Обезвоживание сырной массы достигается совместным воздействием на нее второго нагревания и обработки сырного зерна. Первый вариант данной серии опытов (ванна первая) отражает часто встречающиеся в практике случаи, когда повышенная температура второго нагревания при длительной обработке сырного зерна приводит к значительному обезвоживанию сыра; третий вариант (ванна третья) обеспечивает получение сыра с нежной консистенцией (высокой влажностью) в условиях низкой температуры второго нагревания.
В этой серии опытов подготовка молока проводилась в том же порядке, что и в предыдущих. Продолжительность обработки сырного зерна в первой ванне составляла 118 мин. при температуре второго нагревания 46°, во второй — 75 мин. при 43° и в третьей — 65 мин. при 40°. Посолка сыра из первой ванны длилась 7 суток, из второй 6 и из третьей — 5,5 суток.
Химические показатели сыра после прессования и зрелого приведены в табл. 9.
При температуре второго нагревания 46° и обработке сырного зерна в течение 118 мин. происходило значительное обезвоживание (влажность сыра после прессования 38,8%).
С понижением температуры второго нагревания и сокращением длительности обработки влажность сыров после прессования повышалась (из второй ванны до 41,9%, из третьей—до 44,7%).
С повышением влажности увеличивалось в сыре количество молочного сахара, однако его концентрация в водной фазе находилась почти на одном уровне во всех сырах, несмотря на более длительную обработку сырного зерна в первой ванне. Это объясняется тем, что высокая температура действовала подавляюще на развитие бактерий, вследствие чего замедлилось сбраживание молочного сахара.
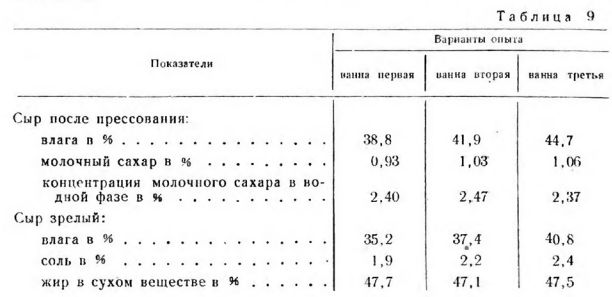
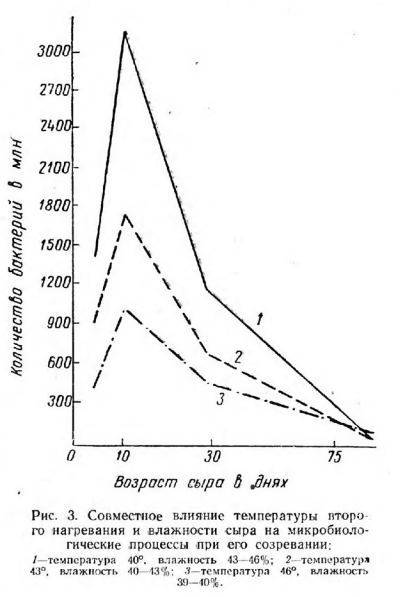
Наибольшего объема микробиологические процессы достигли в сырах, выработанных при пониженной температуре второго нагревания (40°), и большей влажностью (рис. 3 и табл. 10).
Повышение темпе ратуры до 43° подавляло развитие бактерий примерно в 2 раза.
Однако уровень микробиологических процессов в этих сырах был все еще достаточно высоким (913—1774—705 млн.) Резкое торможение развития микрофлоры наблюдалось при повышении температуры второго нагревания до 46° и понижении влажности сыра до 38,8%.
Сопоставление данных, показанных на рис. 1 (влияние температур второго нагревания на развитие бактерий при одинаковой влажности сыра после прессования) и на рис. 3 (влияние температур второго нагревания и влажности), .позволяет отметить большое сходство в развитии микробиологических процессов, хотя сыры различались по влажности и продолжительности обработки сырного зерна.
Так, в первой серии опытов (рис. 1) продолжительность обработки зерна при 46° в 2 раза меньше, чем в данной серии (54 и 118 мин.), и влажность 41,1 и 38,8%, однако подавление развития бактерий было почти одинаковым (в сыре после прессования 355—453 млн., и через 10 дней— 1263—1198 млн.).
При выработке сыра продолжительность обработки всегда сочетается с температурой второго нагревания и влажностью сыра, которые оказывают существенное влияние на микробиологические процессы. Объем микрофлоры в сыре мало зависит от температуры созревания; наиболее интенсивно микробиологические процессы протекают в первые 10 дней его созревания (при выработке, прессовании и посолке), а после израсходования молочного сахара они идут уже на убыль.
Таким образом, главным регулятором объема микробиологических процессов в производстве сыра является температура второго нагревания, а вторым по значимости регулятором — влажность сыра после прессования.
Сопоставление данных (табл. 10), полученных при непосредственном подсчете, с содержанием бактерий при посеве сыра на твердую среду позволило отметить быстроту автолиза бактериальных клеток.
В сырах всех трех вариантов автолиз происходил активно в течение 30 дней и постепенно замедлялся с возрастом сыра, что объясняется, по- видимому, накапливанием продуктов распада и «консервированием» клеток.
С целью выявления селекционирующего дейстзия указанных факторов было проведено детальное изучение микрофлоры закваски и сыра. При исследовании микрофлоры закваски выявлено, что около 60% всех бактерий составляли Str. lactis, 39,5% — Str, diacetilactis и 0,5% — Str. paracitrovorus; молочнокислые палочки при посевах не были обнаружены. Преобладающее большинство (87,5%) штаммов в закваске оказалось энергичными кислотообразователями (при температуре 30° через 48 час., кислотность составляла 90—118° Т); а остальные были более слабыми (кислотность 80—89°Т и ниже).
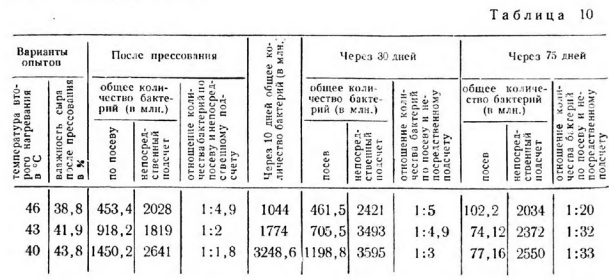
Результаты исследований, характеризующие состав микрофлоры и биохимическую активность штаммов, выделенных из сыра, представлены на рис. 4.
Температура второго нагревания является главным фактором, который не только регулирует объем микробиологических процессов, но и селекционирует микрофлору сыра. Изменение температуры нагревания на 6° и влажности сыра на 5% оказывает сильное воздействие в первую очередь на развитие ароматообразующих стрептококков тина Sir. diacetilactis и молочнокислых палочек.
Первой отличительной особенностью сыров, выработанных при температуре второго нагревания 46° (с влажностью 38,8%), являлось активное развитие молочнокислых палочек с самого начала процесса, в результате чего количество их после прессования составляло 16,6% от всей микрофлоры сыра. В дальнейшем, в процессе созревания сыра с температурой второго нагревания 46°, содержание молочнокислых палочек еще более , увеличивалось и достигло через 30 дней 52,5%, а через 75 дней — 72% всей микрофлоры.
Второй особенностью являлось подавление развития ароматообразующих стрептококков типа Str. diacetilactis. Они составляли только 12,5% от всей микрофлоры, тогда как в закваске их было около 40%.
Количество ароматообразуюшлх Str. diacetilactis и Str. paracitrovorus на всем протяжении созревания сыра находилось на низком уровне.
Понижение температуры второго нагревания только на 3°, а тем более на 6° и соответственно повышение влажности сыра до 41,9 и 43,8% вызвало резкие изменения в составе микрофлоры сыра.
В сырах, выработанных при более низкой температуре второго нагревания (43 и 40°) весьма активно развивались ароматообразующие бактерии Str. diacetilactis. Количество их в сыре после прессования и через 30 дней составляло 49—43% всей микрофлоры и было более высоким,чем в производственной закваске (40%), но к концу созревания, через 75 дней, наблюдалось значительное их вымирание, причем более сильное в сырах с температурой второго нагревания 43°. Количество Str. paracitrovorus и в этих сырах было ничтожным (0,7—2,7%).
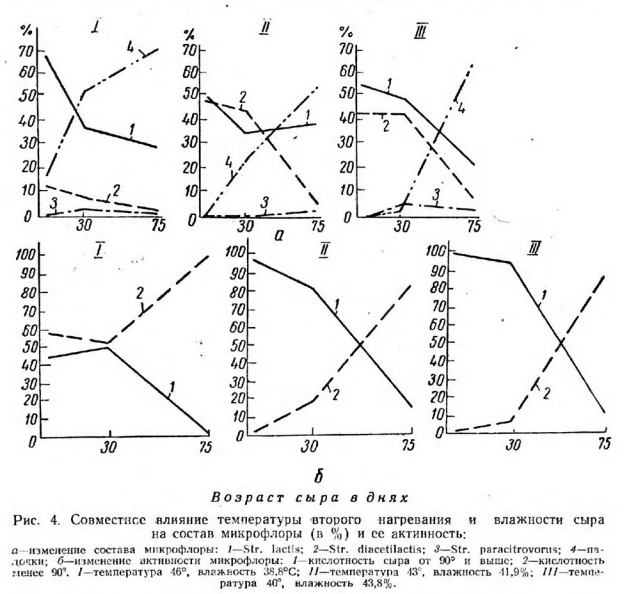
В этих сырах в первые 30 дней происходило менее интенсивное развитие молочнокислых палочек. В сырах, выработанных при температуре второго нагревания 43° (с влажностью 41,9%), развитие палочек протекало несколько активнее: количество их после прессования составляло 0,7%, а через 30 дней—20% всей микрофлоры. Тогда как в сырах при температуре второго нагревания 40° (с влажностью 43,8%) палочек после прессования не было, а через 30 дней количество их все еще было незначительным — 3,5%.
Правда, к концу созревания сыров количество молочнокислых палочек почти достигло уровня, отмеченного в сырах с температурой второго нагревания 46° (рис. 5).
В сырах с температурой второго нагревания 46° (влажность 38,8%) развитие молочнокислых палочек, энергичных по кислотообразованию (128—180°), было наиболее интенсивным. В сыре после прессования они составляли 16,6%, а через 30 дней содержание их увеличивалось до 38% от всей микрофлоры, но в 75-диевных сырах этих палочек уже не было.
В сырах с температурой второго нагревания 43э количество молочнокислых палочек—энергичных кислотообразователей — было значительно меньшим: после прессования 0,7%, а через 30 дней 20%. Наименьшее количество их было в сырах с температурой второго нагревания 40 : в сырах после прессования их не было, а через 75 дней они составляли всего 3% от всей микрофлоры.
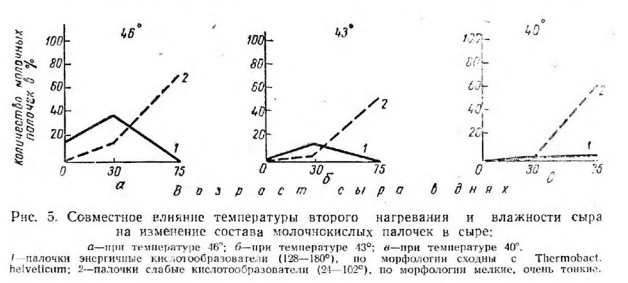
Молочнокислые палочки, являющиеся слабыми кислотообразователями (28—50°), мы обнаружили в сыре только через 30 дней.
Однако и в этом случае наибольшее количество их (14,5%) наблюдалось в сырах, выработанных при температуре второго нагревания 46°, пониженное (5%) — при температуре второго нагревания 43° и самое низкое (2,5%) —при 40°. В дальнейшем, к концу созревания сыров всех трех вариантов, наблюдается сходство в развитии молочнокислых палочек со слабой способностью к кислотообразованию: количество их через 75 дней достигло соответственно температуре второго нагревания 73, 59 и 67%. Следовательно, увеличение количества молочнокислых палочек к концу созревания всех сыров, независимо от температуры второго нагревания и влажности, происходило за счет палочек со слабой способностью к кислотообразованию. Появление их связано с развитием остаточной микрофлоры пастеризованного молока.
В результате проведенных исследований установлено также, что температура второго нагревания и влажность сыра являются сильными регуляторами не только количества и состава микрофлоры при созревании сыра, но и ферментативной деятельности микробов.
В сырах, изготовленных при температуре второго нагревания 46°, с низкой влажностью, уже после прессования микрофлора сыра была представлена преимущественно молочнокислыми бактериями с подавленной способностью к кислотообразованию, хотя в состав закваски, которую применяли для выработки этого сыра, входило 87,5% штаммов с сильной способностью к кислотообразованию (см. рис. 4).
Наоборот, сыры с более высокой влажностью, выработанные при более низкой температуре второго нагревания (43 и 40°) отличались очень высоким содержанием молочнокислых бактерий, энергичных по кислотообразованию (после прессования 95—97%, а через 30 дней хранения 80—82%), причем количество их было выше, чем в исходной закваске.
Это свидетельствует, что сыр является средой весьма благоприятной для жизнедеятельности бактерий. В результате можно сделать заключение, что при температуре второго нагревания 43°, хотя и наблюдается некоторое подавление микробиологических процессов, но ферментативная активность бактерий сохраняется полностью.
При изучении протеолитической активности молочнокислых бактерий, выделенных из сыров на разных стадиях их созревания, выявлено, что молочнокислые палочки и стрептококки, обладавшие более высокой способностью к кислотообразованию, одновременно являлись и более активными по протеолизу.
Например, у молочнокислых палочек — энергичных кислотообразова- телей (128—180° Т через 48 часов) —отмечена полная корреляция между силой кислотообразования и протеолиза. Эти штаммы обладали наибольшей протеолитической активностью.
Среди молочнокислых палочек слабых кислотообразователей примерно 75% штаммов оказались сильными по протеолитической активности и только 25%—совсем слабыми в этом отношении.
У молочнокислых стрептококков, выделенных из сыра (как Sir. laclis. так и Sir. diacetilactis), отмечалась большая степень совпадения между способностью к кислотообразованию и протеолизу. Наибольшая протеолитическая активность в молоке у молочнокислых палочек выражалась числом 30 (алкалиметрический метод), а у стрептококков — 10—13.
В процессе созревания сыров биохимические свойства культур изменялись. Ароматообразующие стрептококки, выделенные из сыров после прессования, обладали довольно высокой способностью к продуцированию летучих кислот и одновременно они образовывали углекислый газ. В сырах через 30 и 75 дней культуры продуцировали меньше летучих кислот и многие из них утрачивали способность к газообразованию.
Ни в одном случае в исследованных сырах мы не обнаружили бактериофага. Все штаммы (4800), выделенные из сыра, за редким исключением, были фагорсзистентными.
Изменение активной кислотности при созревании сыра соответствует протекающим в нем микробиологическим процессам и последующим биохимическим изменениям сырной массы (табл. 11).
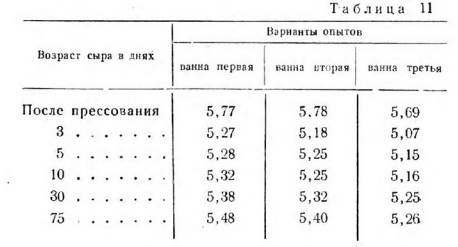
С увеличением влажности и понижением температуры второго нагревания повышается активная кислотность сыра, которая достигает максимальной величины через 3 дня после прессования. В этих сырах она находится на более высоком уровне до конца их созревания, что является одним из факторов, действующих на биохимические процессы распада параказеина (табл. 12).
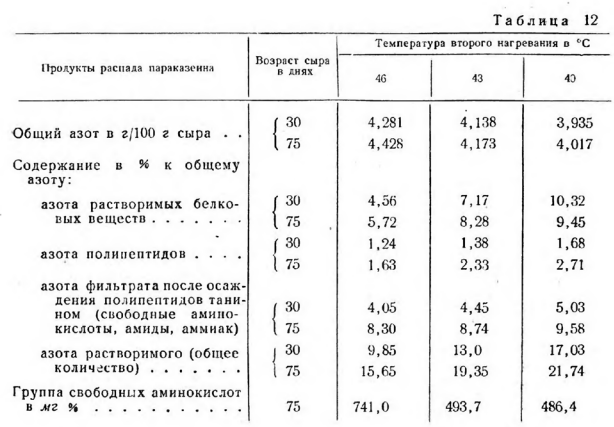
Количество азота растворимых белковых веществ и полипептидов уменьшается с понижением влажности сыра и повышением температуры второго нагревания. Разница в количестве азота раствормых белковых веществ в сырах, полученных из первой и третьей ванн, составляла 3,73% к общему азоту и полипептидов — 1,08 %.
При сравнении содержания азота в фракциях сыров, выработанных в условиях одинаковой температуры второго нагревания и отличающихся по влажности, можно заметить, что разница для растворимых белковых веществ составляет только 0,17% и полипептидов — 0,20%. Следовательно, влияние температуры на подавление развития молочнокислой микрофлоры при выработке сыра явилось решающим в определении интенсивности протеолиза параказеина.
Температура второго нагревания и связанная с ней влажность сыра оказали влияние на общий объем распада белковых веществ и усилили биохимические процессы образования свободных аминокислот (табл. 13).
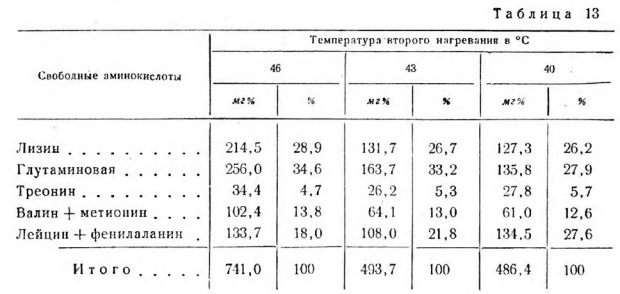
В сырах, выработанных при температуре второго нагревания 46°. влажностью 38,8% содержится на 52% больше свободных аминокислот, чем в сырах, выработанных при температуре второго нагревания 40° влажностью 44,7%. С повышением температуры второго нагревания увеличивается относительное количество глутаминовой кислоты и уменьшается сумма лейцина и фенилаланина. Это характерно для всех опытов, в которых изменялась температура второго нагревания.
Состав обезжиренного нерастворимого остатка сыров, выработанных при различных температурах второго нагревания, также изменяется (табл. 14).

В сырах с низкой влажностью, полученных при высокой температуре второго нагревания, нерастворимый остаток через 30 дней созревания сыра на 93,8% состоит из параказеина и через 75 дней — на 95,6%. В данном случае уменьшается количество промежуточных продуктов распада параказеина, которые входят в состав нерастворимого остатка, за счет чего повышается относительное содержание параказеина. В сырах с высокой влажностью, полученных при низкой температуре второго нагревания, содержание параказеина соответственно равно 92,2 и 90,9%, т. е. его содержание уменьшается вследствие увеличения нерастворимых продуктов его распада. Характер изменения состава нерастворимого остатка влияет на увеличение аминокислот в водной фазе сыра (см. табл. 13).
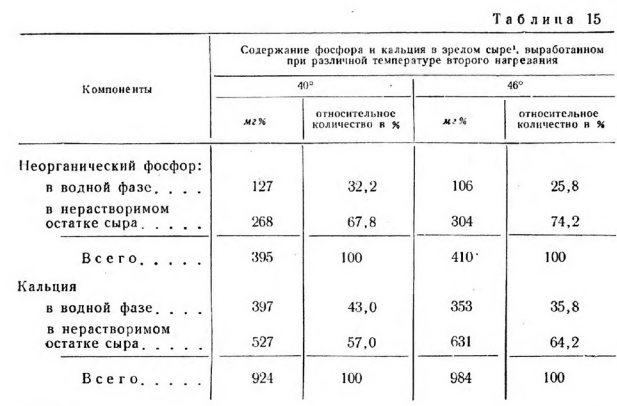
С увеличением влажности сыра содержание неорганического фосфора в его водной фазе увеличивается, а в нерастворимом остатке — уменьшается (табл. 15).
Та же закономерность наблюдается и в содержании кальция. Основная масса кальция и неорганического фосфора находится в нерастворимом остатке сыра. Относительное содержание их в водной фазе с повышением влажности сыра увеличивается, а в нерастворимом остатке уменьшается. Это явление объясняется величиной активной кислотности и более интенсивным распадом фосфорнокальциевого комплекса параказеина при созревании влажных сыров. Общее же количество этих неорганических компонентов с повышением влажности уменьшается, так как оно больше зависит от концентрации параказеина, чем от количества влаги.
В водной фазе на единицу фосфора приходится больше кальция, чем в нерастворимом остатке. Причем это отношение в более влажных сырах находится на менее высоком уровне, что обусловлено их активной кислотностью. Судя по составу неорганического фосфорнокальциевого соединения нерастворимого остатка, можно сказать, что фосфорнокальциевый компонент параказеина состоит преимущественно из фосфорнокальциевой соли. В составе же водной фазы сыра наряду с фосфатами содержатся значительные количества других кальциевых солей.
Решающую роль в биохимических процессах созревания сыра играют бактериальные ферменты, выделяемые микроорганизмами как во время их жизнедеятельности, так и при автолизе клетки. Эти процессы не ограничиваются только явлениями протеолиза белковых веществ, так как применением протеаз животного происхождения легко можно достигнуть высокой степени распада параказеина, но нельзя получить типичного вкуса сыра.
При созревании сыра происходят окислительно-восстановительные процессы, тесно связанные с деятельностью анаэробных дегидраз. Выделение свободной углекислоты и аммиака является следствием окислителыю-восстановительного процесса, сопровождающегося дезаминированием и декарбоксилированием азотистых соединений сыра.
В анаэробных условиях дегидразы способны восстанавливать метиленовую синь, причем скорость обесцвечивания пропорциональна активности этих ферментов.
На протяжении всего процесса созревания сыра активность дегидраз уменьшается, причем наиболее интенсивно в сырах с более высокой температурой второго нагревания (табл. 16).
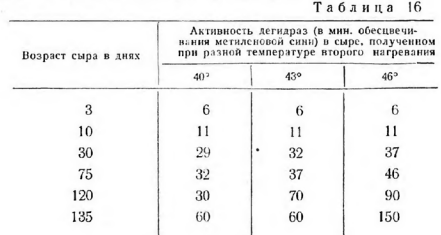
Скорость обесцвечивания метиленовой сини анаэробными дегидразами определяется ферментативной активностью как живых бактерий, так и автолизированных. Как показали микробиологические исследования, в сырах с повышенной влажностью (температура второго нагревания 40°) физиологическая активность микрофлоры сохраняется дольше, чем в сырах с пониженной влажностью (температура второго нагревания 46°). На основании этого можно предполагать, что разница между дегидразной активностью сыров с повышенной и пониженной температурой второго нагревания объясняется ферментативной активностью живых клеток.
Молочнокислые бактерии являются носителями ферментов, которые обусловливают созревание сыра. Можно полагать, что их концентрация будет пропорциональна концентрации бактериальной массы. О бактериальной массе в сыре обычно судят по количеству бактерий. Но это число не полностью соответствует объему микробной массы, так как наряду с размножением микрофлоры происходит ее автолиз. Необходимо также иметь в виду, что молодые и старые бактериальные клетки обладают разной ферментативной активностью.
В связи с этим нами предпринято предварительное определение в сыре пентоз, структурных элементов нуклеиновых кислот, которые являются составной частью микробной клетки. Содержание нуклеиновых кислот колебалось от 22 до 28% сухого вещества в молодой клетке и от 9 до 10%—в шестисуточпых культурах. В молочнокислых бактериях нуклеиновых кислот содержится около 13%. В табл. 17 приведены результаты исследований пентоз на различных стадиях созревания сыра.
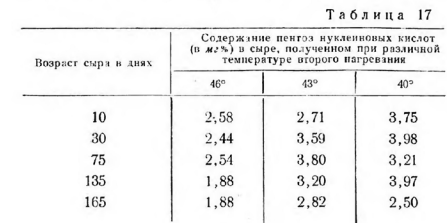
Количество пентоз с понижением температуры второго нагревания увеличивается, а с возрастом сыра—уменьшается, причем наиболее заметно в сырах при повышенной температуре второго нагревания.
Органолептические свойства сыров каждого варианта резко отличаются. Вкус сыров с влажностью после прессования 38—40%. выработанных при температуре второго нагревания 46°, хороший и удовлетворительный, но с довольно сильным оттенком пряности. Последнее не соответствовало типичным признакам костромского сыра и потому оценка его была снижена (средний балл 39,2). Особенно неудовлетворительной была консистенция сыра—плотная и твердая (средний балл 21.8). Рисунок сыра обычно был нормальным, но в некоторых случаях недостаточно развитым (средний балл 9,0).
Сыры с влажностью после прессования выше 43,5%, полученные при температуре второго нагревания 40°, имели хорошую консистенцию (средний балл 24), но вместе с этим появились существенные недостатки во вкусе и запахе (средний балл 38,8). В сырах ощущался горьковатый и кисловатый привкусы, что привело к снижению их балловой оценки, у многих образцов сыра был неравномерный рисунок (средний балл 8,6).
Сыры с влажностью 41—43% (температура второго нагревания 43°) преимущественно имели хороший вкус (средний балл 40,0). Однако их консистенция уступала сырам с высокой влажностью (средний балл 23,0). Сыры этого варианта получили наиболее высокую общую оценку по сравнению с другими.
Общая балловая оценка сыров, полученных при температуре второго нагревания 46°, составляла 89,8 балла, при температуре 43° — 92,0 и температуре 40° — 91,2 балла.
В результате исследования сыров с различной 'влажностью, полученных в различных температурных условиях второго нагревания, можно сделать следующие выводы:
- Температура второго нагревания даже при таких сравнительно незначительных колебаниях, как 40, 43 и 46°, является регулятором микробиологических процессов в сыре, определяющим не только количество микрофлоры, ее состав, но и ферментативную активность бактерий. С повышением температуры второго нагревания до 46° количество бактерий и их биохимическая активность резко понижаются.
- При повышенной температуре второго нагревания (46°) и низкой влажности сыров после прессования (38—40%) подавляется развитие ароматообразующих бактерий типа Str. diacctilactis, но активизируется развитие молочнокислых палочек и почти не изменяется количество Str. paracitrovorus.
- В процессе созревания сыра с приближением кондиционной зрелости (75 дней) ферментативная активность бактерий понижается и в микрофлоре зрелых сыров начинают преобладать малоактивные формы стрептококков и палочек с резко ослабленной способностью к кислото- образованию, продуцированию летучих кислот и углекислого газа.
- Характер развития микробиологических процессов под влиянием повышенной температуры второго нагревания определяет объем и направление протеолиза белковых веществ сыра. С повышением температуры уменьшается количество растворимых белковых веществ и полипептидов и увеличивается количество свободных аминокислот. При этом их соотношение изменяется таким образом, что доля лейцина и фенилаланина уменьшается, а глутаминовой кислоты увеличивается.
- С повышением влажности увеличивается активная кислотность сыров, которая находится на более высоком уровне на протяжении всего процесса созревания.
- Активность анаэробных дегидраз резко уменьшается при созревании сыра, однако скорость этого процесса зависит от его влажности и температуры второго нагревания. В сырах с повышенным содержанием влаги эта активность находится на более высоком уровне, чем в менее влажных, что связано с более длительным сохранением ферментативной активности бактерий.
- Количество пентоз, компонентов нуклеиновых кислот, с повышением влажности увеличивается и в некоторой степени соответствует росту микрофлоры. Наблюдается тенденция заметного их уменьшения при созревании сыров с пониженной влажностью и стабильное содержание в сырах с высокой влажностью. Дегидразная активность, по-видимому, связана с более длительным сохранением ферментативной активности микрофлоры сыра.
- С повышением влажности сыра после прессования происходит значительное улучшение его консистенции. Однако при влажности сыра 43—46% во вкусе появляется горечь и излишняя кислотность, которые резко снижают его общую балловую оценку.
- При низкой влажности сыров (до 40%) и повышенной температуре второго нагревания (46°) появляется пряный вкус, не свойственный данной группе сыров, консистенция его становится непластичной, грубой.
- Наилучшую оценку из данной серии опытов получили сыры с. влажностью 41—43%, выработанные при температуре второго нагревания 43°. В этой группе 80% сыров было с хорошими вкусовыми достоинствами и 20% —с удовлетворительными. Консистенция же их уступала консистенции сыров с более высокой влажностью. Таким образом, одним повышением влажности сыра после прессования не всегда можно достигнуть общего улучшения его качества. Необходимо учитывать другие факторы, под влиянием которых формируются органолептические свойства. К ним относится активная кислотность, зависящая от содержания в водной фазе сыра молочного сахара и развития молочнокислого процесса до его посолки, в особенности при обработке сырной массы в ванне.
