту, но прекращающих размножение при повышенных температурах II нагревания, что может уменьшить вероятность появления горечи в сырах и повысит устойчивость закваски к бактериофагу (репродукция бактериофагов происходит только в размножающихся культурах).
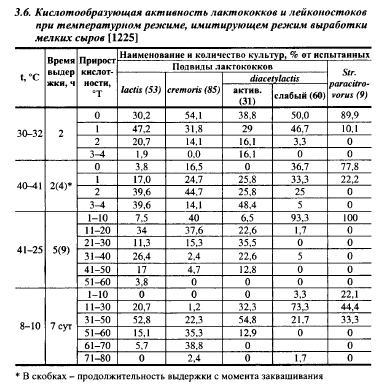
Изучено развитие лактококков и лейконостоков в пастеризованном (74-75° С) молоке с сычужным порошком при температурном режиме, имитирующем температурный режим выработки твердых сыров с низкой температурой II нагревания. Молоко после инокуляции 0,5% исследуемой культуры выдерживали 2 ч при 30-32° С, нагревали 10-15 мин до 40-41° С, выдерживали при этой температуре 2 ч, после чего оставляли на 5 ч при комнатной температуре, а затем инкубировали при 8-10° С в течении 7 сут. Кислотообразование культурами при этом режиме показано в табл. 3.6. Приведенные данные свидетельствуют о более высокой кислотобразующей активности молочного и активного диацетильного лактококков по сравнению со сливочным в первые 9 ч инкубации при комбинированном режиме: через 4 ч инкубации (после выдержки при температурах II нагревания) кислотность молока на 3-4° Т повысили 39,6 и 48,4% штаммов первых двух подвидов соответственно и только 14,1% сливочного лактококка; после 9 ч инкубации кислотность повысилась более чем на 31° Т в 47,2% культур молочного, 35,4% активного диацетильного и 7,1% сливочного лактококков. Положение изменилось после 7-суточной выдержки при 8-10° С: в конце этого периода только в 5,7% культур молочного лактококка кислотность повысилась на 61-80° Т по сравнению с 41,2% культур сливочного лактококка. Авторы работы не объясняют этот феномен. Возможно, при этих температурах степень гетеротрофное процесса сбраживания лактозы у сливочного лактококка ниже, а выход кислоты выше, чем у молочного и, особенно, диацетильного лактококка. Если это так, то свидетельствует о преимуществах сливочного лактококка, так как продукты гетероферментативного брожения участвуют в синтезе соединений, вызывающих пороки в сырах (гл. 12).
Определенный интерес для сыроделия представляет минимальная температура для роста лактококков, равная для всех видов 7-10° С. При 8° С размножались 50% штаммов молочного лактококка, выделенных из Голландского сыра, при этом 20% повышали кислотность молока при этой температуре за 7 сут на 30-60° Т, 20% - на 61-80° Т, 10% - более чем на 80 °Т; 67,7% штаммов сливочного лактококка росли при 8° С. Оптимальные температуры для созревания большинства твердых сыров выше 10° С, следовательно, температуры созревания сыра не препятствуют росту лактококков.
Из пастеризованного молока были выделены штаммы молочного лактококка, способные расти на агаре из гидролизованного молока при 5° С. Интересно, что в сыром молоке такие штаммы не были обнаружены. Вероятно, они попали в пастеризованное молоко после пастеризации, а следовательно, местом обитания таких штаммов является сам завод. Позднее о выделении из пастеризованного молока психротрофных вариантов всех трех подвидов лактококков сообщили и другие авторы. Интересно, что эти штаммы проявляли более высокую протеолитическую активность при 5° С, чем при 20° С. Их кислотообразующая активность при 5° С была очень низкой. Появились предложения включить такие штаммы в состав мезофильных заквасок для сыра и снизить температуру созревания мелких сычужных сыров, что могло бы ограничить возможности роста технически вредной поверхностной микрофлоры. Однако снижение температуры резко замедляет созревание сыра, а также гибель патогенной и условно патогенной микрофлоры, полной гарантии отсутствия которой в сыре на начальных этапах созревания при современном способе производства дать нельзя. Во время созревания большинство этой микрофлоры погибает, поэтому в США разрешается вырабатывать сыры из непастеризованного молока с низкой бактериальной обсемененностью, если они созревают при определенных температурах не менее 60 сут. При низких температурах созревания скорость отмирания патогенной микрофлоры резко снижается.
Психротрофные лактококки могут найти применение при созревании при низких температурах пастеризованного молока, предотвращающем репарацию сублетальных повреждений клеток посторонней микрофлоры, полученных в результате пастеризации, а следовательно, их реактивацию и размножение во время созревания.
Для молочного лактококка обычно характерна меньшая продолжительность фазы задержки роста и более высокая скорость кислотообразования в молоке, чем у сливочного лактококка. Так, в молоке с сычужным порошком при посевной дозе 0,5% при 30-32° С не дали прироста кислотности за 2 ч 30% штаммов молочного и 54% сливочного лактококков. Количество же штаммов, повысивших за это время кислотность молока на 2° Т и более, у первого подвида равнялось 23, у второго - 14%. При 3 %-ной посевной дозе прирост кислотности за первые 6 ч инкубации при 30-32° С у штаммов молочного лактококка составил 14—21° Т, сливочного - 14-16° Т, диацетильного - 10-20° Т. От стартовой скорости развития лактококков при производстве сыра во многом зависит качество продукта, так как во время выработки сыра в сырной ванне условия для их размножения оптимальны и, если они не будут использованы лактококками в максимальной степени, то значительно больше лактозы будет сброжено посторонней микрофлорой или лактококками во время созревания сыра, когда лактоза сбраживается ими по гетероферментативному пути из-за низкой температуры. Образование в сырах больших количеств продуктов гегероферментативного брожения усиливает опасность появления многих пороков сыра.
Скорость кислотообразования (моль молочной кислоты/ч) в молоке в условиях, моделирующих временные и температурные режимы выработки и прессования российских сыров с низкими температурами II нагревания, у штаммов молочного и диацетильного (активного) лактококков (90,6% изученных штаммов) находилась в интервале 5,5-11,5, у штаммов сливочного лактококка - 1,0-5,5, у слабых штаммов диацетильного лактококка 1,05-3,50. Эти различия для сливочного лактококка обусловлены прежде всего его отрицательной реакцией на температуры нагревания (39-41° С), а у слабых штаммов диацетильного лактококка, вероятно, недостаточной протеолитической активностью и низким содержанием в молоке низкомолекулярных азотистых соединений, необходимых для роста лактококков. Внесение в восстановленное молоко для выработки сыра 0,1% дрожжевого автолизата сделало возможным выработать сыр на «медленных» штаммах лактококков, при этом выход сыра увеличился. Объяснить увеличение выхода можно тем, что «медленные» штаммы лактококков обладают низкой протеолитической активностью в молоке, и поэтому при их использовании в качестве закваски потери белка с сывороткой ниже, чем при использовании быстрорастущих в молоке штаммов лактобактерий, которые обладают высокой протеолитической активностью в молоке. Обогащение молока дрожжевым экстрактом увеличило скорости размножения и кислотооб- разования «медленными» штаммами лактококков. Как указывалось выше, лактококки на начальных этапах роста в молоке, по крайней мере в течение первых 2-3 генераций, не могут расщеплять казеин и поэтому нуждаются в низкомолекулярных пептидах или/и свободных аминокислотах, содержание которых в молоке может быть недостаточным для полного удовлетворения потребностей, особенно сливочного и слабого диацетильного лактококков (разд. 3.2.3). Поэтому добавление в молоко низкомолекулярных азотистых соединений стимулирует развитие и «быстрых» лактококков. При совместном развитии в молоке большинство штаммов сливочного и молочного лактококков, исключая низинобразующие, стимулируют рост друг друга и других видов лакто- кобактерий с низкой скоростью роста в молоке.
Сливочный лакто кокк менее устойчив к соли. Отсутствие роста его в бульоне с 4% NaCl является диагностическим признаком (табл. 3.4), так как два других подвида лактококков растут в среде с 5,0-5,5% соли. Имеются данные о способности сливочного лактококка образовывать кислоту, но не размножаться при содержании в среде более 4% соли.
Более точным показателем устойчивости микроорганизмов к соли является минимальная активность воды для роста. Этот показатель зависит от концентрации и химического состава всех растворенных в водной фазе веществ. Минимальная активность воды для роста молочного лактококка равна 0,955, а для сливочного лактококка - 0,975, что соответствует активности воды в сыре Российский после прессования с содержанием 42-44% влаги и соответственно 3,5 и 2,0% соли. В табл. 3.4. приведены значения минимальной активности воды для роста лактококков, полученные в питательных средах с NaCl. В средах с использованием в качестве водосвязывающего вещества глицерина они значительно ниже (0,924 для молочного лактококка). Следовательно, соль ингибирует рост лактококков не только за счет связывания воды в среде. Особого значения устойчивость к соли лактококков в производстве отечесгвенных твердых сыров не имеет, так как рост их в сыре заканчивается намного раньше, чем соль распределится по всей массе головки сыра. Исключением является сыр Российский с полной посолкой в зерне, производство которого в настоящее время запрещено именно в связи с ингибированием солью роста лактококков. Есть данные, что 1-2% NaCI в среде стимулируют кислотообразование диацетильного лактококка.
Чрезвычайно важна устойчивость лактококков к соли в производстве Чеддера, который равномерно просаливается в конце выработки перед прессованием. Сливочный стрептококк сбраживал лактозу в этом сыре, если концентрация соли в водной фазе сыра не превышала 4% (около 1,6% в сыре). В сырах с 6% соли в водной фазе лактоза очень долго оставалась несброженной при использовании для выработки закваски из сливочного лактококка, и сбраживали ее не микроорганизмы закваски, а педиококки.
Молочный лактококк медленнее вымирает в сухих и замороженных заквасках во время хранения, менее чувствителен к перекиси водорода, хотя этот показатель в большой степени зависит от штамма. Некоторые виды и штаммы молочнокислых палочек могут образовывать в среде такое количество перекиси водорода, которое существенно снижает активность сливочного лактококка при совместном его росте с лактобациллами. Устойчивость сливочного и молочного лактококков к Н202 можно повысить их выдержкой в присутствии сублетальных концентраций перекиси или часовой экспозицией в аэробных условиях. Это очень важно с точки зрения разработки биологических методов борьбы с посторонней микрофлорой (разд. 3.2.3).
Лактококки отличаются довольно высокой устойчивостью к кислотности среды. Минимальный pH для них равен 4,2-Л,~4, предельная титруемая кислотность в молоке 100-120° Т. Однако эти пределы установлены по прекращению образования кислоты лактококками, а размножение их прекращается раньше, при pH ~4,5. Выдержка лактококков в среде с pH меньше 5 вызывает существенные изменения в клетках и увеличивает продолжительность фазы задержки роста при последующих пересадках [628]. Максимальную активность при пересадках показывают культуры при pH 5,0, когда количество жизнеспособных клеток достигает достаточно высокого уровня, а их активность еще не начала снижаться. Оптимальный pH для роста лактококков в молоке и бульоне 6,3-6,5, скорость размножения существенно снижается при pH ниже 6 или выше 7. Предельная кислотность в молоке зависит от подвида и штамма: для 16 штаммов лактококков, исследованных по этому показателю в отделе микробиологии ВНИИМС, она составила от 93 (штаммы диацетильного лактококка с низкой скоростью кислотообразования) до 115° Т, минимальный pH изменялся от 4,46 до 4,22. У большинства штаммов диацетильного лактококка с низкой скоростью кислотообразования в молоке предельная кислотность в молоке была меньше 80° Т.
В оптимальных по составу питательных средах главным фактором, лимитирующим рост сливочного лактококка, является активная кислотность. Так, урожай клеток лактококков в оптимальной среде с контролируемым pH достигает (З-б)ТО10 /мл (7-8 мг сухого веса/мл), а в этой же среде с неконтролируемым pH он равен 1,5 107мл или ~ 0,5 мг сухого веса/мл. Сухой вес сливочного лактококка в pH-стате при pH 6,3 и прочих оптимальных условиях достигает 10 мг/мл. Оптимальный для скорости роста в полноценной среде pH 6,3-6,9, при концентрации лактозы 37 г/л. Выходы кислоты и биомассы не зависят от pH в интервале 5,6-7,5 и концентрации лактозы 48,5-38 г/л. Сыры отличаются довольно высокой буферностью, поэтому представляет интерес действие недиссоцииро- ванной молочной кислоты на развитие лактококков. Предельная концентрация лактатов, при которой возможен их рост при прочих оптимальных условиях, в зависимости от штамма равна 5,0-7,0%. Сливочный лактококк не всегда рос в бульоне с содержанием лактатов больше 4%, другие подвиды росли при более высокой концентрации лактатов. Торможение развития лактококков и лейконостоков в питательной среде при прочих оптимальных условиях, по наблюдениям Перфильева и Рогова (1989), происходит при содержании больше 2% лактатов.
В сырах с 40% влаги содержание молочной кислоты в водной фазе, равное 5%, соответствует 2,1% кислоты в сыре. В зарубежных сырах голландской группы, Чеддере содержание лактатов примерно равно 1,5-1,8%, в отечественных - 1,5-2,1%, в сырах с колющейся консистенцией содержание кислоты может достигать 2,4-2,7%, или примерно 5,6-6,3% в водной фазе, в Швейцарском сыре - 1,35%. Более высокое содержание кислоты в отечественных сырах обусловлено меньшим количеством воды, добавляемой в сыворотку во время их выработки. Таким образом, содержание молочной кислоты в сырах с низкой температурой II нагревания, особенно отечественного производства, находится на уровне, при котором рост лактококков невозможен. При этом нужно учесть, что температура созревания сыров также неблагоприятна для их роста. Показано, что рост и кислотообразование микрофлоры мезофильных заквасок в значительной степени тормозится уже при pH меньше 5,7 и содержании лактатов больше 1%.
Таким образом, напрашивается вывод, что рост лактококков в сырах прекращается не после того, как в них перестанет обнаруживаться лактоза, как обычно считается, а раньше. Другими словами, лактококки не всегда могут сбродить всю лактозу, остающуюся в сырной массе. В экстракте трехмесячного сыра Манчего лактококки не росли даже после обогащения его дрожжевым экстрактом и лактозой и внесении в экстракт каталазы для дезактивации токсичных радикалов 02, которые могли образоваться во время созревания сыра. Скорее всего рост лактококков в экстрактах был ингибирован или подавлен комбинацией низкого pH, высокой концентрации недиссоциированной кислоты, а в сырах также низкими температурами созревания. Каждый этот фактор в сыре недостаточен для подавления развития лактококков, а при совместном действии они усиливают действие друг друга.
Увеличение биомассы лактобацилл прекращается позднее, чем у лактококков, из-за более высокой их устойчивости к кислоте. Добавление в экстракт сыра лактозы возобновляет накопление биомассы Lbc. casei, следовательно, рост ее в сыре прекратился в связи с исчерпанием источников энергии.
Количество остающейся в сырной массе лактозы и молочной кислоты зависит от технологии сыра, в частности от степени разбавления сыворотки водой, скорости молочнокислого процесса в сырной ванне. Регулируя содержание этих компонентов в сырах после прессования, мы будем в определенной степени регулировать и уровень развития в сырах молочнокислых бактерий незаквасочного происхождения, в частности лактобацилл, педиококков, энтерококков.
После достижения максимума содержание жизнеспособных клеток лактококков в сырах начинает снижаться. Однако это снижение нельзя считать достаточным доказательством прекращения размножения лактококков, поскольку процесс размножения может идти параллельно с процессом отмирания клеток.
Лактококки не используют кислород в энергетическом обмене, но связывают большую часть растворенного в среде кислорода и тем самым снижают окислительно-восстановительный потенциал и создают условия, необходимые для своего размножения, поскольку несвязанный кислород ингибирует их развитие, особенно на лактозе. Справедливости ради нужно отметить, что небольшие концентрации кислорода в среде могут стимулировать рост лактококков: рост сливочного и молочного лактококков при 20° С и pH 5,05-5,45 в атмосфере азота в присутствии 0,5% 02 был на 17,5% выше, чем в отсутствие 02. Лактококки в молоке и синтетических средах обладают абсолютной потребностью в С02.
При посевной дозе 1% и температуре 30° С лактококки за 8-10 ч инкубации поглощали от 30,6% (активный диацетильный лактококк) до 77,2% (сливочный лактококк) растворенного в молоке кислорода, его концентрация снижалась до 2,5-5,9 мкг/мл. Поглощение кислорода сопровождается накоплением в молоке 0,03-0,19 ммоль Н202, которая затем расщепляется без образования кислорода с помощью NADH-перок- сидазы. За короткий срок окислительно-восстановительный потенциал в культурах лактококков снижался до -(250-410) мВ. Поглощение кислорода и снижение Eh лактококками играет большую роль в сыроделии, так как это создает необходимые условия для биохимических процессов, лежащих в основе созревания сыра, и делает невозможным размножение строгих аэробов в сырной массе.
Молочный лактококк широко распространен в природе благодаря высокой метаболитической активности и устойчивости к внешним факторам, что облегчает пополнение коллекции производственных штаммов лактококков.
Сгусток молока, образованный молочным лактококком, обладает более высокой синеретической способностью, чем образованный сливочным лактококком. В опытах Сухоцкене сгустки с очень хорошей, хорошей и удовлетворительной синеретической способностью давали соответственно 44, 50 и 6% штаммов молочного и 8, 58 и 34% штаммов сливочного лактококка.
Сгусток, образованный сычужным энзимом в кооперации со сливочным лактококком, медленнее отделяет сыворотку, что замедляет обсушку зерна и увеличивает влажность и выход сыра, ускоряет его созревание и улучшает консистенцию, но также может быть причиной более интенсивного роста посторонней микрофлоры и излишне кислого вкуса сыра.
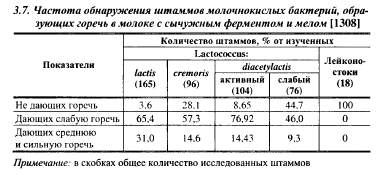
Одной из причин появления горечи в сырах могут быть специфические особенности расщепления казеина входящими в закваску штаммами молочнокислых бактерий. В табл. 3.7. показана частота обнаружения штаммов лакококков и лейконостоков, образующих горечь в молоке. По частоте обнаружения штаммов, способных накапливать горькие продукты в молоке с сычужным энзимом и мелом, лактококки располагались в следующем порядке: молочный > диацетильный активный > сливочный > диаце- тильный слабый. Штаммы, не дающие горечь в молоке, среди штаммов сливочного лактококка встречались в 7,8 раза чаще, чем среди штаммов молочного лактококка. Как правило, штаммы молочного лактококка, обладающие достаточно высокими скоростями роста и кислотообразования при повышенных температурах, образовывали среднюю и сильную горечь в молоке после 7 сут инкубации при 30° С (образование горечи в молоке коррелирует с ее образованием в сыре). Штаммы с низкой скоростью кислотообразования в молоке чаще всего не дают горечи, в закваски для производства сыров целесообразно вносить комбинации «горьких» и «негорьких» штаммов, которые обеспечивают достаточную скорость кислотообразования и не дают в сырах горечи. Таким образом, в отношении образования горьких продуктов сливочный лактококк имеет несомненное преимущество перед молочным, что подтверждается многими авторами.
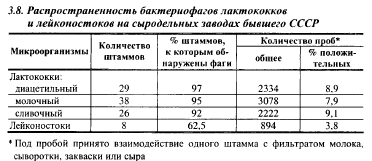
Имеются сообщения о более высокой специфичности сливочного лактококка к бактериофагам. Однако исследования видовой и штаммовой специфичности бактериофагов к микрофлоре мезофильных заквасок, выделенных на 13 сыродельных заводах бывшего СССР, расположенных в различных географических зонах, это не подтвердили (табл. 3.8). Были обнаружены бактериофаги к большинству штаммов лактококков, использованных в качестве тест-культур в этих исследованиях; частота обнаруживаемости бактериофагов по отношению к различным подвидам лактококков была почти одинаковой. Бактериофаги к двум штаммам молочного (5%) и двум штаммам сливочного (8%) лактококков не были найдены.
Некоторые штаммы молочного лактококка синтезируют антибиотик низин, обладающий достаточно широким спектром антибактериального действия по отношению к посторонней микрофлоре сыров. К сожалению, эти штаммы пока не нашли широкого применения в сыроделии, поскольку они обладают низкой кислотообразующей активностью и высокой чувствительностью к бактериофагам, а сам низин ингибирует рост активных кислотообразователей закваски и пропионовокислых бактерий.
Штаммы сливочного и молочного лактококков, образующие другой антибиотик - диплококкцин (типа бактериоцинов), также не нашли достаточно широкого применения из-за узкого спектра действия этого антибиотика, направленного в основном на другие виды и штаммы молочнокислых бактерий. Некоторые штаммы лактококков образуют бактериоцины с широким спектром действия против грамположительных и грамотрицательных бактерий.
Некоторые штаммы Lc. lactis образуют в сырах фруктовый и солодовый привкус, сливочный лактококк такой способностью не обладает.
Из анализа недостатков и достоинств молочного и сливочного лактококков нельзя сделать однозначный вывод о том, какой подвид следует включать в закваски для сыров с низкими температурами II нагревания.
Выработки сыров, проведенные в экспериментальном цехе ВНИИМС, показали, что сыры высокого качества можно вырабатывать на угличской закваске, в которой доминируют молочный и диацетильный лактококки, и зарубежных заквасках, в которых лактококки представлены сливочным и диацетильным лактококками с низкой кислотообразующей активностью, хотя в некоторых случаях сыры с зарубежными заквасками оценивались немного выше. В производственных условиях, наоборот, сыры с заквасками, в которых преобладали штаммы молочного лактококка, как правило, оценивались выше. Закваски, в которых кислотообразующие микроорганизмы представлены сливочным лактококком, требуют более высокого качества молока и санитарно-гигиенического уровня производства и соответствующей корректировки технологии. Некоторые авторы считают, что лучшие результаты дает закваска, в состав которой входят сливочный и молочный лактококки. Этот принцип соблюдается в подборе микрофлоры угличских заквасок, но из-за различных скоростей роста сливочного, молочного и диацетильного лактококков, особенно во время II нагревания и какое-то время после него, в сырах, выработанных на этих заквасках, доминируют последние два подвида. Litnsowtin считает целесообразным использовать в составе закваски для производства сыра Чеддер сливочный и молочный лактококки в отношении 2:1 [648]. Продолжительность выработки сыра Чеддер с комбинированными культурами этих подвидов сокращалась на 7,5-12,5%.
