Протеолиз в мелких сычужных сырах
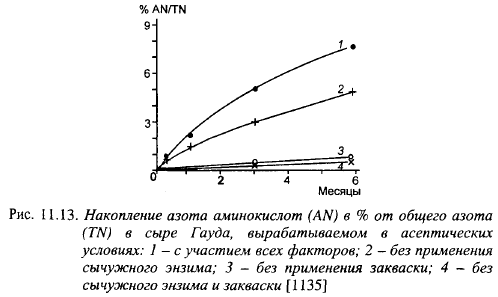
Протеолиз - основополагающий процесс созревания сыров. Мелкие сычужные сыры отличаются меньшей шириной и глубиной протеолиза (табл. 11.7). Количество аминного азота в % от растворимого в мелких сырах (Сен-Полен) было в 2,06 раза меньше, чем в Грюйере. В отечественных мелких сырах, созревающих 60 сут, количество водорастворимого азота составляет 20-25%, остальной азот представлен негидролизованными белками или белками с очень низкой степенью гидролиза, в т. ч. пара-æ-казеином.
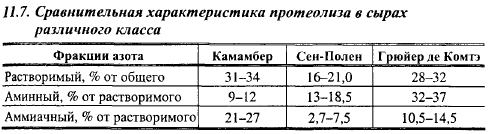
Характер протеолиза в сырах с низкими температурами II нагревания изучен на сыре Гауда, выработанном в асептических условиях, позволяющих исключить тот или иной фактор из производства и выявить его роль в протеолизе. Накопление продуктов протеолиза в сырах, выработанных в асептических условиях, под действием отдельных главных факторов и их комбинаций показано в табл. 11.3, на рис. 11.12 и 11.13. В сырах, выработанных с реннином и лакто кокковой закваской, аs1казеин практически полностью расщеплялся в течение месяца, в то время как 50% β-казеина не было расщеплено в течение 6 мес. Расщепление β-казеина с заметной скоростью началось спустя месяц после выработки. В сырах без закваски с доведением pH сгустка с помощью глюко- но-5-лактона (в результате гидролиза которого кислотность сыра медленно повышается, имитируя повышение кислотности и конечный pH, вызываемый развитием микрофлоры закваски) количество водорастворимого азота было намного ниже, чем в сырах с комплексом факторов; оно нарастало в течение 6 мес созревания пропорционально дозе вносимого реннина. В сырах, выработанных без закваски, в 1-, 2- и 6-месячном возрасте содержание азота свободных аминокислот составило соответственно 0,1; 0,2 и 0,3% от общего содержания азота в сыре, а в сыре с заквасками, но без реннина - соответственно 1,3; 2,3 и 4,0% (табл. 11.3). Таким образом, исключение заквасок уменьшило содержание свободных аминокислот в сыре примерно в 13 раз. В сырах без заквасок содержание свободных аминокислот не зависело от дозы реннина.
В сырах без реннина очень сильно уменьшилась скорость гидролиза сщ-казеина и накопление водорастворимого при pH 4,6 азота. В сыре Гауда, выработанном без реннина, количество водорастворимого азота, образованного за счет протеолитических систем закваски (предполагая, что активность молочных протеиназ в сырах обоих вариантов была одинаковой), в 1-, 3- и 6-месячном возрасте составило 10,6; 12,7 и 19,5% от их количества в сырах, выработанных с реннином, но без закваски. Исключение реннина сильно снизило количество свободных аминокислот, хотя в сырах без закваски реннин практически не образовывал свободных аминокислот. Объяснить это можно тем, что лактококки значительную часть свободных аминокислот образуют из продуктов протеолиза казеина реннином.
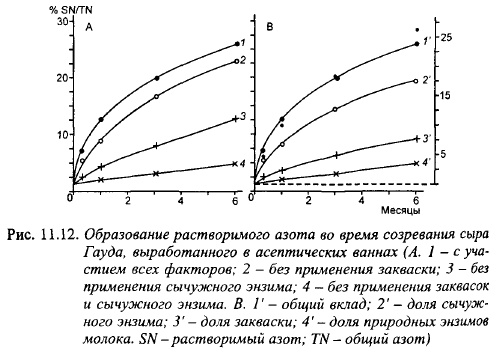
В сырах без реннина и закваски наблюдалось незначительное расщепление β казеина и очень небольшой гидролиз asl- и а52-казеинов с образованием за 6 мес созревания около 4% растворимого азота и 0,4% азота свободных аминокислот от общего содержания азота, что вызвано действием плазмина. Плазмин не влиял на протеолитическую активность реннина и закваски.
Количество водорастворимого азота в сырах голландской группы почти линейно зависит от дозы реннина, количество свободных аминокислот от дозы энзима практически не зависит.
Таким образом, реннин расщепляет казенны с образованием крупных, средних и мелких пептидов, т. е. в ширину, микрофлора закваски - с образованием низкомолекулярных пептидов и, особенно, свободных аминокислот, т. е. в глубину, протеиназа молока расщепляет казенны с образованием низкомолекулярных пептидов.
Основное расщепление asl-казеина в сырах с низкими температурами II нагревания происходит под действием молокосвертывающих энзимов, остающихся в сырной массе; протеолитическая активность микрофлоры заквасок становится заметной через месяц созревания, очевидно, после начала лизиса бактериальных клеток и высвобождения внутриклеточных энзимов. Протеолитическая активность микрофлоры заквасок для мелких сыров, связана с расщеплением β-казеина (а-казеин к этому времени почти весь гидролизован реннином) и пептидов, в том числе образующихся в результате гидролиза казеина молокосвертывающими энзимами. В целом протеолиз в мелких сырах является результатом синергетического действия молокосвертывающих энзимов и микрофлоры закваски.
Для проверки влияния состава заквасок на протеолиз в асептических условиях вырабатывали сыры с семью различными заквасками при примерно одинаковом количестве реннина, остающегося в сгустке. В 3-месячных сырах в зависимости от вида закваски содержание азота свободных аминокислот составляло (в % от общего азота) от 3 до 5,2%, в 6-месячных - от 3,8 до 9,3%; количество водорастворимого азота во всех вариантах было почти одинаковым (от 17 до 20% в 3-месячных сырах).
Выработки сыров в асептических условиях показывают, что для формирования типичных органолептических показателей сыров с низкими температурами II нагревания, включая группу Чеддера, достаточно молокосвертывающих энзимов и лактококковых заквасок.
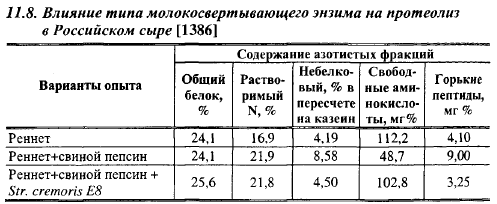
Протеолиз в Российском сыре, выработанном в обычной ванне, и влияние на него типа молокосвертывающего энзима показаны в табл. 11.8. Сыры вырабатывали на угличской закваске; в третьем варианте в молоко вносили культуральную среду (без клеток) «негорького» штамма Е8 в количестве 30% от дозы закваски, использованной в 1 и 2 вариантах, с соответствующим снижением дозы закваски. Соотношение в смеси реннета и пепсина равнялось 50:50. Замена сычужного порошка смесью его со свиным пепсином увеличило в сыре содержание водорастворимого и небелкового азота, но уменьшило содержание свободных аминокислот. По- видимому, пептиды, образованные свиным пепсином, медленнее расщеплялись микрофлорой закваски до свободных аминокислот, чем пептиды, образованные реннином. По количеству свободных аминокислот, образуемых в основном под действием закваски, сыры Гауда не отличались от Российского сыра. Внесение культуральной среды «негорького» штамма сливочного лактококка увеличило содержание в сыре свободных аминокислот и уменьшило содержание «горьких» пептидов.
Количество водорастворимого и небелкового азота в сыре Чеддер, вырабатываемом с участием плесневых молокосвертывающих энзимов, растет быстрее, чем с реннином, что, очевидно, обусловлено их более высоким неспецифическим протеолизом, в частности способностью атаковать в сыре не только as -, но и β-казеин.
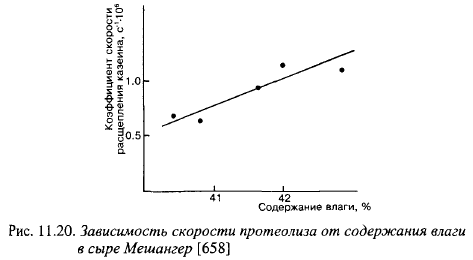
На протеолиз огромное влияние оказывает состав и физико-химические показатели сырной массы. Чем ниже отношение содержания влаги к содержанию казеина (или СОМО), тем медленнее идет протеолиз (рис. 11.20). Даже небольшое снижение Ав понижает количество молокосвертывающих энзимов, остающихся в сырной массе, по отношению к содержанию казеина, в результате чего уменьшается скорость протеолиза.
Чем выше отношение содержания соли к содержанию влаги, тем медленнее идет протеолиз и более твердой будет консистенция сыра. Особенно чувствительно к содержанию соли расщепление β-казеина (разд. 10.6). Протеолиз в сырах идет с достаточной скоростью, если концентрация соли в водной фазе сыров лежит в интервале от 4,5 до 5,7%. При более низкой концентрации соли интенсивность протеолиза увеличивается, что сопряжено с появлением в сыре пастообразной, мажущейся консистенции, горечи. При одной и той же концентрации соли повышение содержания в сыре Са замедляет протеолиз, однако высокое содержание Са обычно бывает только в сырах с повышенным pH, а pH сильнее влияет на скорость протеолиза, чем Са (с повышением pH скорость протеолиза возрастает).
Скорость протеолиза не единственный критерий при оценке режима выработки сыра, так как увеличение pH и содержания влаги, снижение содержания соли не только ускоряют протеолиз, но и создают благоприятные условия для развития посторонней микрофлоры в сыре. Поэтому выработку сыра с оптимальными параметрами можно проводить только при хорошем качестве молока, стабильной и достаточно высокой активности заквасок и высоком уровне гигиены производства заквасок.
Анализ действия физико-химических факторов на протеолиз позволяет объяснить небольшие масштабы протеолиза в Чеддере (даже в старых сырах этого вида содержание азота свободных аминокислот не превышает 3% от общего содержания азота). Причинами, очевидно, являются низкий pH и равномерное распределение соли в головке сразу же после выработки.
Особенности протеолиза в сырах с высокой и средней температурой II нагревания
Движущие силы протеолиза в крупных и мелких сырах неодинаковы. Высокие температуры II нагревания в крупных сырах инактивируют молокосвертывающие энзимы и резко ограничивают развитие лактококков, которые играют главную роль в протеолизе в мелких сырах. Значительная часть крупных сыров вырабатывается из сырого или тер- мизованного молока, поэтому в них возрастает роль плазмина, резистентного к температурам II нагревания. Плазмин в большем количестве, чем сычужный энзим, переходит из молока в сыр.
Основная микрофлора заквасок для крупных сыров - термофильные стрептококк и лактобациллы, действие которых на белки молока отличается от действия лактококков (11.2.4). Нельзя исключить и влияние на протеолиз в крупных сырах мезофильных лактобацилл. Неотъемлемой микрофлорой крупных сыров, за исключением терочных, являются пропионовокислые бактерии, обладающие специфической, хотя и небольшой протеолитической активностью.
Для выявления роли плазмина в крупных сырах готовили смесь молокосвертывающего энзима (Реннилазы), энзимного препарата из Lbc. helveticus (протеолитические энзимы закваски) и плазмина и добавляли в обезжиренное или пастеризованное молоко, обогащенное Са2+. После выдержки анализировали продукты протеолиза. Только смеси, в которые входил плазмин, давали электрофореограммы, подобные полученным при анализе крупных сыров. Реннилаза в основном гидролизовала γ казеин и протеозо-пептонные фракции, получаемые в результате действия плазмина на β-казеин. Плазмин слабо атакует α-казеин, гидролиз которого очень важен в формировании консистенции в мелких сырах, что несомненно является одной из причин различий в консистенции крупных и мелких сыров. Низкая активность плазмина в некоторых крупных сырах вызывала появление постороннего привкуса. Проведенные исследования показывают решающую роль плазмина в гидролизе β-казеина в крупных сырах.
В отличие от лактококков, лактобациллы, используемые в заквасках для крупных сыров, обладают более высокой протеолитической активностью и атакуют все казенны, хотя их протеиназная активность зависит от вида и штамма (разд. 11.2.4). Закваски с Lbc. helveticus более чувствительны к соли, чем закваски с Lbc. lactis.
Закваски для крупных сыров больше накапливали низкомолекулярных продуктов расщепления казеина, чем лактококки. Важно соотношение между видами бактерий в заквасках. Повышение дозы Lbc. bulgaricus в интервале от 0,5 до 1000 мл /200 кг молока, например, увеличивает образование пролина, но угнетает развитие пропионовокислых бактерий.
Активность внутриклеточных и внеклеточной протеиназ термофильного стрептококка невелика и зависит от штамма, хотя некоторые штаммы расщепляют все казенны.
Следует иметь в виду, что мезофильные лактобациллы играют более важную роль в крупных сырах, чем в мелких, а они обладают высокой протеолитической активностью в сравнении с другими лактобактериями.
Таким образом, в крупных сырах принимают участие в протеолизе энзимы из различных источников, что должно сопровождаться большей глубиной расщепления белков и разнообразием продуктов протеолиза. Пониженная влажность этих сыров, безусловно, замедляет протеолиз, но более низкое содержание в них соли, выдержка определенное время при повышенных температурах, длительное созревание, более высокий уровень pH, обусловленный пониженной влажностью и более низким содержанием лактозы, ферментацией части лактатов пропионовокислыми бактериями, не только компенсируют негативное влияние низкой влажности на протеолиз, но обеспечивают большую интенсивность его в крупных сырах. Об этом свидетельствует динамика протеолиза в Эмментальском сыре, показанная в табл. 11.9. В 5-месячном Эмментальском сыре азот свободных аминокислот составил 40% от общего содержания азота в сыре.
Протеолиз и формирование консистенции
Казалось бы, с влиянием протеолиза на консистенцию сыра все ясно: протеолиз постепенно разрушает казеиновый каркас, являющийся основой структуры сырного сгустка, поэтому по мере созревания прочность и эластичность сгустка должны снижаться, а консистенция сыра становиться более нежной и пластичной. На самом деле консистенция сыра зависит не только от протеолиза, но и от других факторов, что усложняет процесс ее формирования, раскрытый далеко не полностью. Белоусов еще в начале 50-х годов установил, что разрушение образцов сыра по мере созревания происходит при все меньшей относительной деформации. Относительную деформацию, при которой происходит разрушение образца, Белоусов назвал критической. Критическая деформация характеризует связность сырной массы. Следовательно, по мере созревания связность сырной массы снижается, а хрупкость увеличивается.
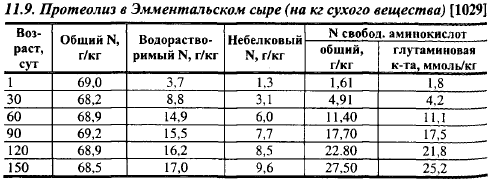
Наблюдения Белоусова хорошо согласуются с результатами более поздних опытов Неберта с сотр., изучавшими изменения пенетрации Костромского сыра в процессе созревания (рис. 11.14; 11.15). На первых стадиях созревания реннин гидролизует as1-казеин всего по нескольким связям, но даже отщепление одного asr I пептида достаточно для значительного изменения реологических показателей мелких сыров. На рис. 11.14 видно, что в течение 10 сут после прессования величина пенетрации снизилась по сравнению с ее величиной в сыре после прессования примерно на 24%. Показатели пенетрации не переводятся в известные физические величины. Однако существует статистически достоверная корреляция между пенетрацией и твердостью сыра. Взаимосвязь между пенетрацией и твердостью на примере Швейцарского сыра показана на рис. 11.16. Чем тверже консистенция, тем ниже показатели пенетрации, хотя связь между ними не является линейной.
После 10 сут показатели пенетрации в Костромском сыре, хотя и с меньшей скоростью, продолжают снижаться до 45-суточного возраста параллельно с уменьшением влажности сыра (табл. 11.10). По Табачникову и Тетеревой, коэффициент корреляции между показателем пенетрации и содержанием влаги в сыре равен 0,89±0,02. Затем показатели пенетрации увеличиваются и к 60-суточному возрасту они возвращаются к уровню 30-суточного сыра, после чего до 90-суточного возраста (время наблюдения) очень медленно увеличиваются. Таким образом, твердость Костромского сыра увеличивается в течение 45 сут созревания, а не снижается, как казалось бы, должно происходить в связи с протеолизом.
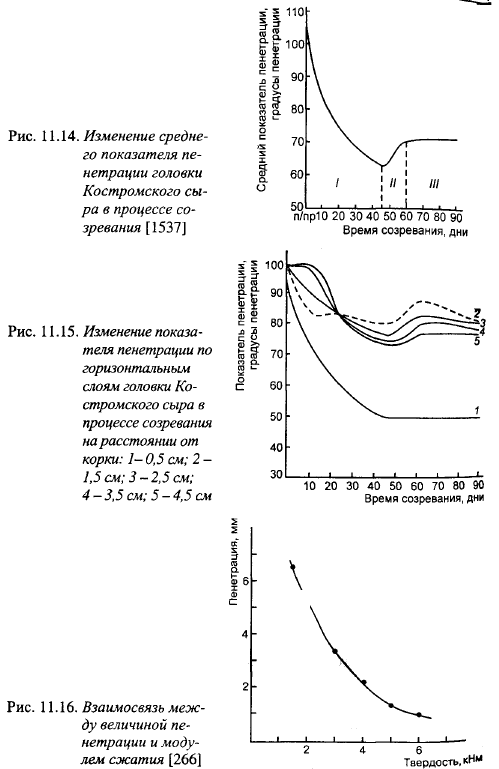

Существует отрицательная корреляция между твердостью и упругостью сыра, с одной стороны, и протеолизом, с другой. Реологические показатели Костромского сыра в процессе созревания изменяются в различных слоях головки по-разному: наибольшее снижение показателей пенетрации наблюдается в слое, расположенном на расстоянии 0,5 см от поверхности, а в более удаленных слоях при одинаковой тенденции масштабы изменений уменьшаются (рис. 11.15). Степень неоднородности сырного теста возрастает с увеличением протеолиза.
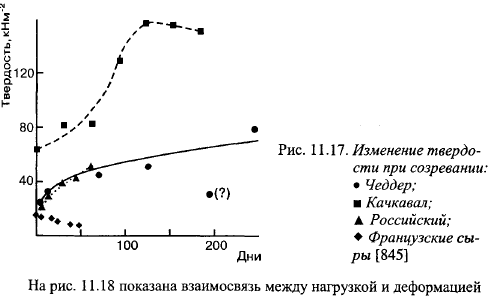
На рис. 11.17 показано изменение твердости трех других видов сыров, которые подтверждают результаты опытов Неберта с соавт. об изменении твердости Костромского сыра.
На рис. 11.18 показана взаимосвязь между нагрузкой и деформацией молодых и зрелых сыров в пробе на сжатие. Из него видно, что в зрелых сырах уменьшается деформация, при которой происходит разрушение образца; усилие, при котором разрушается образец при сжатии, в зрелом сыре Гауда намного выше, чем в молодом, а в сыре Чеддер - наоборот. Снижение деформации сыра в момент достижения предела текучести показано на рис. 11.19. Упругость сыров снижается во время созревания.
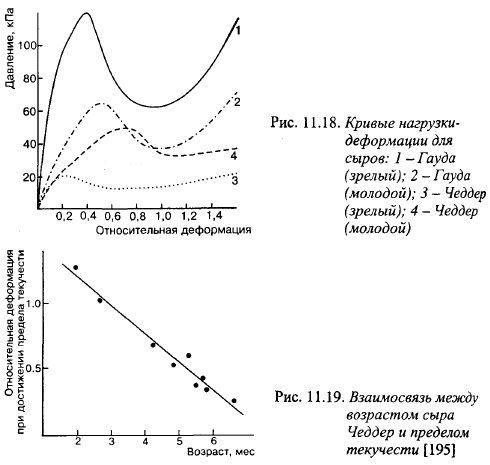
Величина уменьшения деформации, при которой начинает разрушаться образец, является единственным реологическим показателем, коррелирующим со степенью зрелости мелких сыров. Это свидетельствует о снижении связности сырной массы по мере созревания. Снижение связности сырной массы в процессе созревания можно считать логическим следствием протеолиза и уменьшения активности воды. В то же время при органолептической оценке сырная масса по мере созревания становится более однородной, хорошо расходящейся во рту.
Неберт и др. и Lawrence et al. объясняют характер изменений реологических показателей мелких сыров при созревании тем, что в созревающем сыре идут два процесса: протеолиз, снижающий прочность казеинового каркаса и размягчающий консистенцию, и потеря сгустком влаги, которая особенно велика на начальных этапах созревания и в поверхностном слое, в результате чего твердость сыра возрастает. Суммарный результат на каждом этапе и в каждом слое зависит от соотношения между этими процессами. Неодинаковый характер изменения реологических показателей по слоям сыра они связывают с миграцией соли и влаги в головке. К 45-суточному возрасту миграция практически прекращается, а дальнейшее снижение твердости и повышение пластичности сыра, очевидно, происходит главным образом под действием протеолиза. К 60-суточному возрасту заканчивается расщепление as1-казеина, обусловливающего прочность казеинового каркаса сычужного сгустка, и величина пенетрации стабилизируется. Если это так, то продолжительность созревания мелких сыров, вырабатываемых по традиционной технологии, должна составлять 45-60 сут.
Таким образом, максимальные изменения показателей пенетрации, а следовательно, и твердости отмечаются в поверхностных слоях сыра, в которых наиболее сильно снижается активность воды из-за потери влаги и поступления большого количества соли из рассола. Однако, различия в консистенции внутренних и внешних слоев сыра могут быть связаны также со снижением скорости протеолиза, которая в сильной степени зависит от этих факторов (рис. 11.20). Следует отметить, что во время созревания сыра Чеддер, в котором в связи с особенностями посолки (разд. 10.3.3) миграция влаги и соли в пределах головки незначительна, его твердость также увеличивалась, но медленнее, чем в Костромском сыре (рис. 11.17). Кривые увеличения твердости сыров Чеддера и Российского во время созревания почти совпадают, хотя Российский сыр солят в рассоле после прессования. Это свидетельствует о том, что не только миграция соли и влаги являются причиной повышения твердости сыров во время созревания.
Активность воды во время созревания также снижается в результате протеолиза, так как расщепление каждой пептидной связи ведет к появлению двух новых ионных групп, связывающих воду и таким образом снижающих количество доступной для сольватации белков воды в системе. Сырная масса становится более твердой и более ломкой, т. е. разрушающейся при повышенных нагрузках, но малых деформациях. На первом этапе, когда величина пенетрации в Костромском сыре снижается особенно быстро, активность воды в сыре уменьшается также в результате сбраживания лактозы, поскольку образующиеся при этом кислоты обладают гораздо большей водосвязывающей активностью, чем исходная лактоза.
В сырах Чеддер, отличающихся низким содержанием влаги (36-39%), консистенция остается ломкой, несмотря на продолжительное созревание.
При определенном снижении Ав в системе консистенция становиться крошливой, что часто бывает в старых сырах Чеддер. Ведущую роль активности воды в формировании ломкой консистенции доказывают опыты с сыром Свесия: увеличение концентрации соли в водной фазе этого сыра с 1,55 до 4,5% (соль вносили в зерно после удаления сыворотки) привело к трансформации консистенции сыра из слегка ломкой - типичной для этого сыра - в более плотную, менее связную и гомогенную, несмотря на более высокую влажность сыров с повышенной концентрацией соли.
При высокой влажности консистенция сыра во время созревания размягчается пропорционально степени протеолиза, главным образом, аs1-казеина . Раманаускас показывает, что при высокой влажности Российского сыра в процессе созревания содержание в нем небелковых азотистых веществ возрастает, эластичность снижается, т. е. превалируют процессы, обусловливаемые расщеплением белков, а не снижением активности воды. В то же время интенсификация протеолиза в сыре Проволоне с низкой влажностью внесением в смесь термофильных лактобацилл, находящихся в состоянии теплового шока, приводит не к размягчению консистенции, а к ускорению формирования типичной для этого сыра ломкой консистенции. В этом случае большее влияние на консистенцию оказывает изменение Ав. Возможно, потеря сыром во время созревания части влаги сближает параказеиновые пряди, лежащие на поверхности жировых шариков и образующие ячеистую структуру, и между ними возникают новые физические или химические связи.
Сканирующая электронная микроскопия показала, что сырная масса представляет непрерывную белковую матрицу, структурные элементы которой в различных видах сыров имеют существенные различия (рис. 11.21). Структурные единицы белковой матрицы в сыре Гауда имеют такую же глобулярную форму (10-15 нм в диаметре), как в исходном молоке, а в сыре Чешир они намного меньше (3—4 нм) и имеют вид цепочек или прядей. Чеддер занимает промежуточное положение: часть его структурных единиц имеет глобулярную форму, как в сыре Гауда, часть - такую же форму, как в сыре Чешир.
Изменение структуры сыров коррелирует с pH сыров. По уровню pH сыры располагаются следующим образом: Стильтон (pH 4,6-4,75) < < Чешир (4,75-4,9) < Чеддер (4,9-5,3) < Российский (5,1-5,3) < Костромской, Гауда (5,2-5,35). Чем ближе pH сыра к изоэлектрической точке параказеина, тем более компактную форму принимают белки, тем при меньшей деформации сжатия разрушается структура. Уменьшение размеров структурных единиц казеинового каркаса при понижении pH сыра может быть вызвано понижением степени гидратации белков. Активная кислотность может влиять на консистенцию через протеолиз, так как чем ниже pH сыров, тем менее интенсивно идет расщепление белков.
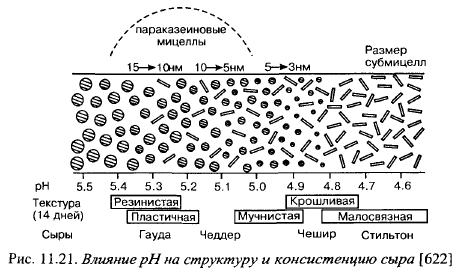
Однако в этом случае замедлялся бы переход консистенции от резинистой к более хрупкой, а для сыров с низким pH характерными пороками консистенции как раз являются крошливость, ломкость, малосвязность, самокол. В гл. 2 показано, что слишком быстрое нарастание кислотности во время выработки сыра влияет на переход мицеллярного фосфата кальция в сыворотку, что оказывает большое влияние на консистенцию сыров.
Консистенция сыра становится менее твердой при уменьшении содержания кальция. В мелких сырах диапазон содержания Са выше, чем в сырах группы Чеддер, что повышает его роль в формировании консистенции.
На формирование консистенции сыров большое влияние оказывает температура созревания сыра. Так, консистенция 8-недельного сыра Чеддер, созревающего при 15° С, была такой же, как консистенция 16-недельного сыра этой же выработки, созревающего при 8° С. Повышение температуры созревания до 20° С привело к формированию хрупкой и крошливой консистенции.
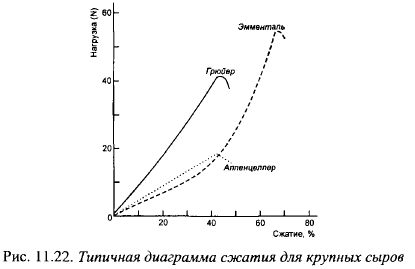
Консистенция сыров с высокими температурами II нагревания существенно отличается от консистенции мелких сыров. На рис. 11.22 показано сопротивление сжатию распространенных в Швейцарии сыров с высокими температурами II нагревания. Эта диаграмма позволяет определить усилие, которое необходимо приложить для разрушения образца, и высоту образца в момент разрушения в % от первоначальной.
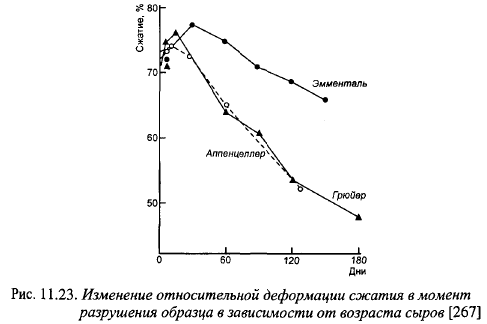
Степень сжатия в момент разрушения образца свидетельствует об однородности, связности консистенции: разрушение при низкой компрессии свидетельствует о крошливости, малой связности консистенции, и наоборот. Усилие, необходимое для сжатия образца на одну треть, используют для характеристики твердости крупных сыров. Величина усилия, разрушающего образец при сжатии, зависит от твердости и характера консистенции сыра. На рис. 11.23 показано изменение степени сжатия в момент разрушения образца в процессе созревания. Степень сжатия образцов Эмментальского сыра и усилие в момент разрушения значительно выше, чем сыров Грюйер и Аппенцеллер, что свидетельствует о большей гомогенности, связности его консистенции и более ломкой консистенции Грюйера и Аппенцеллера.
Связность консистенции Эмменталя увеличивается в первые 30 сут созревания, затем она начинает снижаться, консистенция становится более рыхлой. Повышение связности консистенции сыров Грюйер и Аппенцеллер в первый период созревания (примерно до окончания образования глазков) менее выражено, а скорость ее снижения в последующие периоды больше.
Различия в реологических показателях между Эмменталем и двумя другими видами крупных сыров главным образом обусловлены более сильным протеолизом в двух последних видах. Существует обратная корреляция между степенью сжатия образца сыра в момент разрушения и содержанием в сыре небелкового азота. Более интенсивный протеолиз в швейцарских сырах Грюйер и Аппенцеллер обусловлен тем, что в их созревании принимает участие микрофлора поверхностной слизи. В зрелых сырах Грюйер и Аппенцеллер содержание небелкового азота было выше, чем в Эмментальском сыре, в 1,38 и 1,17 раза соответственно. Предельная степень сжатия уменьшается с повышением температуры созревания, что также можно объяснить повышением скорости протеолиза.
Даже небольшие изменения в характере протеолиза могут оказать влияние на консистенцию сыра. Так, консистенция сыров с различными молокосвертывающими энзимами всегда отличается от консистенции сыров с реннетом, хотя и остается в рамках, приемлемых для потребителя. Правда, это обусловлено не только различиями в протеолизе во время созревания, но и неодинаковой структурой сгустков, получаемых в процессе свертывания.
Протеолиз и формирование вкуса и аромата сыра
Существует тесная корреляция между характером протеолиза, вкусом и ароматом сыра. При этом скорость формирования и степень выраженности сырного вкуса коррелирует с расщеплением только β-казеина, которое осуществляют в мелких сырах микрофлора закваски, лактобактерии незаквасочного происхождения, плазмин. В формировании органолептических показателей сыров принимают участие продукты протеолиза - пептиды и свободные аминокислоты (САК), а также соединения, получающиеся в результате дальнейшего преобразования САК в результате воздействия на них микрофлоры сыра и чисто химических реакций (рис. 11.1). Кроме этого, протеолиз понижает степень связывания вкусовых и ароматических веществ сгустком и создает условия для их воздействия на органы чувств потребителей.
Образование горького вкуса. О влиянии пептидов - промежуточных продуктов протеолиза - на органолептические показатели сыров мало что известно, кроме того, что гидролиз казеинов может сопровождаться образованием пептидов, придающих горечь продукту. Горечь в сырах является одним из наиболее распространенных пороков. Горькие пептиды, образующиеся в сыре, чаще всего имеют молекулярную массу меньше 1400 и отличаются большой гидрофобностью. Champion & Stanley выделили горькую фракцию из Чеддера, содержащую большие количества лейцина и валина. Горькие фрагменты asl -казеина и β-казеина содержали много гидрофобных концов (фени- лаланил, изолейцил, лейцил, валил, пролил) и имели длину цепочек от 2 до 22 аминокислот. Синтезированы пептиды Арг-Глу-Про-Фен-Про-Иле- Иле-Вал, присутствующий в гидролизате β-казеина, и Арг-Глу-Про- Про-Фен-Иле-Вал, горечь которых обусловлена N-терминальным аргинином и гидрофобным пептидом в С-позиции. Пептиды отличаются необычайно горьким вкусом (вкусовой порог 0,004 и 0,05 мМ). Выделен горький пептид из β-казеина (196-209) со вкусовым порогом 0,015 мМ, горечь которого в 67 раз сильнее, чем горечь кофеина. Горьким вкусом обладал пептид из β-казеина, включающий остатки аминокислот в позиции 53-79.
Горькие пептиды в сырах образуют молокосвертывающие энзимы и лактококки. В опытах с растворами казеинов при pH 5,4 и 15° С реннет и химозин образовывали горькие пептиды из цельного казеина, asrказеина, β-казеина, пара-æ казеина; в сырах химозин не расщепляет β казеин.
Свиной пепсин образовывал горькие пептиды из С-концевой части а51-казеина; практически все гидрофобные пептиды были образованы в результате разрыва пепсином связей Лей, Тир и Фен. Наиболее горькими из гидролизатов казеинов, полученных под воздействием реннета, свиного пепсина, химотрипсина, трипсина, были гидролизаты а51-казеина.
Неразрушенные клетки Lc. cremoris HP, дающего горечь в сырах, образовывали горькие пептиды только из æ- и β-казеинов. Горькие пептиды образовывались под действием протеиназ клеточной стенки. Протеиназо-негативные варианты «горьких» штаммов лактококков, полученные генетическими методами, перестают образовывать горечь в сырах. Штаммы Lc. cremoris, не дающие горечь в сырах, расщепляют горькие пептиды, образуемые химозином из as1- и пара-æ-казеина, но не из β казеина. Расщепляют горькие пептиды внутриклеточные энзимы лактококков.
Горькие пептиды есть в негорьких сырах, но в низких концентрациях. В сырах Гауда главными компонентами горечи являются фрагменты β-казеина, что, по-видимому, обусловлено расщеплением большей части горьких пептидов из a-казеина лактококками закваски. По количеству образуемого водорастворимого азота в сыре различий между «горькими» и «негорькими» штаммами лактококков не обнаружено. Однако «негорькие» штаммы образуют значительно больше свободных аминокислот.
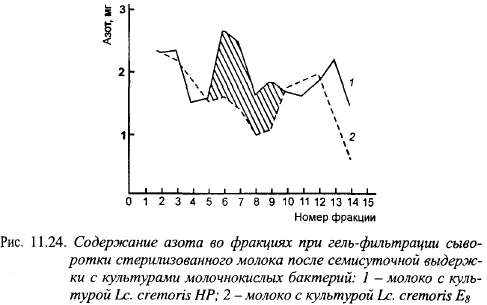
Элюционные профили сыров, выработанных с «горькими» и «негорькими» штаммами лактококков, сильно различаются (рис. 11.24.). В опыте использованы штамм HP, дающий, и штамм E8, не дающий горечи в сырах с низкой температурой II нагревания. Объем фракций 5-10 в варианте с «горьким» штаммом был значительно выше, чем в варианте с «негорьким» штаммом. В этих фракциях и сосредоточены горькие пептиды.
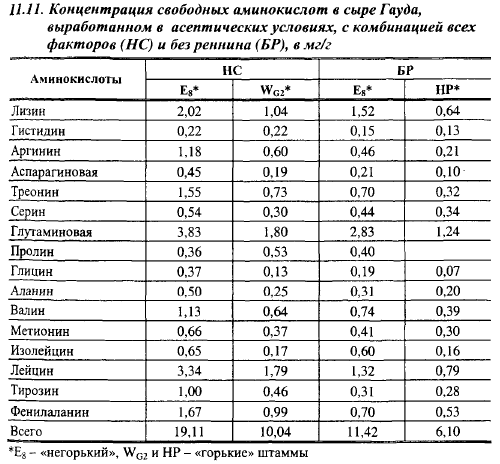
Есть различия в количестве и соотношении свободных аминокислот, образуемых «горькими» и «негорькими» штаммами Lc. cremoris в сыре Гауда (табл. 11.11). Содержание всех аминокислот было выше в сырах, выработанных с «негорьким» штаммом Е8. Содержание в них лизина было в два раза выше, чем в сырах с «горькими» штаммами, аргинина - в два раза, глутаминовой кислоты - в 2,13 раза, изолейцина - в 3,8 раза и т. д.
Эти различия наблюдались как в присутствии, так и в отсутствие молокосвертывающего энзима. Доля лейцина, серина, фенилаланина и пролина в общем содержании аминокислот и количество пептидов было выше у «горьких» штаммов.
Звягинцев и др. считают, что способность высвобождать большое количество глутаминовой кислоты можно использовать как критерий для отбора штаммов, не образующих горечь в сырах. Visser не считает эту пробу специфичной.
Несмотря на то что количество свободных аминокислот в сырах с «негорькими» штаммами всегда было выше, чем в сырах с «горькими» штаммами, соотношение между кислотами в них было близким.
Химозин высвобождает горькие пептиды в начале расщепления казеина. Из табл. 11.3 видно, что он образует в 2,6-3,5 раза больше пептидов с молекулярным весом менее 1400, чем закваски. Количество таких пептидов, образуемое при совместном действии реннина и лактококков, гораздо выше, чем их сумма при раздельном действии этих факторов. Возможно, это обусловлено тем, что лактококки сами не гидролизуют α-казеин в сыре, но способны расщеплять продукты его гидролиза реннином с образованием горьких пептидов. Примеси энзимов, содержащиеся в промышленных молокосвертывающих препаратах, образовывали больше горьких пептидов, чем химозин. Факторы, влияющие на количество остающихся в сыре молокосвертывающих энзимов (кислотность сыворотки и сырной массы в конце обработки зерна, промывка сгустка, температура II нагревания, доза энзимов), оказывают влияние на степень горечи сыров. Заменители сычужного энзима образуют больше горьких пептидов, чем химозин, при прочих одинаковых условиях. В Российском сыре со свиным пепсином, например, было обнаружено в 2,2 раза больше горьких пептидов, чем в сыре из этого же молока с реннином (табл. 11.8). Ионы Са стимулируют образование горьких пептидов.
Горькие пептиды из β-казеина, которые преобладают среди горьких пептидов в зрелых сырах, могут образовываться только микрофлорой закваски или посторонней микрофлорой.
По скорости накопления кислоты и биомассы штаммы лактококков делят на «быстрые» и «медленные». В сырах Чеддер горечь преимущественно вызывают «быстрые» штаммы лактококков, размножающиеся при температурах II нагревания, биомасса которых в сыре после прессования достигает уровня больше 109 КОЕ/г. «Медленные» штаммы, чувствительные к температуре II нагревания, биомасса которых в сырах после прессования находится на более низком уровне (примерно 1Сг КОЕ/г), горечи не вызывают. Горькие пептиды образуются под действием протеиназ клеточной стенки, а более низкая плотность популяции штамма в конце выработки уменьшает количество протеиназ. По данным Hillier et al., протеиназы «негорьких» штаммов не действуют при 40° С, но действуют при 30° С, протеиназы «горьких» штаммов действуют при обеих температурах.
Попытки выработать сыр на закваске только из «медленных» штаммов не принесли успеха; для формирования требуемого вкуса и аромата в закваске должно быть определенное количество «быстрых» штаммов. Mills & Thomas использовали для выработки сыра Чеддер закваску, 45-75% клеток в которой были протеиназо-отрицательными, т. е. относились к «медленным» штаммам, что позволило снизить уровень протеолитической активности при сохранении плотности популяции. В пяти из шести сыров, выработанных с этой закваской, горечь была менее выражена, чем в контрольных сырах.
В опытах Lowrie et al. штамм сливочного лактококка АМ2 в сырах, выработанных в асептических условиях при температуре II нагревания 38° С, имел невысокую концентрацию клеток в сыре перед прессованием и не дал горечи, а при температуре 33° С количество его клеток резко возросло и в сырах появилась горечь. Мезофильные лактобациллы снижают горечь в сырах.
Kempler et al. ликвидировали горечь в Чеддере, заменив в закваске «горький» штамм Lc. lactis С2 его трансдуктантом с пониженной про- теиназной активностью; Шуляк и Новикова предотвратили образование горечи в Костромском сыре, применив закваску с низкой про- теиназной активностью. По Stadhouders et al., для получения хорошего вкуса в мелких сыров в закваске достаточно иметь 20% протеи- назо-положительных штаммов.
При разработке технологии жидкого сыра Чеддер (при его выработке из молока удаляют Са) для ускорения формирования вкуса и запаха использовали нейтральную протеиназу из В. subtilis, но в продукте появилась горечь. Добавленный к нему пептидазный экстракт из «негорького» штамма молочного лактококка полностью ликвидировал горечь.
Снижение плотности популяции микрофлоры закваски, а следовательно, и протеиназной активности на начальных этапах производства сыра должно быть компенсировано, так как в сырах вся лактоза должна быть сброжена и ослабление активности микрофлоры закваски на начальном этапе обусловит необходимость более интенсивного ее размножения на последующих. Тем не менее факты свидетельствуют, что этот прием снижает опасность появления горечи в Чеддере. Это можно объяснить тем, что в Чеддере на последующих этапах резко понижается активность воды в результате посолки, что ингибирует протеолиз, особенно расщепление β-казеина и, как следствие, образование горьких пептидов. В Чеддере горечь не появляется при концентрации соли в водной фазе более 4,7%. Если это единственная причина исчезновения горького вкуса в сырах при снижении плотности популяции микроорганизмов закваски во время выработки, то в сырах с посолкой в рассоле снижение протеиназной активности микрофлоры заквасок на начальном этапе окажет меньшее влияние на уменьшение горечи, так как соль в них медленно распространяется внутри головки и поэтому длительное время не может оказать непосредственного влияния на протеолиз в большей части массы головки. Правда, и в этом случае соль может оказать влияние на протеолиз за счет создания градиента активности воды между поверхностными и внутренними слоями сыра и усиления миграции влаги от центра к поверхности. Кроме этого, гидролиз β-казеина микрофлорой закваски начинается спустя месяц после выработки сыра, когда активность воды снизится и в глубинных слоях. Возможно также, что главная причина появления горечи только при высокой концентрации клеток лактококков в конце выработки сыра заключается в том, что про- теиназы, ассоциированные с клеточной стенкой, наиболее активно образуют горькие пептиды при повышенных уровнях pH и температуры, характерных для сыра во время выработки, которые начинают снижаться уже во время посолки. В этом случае снижение протеиназной активности микрофлоры закваски на начальном этапе будет эффективным средством уменьшения горечи в сырах с посолкой в рассоле.
Соль незначительно влияет на первичную стадию гидролиза asi-казеина - отщепление от него молокосвертывающими энзимами as1-I пептида; последующий гидролиз asrI пептида в большой степени зависит от содержания соли в водной фазе сыра.
Протеиназная активность лактококков в конце стационарной фазы, по сравнению с концом логарифмической, ниже для «горького» штамма Lc. lactis на 10%, а для «негорького» - на 80%. После окончания сбраживания лактозы, т. е. во время созревания сыра, лактококки находятся в фазе гибели, и их протеиназная активность должна все время уменьшаться.
Снижение дозы закваски, повышение температуры II нагревания, т. е. меры, лимитирующие накопление биомассы лактококков во время выработки, уменьшают вероятность появления горечи в сыре Гауда. Пользоваться этими рекомендациями нужно осторожно, так как низкая активность молочнокислого брожения во время выработки создает условия для быстрого развития посторонней микрофлоры.
Внутриклеточные пептидазы «негорьких» штаммов разрушают горькие пептиды при pH больше 4,5, «горьких» штаммов - при pH только выше 5,5. Minagawa et al. показали возможность предотвращения горького вкуса в сырах с помощью аминопептидазы Т, расщепляющей горькие пептиды. Добавление в молоко для выработки Российского сыра со свиным пепсином культуральной среды «негорького» штамма Lc. lactis Е8 в 2,7 раза снизило содержание горьких пептидов в продукте (табл. 11.8). Багдонайте считает, что штаммы лактококков с повышенной аминопептидазной активностью предотвращают появление горечи в сырах.
Горечь в Чеддере, появившаяся из-за использования повышенных доз реннета, исчезала при внесении в молоко определенных доз фагагомолога к штамму лактококков, с которым вырабатывали сыр. Это можно объяснить тем, что горькие пептиды, образуемые реннетом, расщепляются внутриклеточными энзимами лактококков, которые высвобождаются во время лизиса клеток бактериофагами.
Применение закваски, содержащей несбраживающие лактозу и не обладающие протеиназной активностью мутанты лактококков, позволило выработать сыр с повышенным содержанием САК, не имеющий горечи. Пониженная способность закваски сбраживать лактозу позволяет увеличить ее дозу без повышения кислотности сырной массы выше оптимального уровня, а высокие дозы закваски при отсутствии протеиназной активности, связанной с клеточными стенками, увеличивают способность расщеплять пептиды до САК внутриклеточными пептидами (освобождающимися в процессе автолиза) без образования горьких пептидов.
Молокосвертывающие энзимы и микрофлора закваски - не единственные причины образования горьких пептидов в сырах. Энзимы, способные их образовывать, также выделяют психротрофы в молоке до пастеризации (разд. 11.2.2). Интенсивность вызываемой ими горечи возрастает при увеличении продолжительности созревания и хранения сыра.
Размножение в сырах Ent. faecalis subsp. liquefaciens, обладающего мощной протеолитической активностью, может вызвать появление сильной горечи. Порок возникает при исходном содержании в молоке свыше 106 КОЕ/мл этого подвида, что при использовании пастеризации молока вряд ли возможно. Однако при низкой активности микрофлоры закваски энтерококки могут размножиться в сыре до критического уровня и при меньшей исходной их дозе в молоке. Появление горечи в этом случае сопровождается возникновением мажущейся консистенции.
Горечь в сырах, выработанных в асептических условиях, была выше, чем в контрольных, выработанных из этого же молока и с этой же закваской в открытых ваннах. Это свидетельствует о наличии среди незаквасочной микрофлоры бактерий, разрушающих горькие пептиды, которыми скорее всего являются лактобациллы. Allen & Knowles еще в 1934 г. указывали, что без молочнокислых палочек сыры не имеют должного аромата и горькие на вкус. Способность лактобацилл уменьшать горечь в сырах обусловлена их высокой аминопептидазной активностью.
Снижение температуры созревания Чеддера с 13 до 6° С усиливает горечь. Это связано с ингибированием развития лактобацилл и снижением активности внутриклеточных протеиназ лактококков, разрушающих горькие пептиды, образуемые молокосвертывающими энзимами из а-казеинов, низкими температурами. Вероятность появления горечи увеличивается в сырах с низким pH, что связано со снижением активности энзимов, расщепляющих горькие пептиды.
САК и вкус сыра. Роли САК в формировании вкуса сыра посвящено много исследований. Аминокислоты могут быть сладкими, горькими, напоминать по вкусу мясной бульон. В сочетании с другими соединениями САК могут приобретать самый разнообразный вкус. Сторонником гипотезы о тесной корреляции аминокислотного состава с вкусом сыра был Диланян. Он считал, что в состав заквасок нужно включать только те штаммы лактобактерий, которые образуют в молоке свободные САК в том соотношении, в котором они находятся в сырах высшего сорта вырабатываемого вида. В качестве эталона были установлены следующие аминограммы (в % от общего содержания аминокислот): для сыра Швейцарского - лизин 13-14, треонин 5-6, глутаминовая кислота 17-18, пролин 12-13, лейцин 9-10; для сыра Советского- лизин 10, глутаминовая кислота 14-18, пролин 8-10, лейцин 15, фенилаланин 6, серин 10 и валин 10. Диланян с учениками упорно разрабатывали эту тему, но выдвинутая им концепция не получила признания в мировых научных кругах. Причиной этого, очевидно, являются неодинаковые условия для протеолиза лактобактериями в молоке и в сыре, в результате чего спектры образуемых ими САК в средах и сыре не совпадают. Кроме этого, микрофлора закваски образует значительное количество свободных аминокислот не из казеинов, а из продуктов их расщепления молокосвертывающими энзимами.
Чеботарев выявил отличия качественного состава свободных аминокислот в сыре и исходном казеине, что обусловлено трансформацией аминокислот в процессе созревания сыров. В зрелых сырах нет серина, аргинина и метионина, в некоторых сырах отсутствует гистидин, несмотря на их присутствие в казеине. В то же время в сырах появляются орнитин, а- и у-аминомасляная кислоты, отсутствующие в исходном казеине. По данным Чеботарева, в сырах могут трансформироваться аланин в аммиак и пировиноградную кислоту, глутаминовая кислота - в аммиак и а-кетоглутаровую кислоту, лейцин - в масляную кислоту, треонин и валин - в пропионовую кислоту, аспарагиновая кислота - в янтарную, глицин - в глиоксалевую, метионин - в а-амино- масляную кислоты. В молоке такая трансформация аминокислот может и не иметь места.
Долгое время существовало мнение, что САК, кроме пролина в сырах с высокой температурой II нагревания, непосредственно не играют существенной роли в формировании вкуса сыров, выполняя функцию предшественников вкусовых и ароматических веществ. Главное внимание при исследовании вкусообразования в сырах уделяли летучим соединениям, образующимся в результате липолиза и трансформации САК.
Несмотря на то что выдвинутый Диланяном принцип подбора микрофлоры заквасок по спектру образуемых в молоке аминокислот оказался несостоятельным, его и Чеботарева мнение о важнейшей роли свободных аминокислот, а также других нелетучих водорастворимых продуктов протеолиза, в формировании вкуса и аромата сыров получает подтверждение в последних работах. McGugan et al. показали, что растворимая, нелетучая фракция азота наиболее важная детерминанта сырного вкуса (не обязательно качества). Безвкусные, освобожденные от жира остатки старых сыров оказывают более сильное влияние на интенсивность сырного вкуса реконструируемых фракций сыра, чем равные количества подобных остатков молодых сыров. Напрашивается вывод, что вкусовые вещества становятся более доступными компонентами вкуса после разрушения структуры сыра.
Ельцова еще в 1961 г. показала, что в Голландском сыре хорошего качества было больше свободных аминокислот, особенно глутаминовой, чем в сырах с недостаточно выраженным вкусом и ароматом. Stadhouders et al. установили, что сыры, выработанные с протеиназо-отрицательными мутантами лактококков (Прт), имели по сравнению с контрольными сырами одинаковое содержание растворимого, но значительно более низкое содержание аминного азота, что коррелировало с их менее выраженным сырным вкусом. Farkye & Fox также получили подобный результат.
Установлена корреляция между содержанием в сыре Фен, Ала, Про, Глу и Асп и продолжительностью созревания, но не качеством сыра.
В асептически выработанных сырах выявлена корреляция между количеством САК и выраженностью сырного вкуса. По мнению ряда авторов, молокосвертывающие энзимы и эндопептидазы лактококков оказывают основное влияние на формирование вкуса и аромата сыров с низкими температурами II нагревания, а основными продуктами совместного действия этих факторов являются САК. Содержание растворимого в фосфорно-вольфрамовой кислоте азота (в основном азот свободных аминокислот) предлагается использовать как показатель степени выраженности вкуса и аромата сыра.
Для улучшения вкуса, аромата и ускорения созревания египетского сыра Рас в сгусток вносили равные количества аспарагиновой и глутаминовой кислот (по 1 или 2 мг/кг) с добавлением или без добавления 5 г/100 кг молока Na2HP04 . Добавление аминокислот улучшало вкус, но приводило к получению сыра более твердой консистенции. Добавление Na2HP04 устраняло этот порок консистенции. Добавленные аминокислоты ускоряли протеолиз и созревание сыра пропорционально дозе, фосфат также ускорял созревание. Этот опыт подтверждает важную роль САК в формировании вкуса сыра.
Есть экспериментальные данные, которые, на первый взгляд, опровергают эту точку зрения. Раманаускас и Шаломскене показали, что в нестандартном по вкусу и запаху Российском сыре содержится больше САК и летучих жирных кислот (ЛЖК) . В справедливости установленного ими факта не приходится сомневаться, поскольку он получен в результате исследования большого количества сыров промышленной выработки. Однако однозначно интерпретировать его невозможно, так как такое повышение содержания свободных аминокислот и, особенно, жирных кислот вряд ли можно объяснить результатом жизнедеятельности микрофлоры закваски. Скорее всего, это результат жизнедеятельности посторонней микрофлоры, например, психротрофов, которые образуют не только САК, но и десятки других соединений, способных ухудшить органолептические показатели сыров. К таким соединениям можно отнести свободные карбоновые кислоты, содержание которых в дефектных сырах было в 4,6 раза выше, чем в доброкачественных.
То, что повышенное содержание САК не могло быть непосредственной причиной забраковки Российского сыра, доказывает другая работа Раманаускаса и Меялене. В этой работе для изучения возможности повышения степени использования белков молока в сыроделии в смесь для выработки Российского сыра вносили сухие концентраты сывороточных белков с разной массовой долей нативных сывороточных белков. Внесение их в молоко вызвало более интенсивное образование САК в сыре: в 10-дневном сыре количество САК увеличилось в 3 раза, в конце созревания - примерно в 9 раз. Преобладающими кислотами были глутамин, лейцин и лизин, массовая доля которых составила 44% от общего количества САК. Параллельно с увеличением содержания САК в сырах в процессе созревания увеличивалась концентрация свободных жирных кислот (СЖК): в сырах зрелого возраста их было в 2,2 раза больше, чем в сырах после прессования. В кондиционном возрасте опытные сыры обладали нежной, пластичной, однородной по всей массе консистенцией и умеренно выраженным, слегка кисловатым вкусом, получившим более высокую оценку, чем вкус контрольного сыра. Авторы делают вывод, что повышение содержания нативных сывороточных белков в сыре интенсифицирует накопление вкусовых и ароматических веществ, не меняя направленности биохимических процессов созревания. Сывороточные белки не атакуются или очень плохо атакуются молокосвертывающими энзимами и микрофлорой мезофильных заквасок. Их добавление в сыры уменьшает потери влаги сыром во время созревания и хранения, что и должно было интенсифицировать процессы созревания. При этом сыры вырабатывались в экспериментальных условиях, при которых их загрязнение посторонней микрофлорой было сведено до минимума. В опытных сырах содержание молочнокислых бактерий в период максимума было выше, чем в контрольных. В данном случае увеличение содержания САК в опытных сырах произошло за счет жизнедеятельности микрофлоры закваски, и оно положительно повлияло на формирование и характер органолептических показателей сыра.
Law et al. увеличили в 3 раза содержание САК в Чеддере, выработанном в асептических условиях, внесением в молоко обработанных лизоцимом клеток лактококков, потерявших способность сбраживать лактозу. Выраженность сырного вкуса в опытных сырах по сравнению со вкусом сыров, выработанных с обычной закваской, не изменилась, но в них реже обнаруживалась горечь. Отсюда они делают вывод об отсутствии прямого влияния САК на вкус сыра. Снижение горечи в их опыте можно объяснить более ранним высвобождением внутриклеточных энзимов из обработанных лизоцимом клеток.
Выработки в асептических условиях доказали ведущую роль лактококков в образовании САК в сырах с низкими температурами II нагревания и большие различия между заквасками по этому показателю; работы по поиску метода оценки протеолитической активности заквасок должны быть продолжены. Испытание в производстве сыра Гауда двух «быстрых» штаммов Lc. cremoris с высокой активностью образования небелкового азота показали, что один из них, образующий большое количество глутаминовой кислоты, лейцина и треонина, дал хорошо выраженный сырный вкус, второй образовывал горечь.
Лахт и Вилу считают, что каждый вид сыра характеризуется определенным составом САК. Сыры Эдам, вырабатываемые в Финляндии и Венгрии, содержат валин в концентрации 5,6 и 5,9 ммоль/г, метионин - 1,7 и 1,6 ммоль/г, аргинин - 1,2 и 1,2 ммоль/г соответственно. Эмментальские сыры немецкого и французского производства содержали валина 11 и 10 ммоль/г, метионина - 2,4 и 2,4 ммоль/г соответственно. Суммарное содержание САК в обоих сырах составляло 121 ммоль/г. Соотношение Глу/Ала в Эмментальском сыре высокого качества, выработанном в Эстонии, в трехмесячном возрасте равняется 2,66; в выработанном в Финляндии - 2,67 в 9-месячном возрасте. В эстонском Эмментале выше содержание у-казеина, что авторы объясняют более высоким содержанием в молоке соматических клеток, содержащих энзимы, гидролизующие β-казеин с образованием γ-казеина.
Engels & Visser исследовали вклад водорастворимых соединений сыров Чеддер, Гауда, Грюйер, Маасдам, Пармезан в формирование вкуса и аромата. Они нашли, что наибольшее влияние на вкус и аромат сыров оказывают низкомолекулярные соединения (меньше 500 Да), включая пептиды,аминокислоты, свободные жирные кислоты и продукты их расщепления.
Общая протеолитическая активность и качество сыра. Термофильные лактобациллы играют главную роль в интенсивности и специфичности протеолиза, формировании вкуса и аромата в крупных сырах. Включение термофильных и, в меньшей степени, мезофильных лактобацилл в закваски для мелких сыров придает нетипичный для них пряный, сладковатый привкус, характерный для крупных сыров, обусловленный высоким содержанием свободного пролина.
В изучении влияния специфичности протеолитической активности микроорганизмов на органолептические показатели сыра есть определенная трудность, поскольку различия в протеолитической активности сопровождаются различиями в гликолитической активности, играющей важнейшую роль на первых этапах производства сыра. В настоящее время при проведении подобных опытов микроорганизмы подвергают шоковым обработкам, после которых клетки теряют способность сбраживать углеводы во время выработки сыра, но сохраняют протеолитическую активность, в частности активность внутриклеточных протеолитических энзимов.
Лактобациллы незаквасочного происхождения могут интенсифицировать протеолиз и ускорить формирование сырного вкуса, что используют в производстве низкожирных сыров, главным недостатком которых при выработке на лактококковых заквасках являются слабовы- раженный вкус и аромат, твердая консистенция. Внесение в смесь для выработки сыра 10 %-ной жирности термофильных лактобацилл, находящихся в состоянии теплового шока, ликвидировало в нем горечь и усилило аромат: органолептические показатели опытных сыров были такие, как у сыров с 28% жира.
Добавление в молоко для производства сыра Гауда термофильных лактобацилл, подвергнутых шоку, усилило выраженность сырного вкуса и аромата и ликвидировало горечь. Положительный эффект был выше с Lbc. helve tic us, чем с болгарской палочкой. В другом опыте для усиления выраженности вкуса Чеддера с уменьшенным на одну треть содержанием жира в молоко вносили культуры Lbc. helveticus с ослабленной гликолитической активностью (распылительной сушкой при 82 или 120° С, замораживанием); в опытных сырах увеличивалось содержание аминного азота и улучшался вкус. Благодаря ослабленной гликолитической активности лактобациллы не изменили скорость кислотообразования во время выработки сыра.
Добавление в молоко для выработки Чеддера, наряду с обычной закваской, 106 КОЕ/мл лактобацилл ускорило протеолиз и формирование органолептических показателей . Сатиева и Гаврилова ускорили созревание Костромского и Пошехонского сыров использованием в качестве закваски 0,75% лактококков и 0,25% термофильных лактобацилл.
Греческие ученые увеличили на 35% количество растворимого в ТХУ азота в рассольном сыре Фета добавлением в молоко нейтральной или кислой протеиназ микробиального происхождения (В. subtilis и Aspergillus oryzae) и шокированных нагреванием культур термофильного стрептококка и болгарской палочки. Опытные сыры в 40-суточном возрасте имели вкус и консистенцию, подобные вкусу и консистенции 80-суточных контрольных сыров, однако для сбалансирования вкуса и аромата в опытные сыры необходимо также добавлять липазу. Увеличение количества бактериальных протеиназ приводило к появлению горечи.
Сделана попытка улучшить потребительские свойства маложирного сыра Чеддер введением в закваски педиококков. Опытные сыры отличались от контрольных ускорением протеолиза и более высоким содержанием сероводорода, метанэтанов, играющих важную роль в формировании аромата сыров, ацетатов. Маложирные сыры, выработанные с использованием в составе закваски педиококков, имели выраженные вкус и аромат полножирного сыра, отсутствующий у контрольных сыров. Однако в опытном сыре в 2-месячном возрасте появились кристаллы лактата, которых не было в контрольных сырах на протяжении 4-5 мес созревания, что обусловлено трансформацией педиококка- ми L(+)-лактатов в D(-)-лактаты. Использование для этой цели микрококков привело к появлению привкусов в сыре.
Беловым с соавт. установлено сильное влияние особенностей протеолиза заквасок одного и того же видового состава на органолептические показатели Советского сыра. Наивысшую оценку в их опытах получили сыры с минимальным содержанием фракции небелкового азота и средним содержанием аминного азота (отношение небелкового азота к аминному - 1,89), в сырах с минимальной оценкой это отношение равнялось 2,0-3,0; в сырах с низкой оценкой более интенсивно расщеплялся уЗ-казеин. Разница между оценками сыров высокого и низкого качества в этих опытах составила 4 балла. К сожалению, в опыте не учтено влияние незаквасочной микрофлоры на протеолиз.
В крупных сырах с поздним вспучиванием наблюдается более интенсивное расщепление белков, что, по мнению Steffen, стимулируется излишним развитием пропионовокислых бактерий. В дефектных сырах, например, активность лейцинаминопептидазы была в 2,3 раза выше, а количество лактатов в 1,6 раза ниже, чем в сырах без порока. В этом случае причиной позднего вспучивания следует считать те факторы, которые способствовали интенсификации протеолиза в дефектных сырах, поскольку протеолитическая активность самих пропионовокислых бактерий слишком низка, чтобы заметно изменить степень протеолиза в сыре. Высокое содержание небелкового азота приводит к формированию крошливой, малосвязной консистенции; недостаточный протеолиз - к слабовыраженному вкусу, нетипичной консистенции, неравномерному рисунку.
Можно считать, что для органолептических показателей сыра вреден недостаточный и избыточный протеолиз, особенно вызываемый посторонней микрофлорой, продукты которого отличаются от продуктов протеолиза микрофлорой закваски количественно и качественно.
Влияние продуктов трансформации аминокислот на органолептические показатели сыра. Во время созревания аминокислоты подвергаются трансформации путем декарбоксилирования, дезаминирования, десульфурилирования и деметиолирования (рис. 11.6). Продукты трансформации аминокислот играют важную роль в формировании органолептических свойств сыров.
В результате декарбоксилирования аминокислот образуются амины. Их содержание в сыре может повлиять на показатели безопасности продукта, что рассмотрено в гл. 13, а также представляет интерес с точки зрения формирования вкуса и аромата. Декарбоксилирование происходит под действием декарбоксилаз, образуемых микроорганизмами; микрофлора заквасок и гомоферментативные лактобациллы амины в сырах не образуют (разд. 13.9). Правда, Морина полагает, что некоторые штаммы молочнокислых бактерий образуют незначительные количества летучих аминов в питательных средах, но это не значит, что они будут образовывать амины в сыре. Тем не менее амины в сырах есть, особенно в сырах с длительным созреванием или хранением. В табл. 11.12 показано, например, содержание аминов в крупных сырах, вырабатываемых в Швейцарии. В твердых и мягких сырах, вырабатываемых в Польше, содержание тирамина варьировало в пределах 3,8-575 мкг/г, гистамина - 0—157 мкг/г. Содержание тирамина и гистамина в некоторых сырах показано в табл. 13.7.

Амины попадают в сыры с молоком или образуются посторонней микрофлорой, например, энтерококками, гетероферментативными лактобациллами (Lbc. buchneri). Содержание аминов в Грюйере и Аппенцеллере выше, чем в Эмментале, что обусловлено участием поверхностной слизи в созревании первых двух видов сыров. Br. linens обладает способностью декарбоксилировать аминокислоты и образовывать амины. В культуральной среде этого вида нашли шесть летучих аминов (кадаверин, моноэтиламин, монометиламин, диметиламин, триэтиламин и аммоний) и два нелетучих: тирамин и гистидин. Tsugo & Hasono считают, что кадаверин и путресцин, образующиеся при декарбоксилиро- вании, соответственно, лизина и орнитина, играют роль в формировании вкуса сыров, созревающих при участии белой плесени. Содержание путресцина и кадаверина в этих сырах в 3-недельном возрасте составило 724 и 537 мг/кг. В твердых сырах из пастеризованного молока кадаверин и путресцин (обладающие ядовитыми свойствами и неприятным запахом) большинство авторов или вообще не обнаруживали, или обнаруживали в мизерных количествах.
Учитывая низкие концентрации аминов в сырах, можно считать, что их роль в формировании вкуса и запаха твердых сыров весьма сомнительна. Этому, на первый взгляд, противоречат результаты исследований, выполненных под руководством Головни. В этих опытах до 35 летучих аминов обнаружено в Голландском и Российском сырах, в рассольном сыре Чанах. Между количеством и спектром летучих аминов в Российском и Голландском сырах наблюдалось большое сходство, хотя по вкусу эти сыры имеют заметные различия. Не было существенных различий между общим содержанием летучих аминов в Российском и Голландском сырах рассольном сыре, которое равнялось соответственно 36,2; 39,8 и 39 мг/кг. Различия обнаружены в количестве отдельных аминов. В сыре Чанах среди летучих аминов в количественном отношении доминировал ди-н-бутиламин, относительное содержание которого в Российском и Голландском сырах не превышало 2%; в Российском и Голландском сырах максимальная концентрация была у диизопропиламина (около 9%), а в сыре Чанах этот амин присутствовал в следовых количествах. Увеличение продолжительности хранения Российского сыра при минус 3° С до 10 мес существенно изменило содержание в нем некоторых аминов (табл. 11.13). За 6 мес хранения балловая оценка сыра снизилась с 93 до 89 баллов, а количество летучих аминов увеличилось в 1,28 раза, в основном за счет пиперидина и а-пиколина. Авторы этих работ считают возможным увязать снижение качества сыра с увеличением в сыре содержания летучих аминов. Вряд ли такое предположение корректно, так как оценка сыра снизилась в основном за консистенцию, оценка за вкус и запах понизилась только на один балл. Причастность летучих аминов к формированию консистенции, учитывая их мизерные концентрации, сомнительна.

Независимо от того, принимают или не принимают участие в формировании органолептических свойств сыров амины, производство сыра нужно вести так, чтобы их содержание было минимальным и не могло вызвать опасность для здоровья потребителя. Количество аминов в сыре Гауда увеличивается при высоких температурах созревания (18-21° С), продолжительном созревании, с развитием в сырах гетероферментативных лактобацилл, в частности Lbc. buchneri. Солеустойчивые лактобациллы, попадающие в сыр из рассола низкого бактериального качества, образуют в сырах путресцин и кадаверин; энтерококки могут образовать амины, если их содержание в сыре превысит 10' КОЕ/г.
Manning & Robinson из летучей фракции зрелого Чеддера, имеющей запах сыра, выделили три серосодержащих соединения, образованные из метионина: метантиол, диметилсульфид и сероводород. Метантиол (CH3SH) был обнаружен в двух сырах, выработанных в асептических условиях с применением закваски и обладающих нормальным вкусом и ароматом, но не был найден в параллельных сырах, выработанных с применением лактона глюконовой кислоты вместо закваски и не обладающих необходимым вкусом и ароматом, а также в сырах из обезжиренного молока. H2S обнаружен во всех четырех сырах, но его содержание в сырах с нормальным вкусом и ароматом было в два раза выше, чем в безвкусных сырах. Диметилсульфид обнаружен во всех сырах в равных концентрациях. Авторы сделали вывод о необходимости метантиола для формирования вкуса и аромата сыра Чеддер и об участии в этом процессе H2S. Образуется метантиол микрофлорой закваски (в сырах, выработанных в асептических условиях без закваски, он не был обнаружен). По-видимому, в его образовании принимает участие H2S. Коэффициент корреляции между вкусом зрелого Чеддера и содержанием в нем H2S составляет 0,66-0,79. Однако мнения о роли метантиола во вкусообразовании в твердых сычужных сырах, например, в Чеддере расходятся. Некоторые считают ее весьма существенной, Aston & Douglas ее отрицают.
В опытах Barlow et al. на вкус сыра Чеддер наибольшее влияние оказывали серосодержащие соединения, особенно H2S, молочная кислота, бутанон, общее содержание водорастворимого азота и уксусной кислоты. Авторы считают, что по содержанию водорастворимого азота и H2S в 3-месячных сырах можно предсказать вкус 12-месячных сыров. Decrement & Vickers считают наиболее важными для формирования вкуса и аромата Чеддера масляную кислоту, диацетил и метантиол.
По-видимому, наиболее верной является третья точка зрения: в твердых сырах метантиол и сероводород играют положительную роль только в определенных концентрациях, превышение которых ведет к появлению пороков вкуса. Скорее всего, метантиол оказывает воздействие на вкус и аромат Чеддера, вступая в реакции с другими веществами с образованием таких соединений, как тиоэфиры, например, метилтиоацетат.
Метантиол играет важную роль в формировании вкуса и аромата грибных сыров и сыров, созревающих с участием поверхностной слизи и плесневых грибов. В частности, он принимает участие как компонент и предшественник в формировании характерного для многих грибных сыров «чесночного» вкуса и запаха. Интересно, что метантиол и H2S в чистом виде обладают крайне неприятными запахами. Сернистые соединения обладают низким вкусовым порогом; они растворимы в жире, но не в воде, что частично объясняет отсутствие сырного вкуса в сырах из обезжиренного молока. Некоторые сернистые соединения присутствуют в молоке, из которого почти полностью переходят в сыр, что увеличивает их концентрацию в сыре по сравнению с исходным молоком примерно в 10 раз.
Singh & Kristoffersen считают, что диметилсульфид играет важную роль в формировании вкуса и аромата крупных сыров, но только при определенной концентрации. Избыток его ухудшает вкус сыров. В крупных сырах диметилсульфид образуют пропионовокислые бактерии.
Часть жирных кислот с короткой цепочкой и карбонильных соединений также образуется в результате катаболизма аминокислот (рис. 11.6). В сыре Гауда обнаружены молочная, пропионовая и пировиноградная кислоты, содержание которых увеличивалось, и оротовая и уксусная кислоты, содержание которых уменьшилось во время созревания.
Установлено образование из карбонильных соединений (глиоксаль, метилглиоксаль, дигидрооксиацетон, ацетальдегид) и САК (валин, лейцин, изолейцин, метионин, цистеин, фенилаланин, пролин и лизин) ароматических соединений сыров, в том числе 2-ацетил-тиазола из цистеина и метилглиоксаля и алкилпиразинов из лизина и дигидрооксиацетона. В образовании этих соединений принимают участие лактобациллы.
Сырные вкус и аромат являются результатом тесного взаимодействия продуктов протеолиза и липолиза. Silverman & Kosikowski отмечают, что смеси жирных кислот в водных растворах имеют неприятный и прогорклый вкус, который передается сырной массе при их внесении. Смеси аминокислот придают свежей сырной массе приятный вкус, слегка напоминающий вкус Швейцарского сыра. Добавление же аминокислот и жирных кислот дает приятный острый вкус, напоминающий вкус зрелого Чеддера.
