9.1. БИОТЕХНОЛОГИИ, ИНФОРМАЦИЯ
И САМООРГАНИЗАЦИЯ ПРИРОДЫ
Биотехнологии являются информационными технологиями. В большинстве случаев современные биотехнологии – это преобразования (операции), проводимые с генетической информацией организмов, являющихся самоорганизующимися живыми системами. Информатика и самоорганизация – относительно новые отрасли знаний. Их пока изучают не во всех учебных заведениях, выпускники которых работают по биотехнологиям. Рассмотрим кратко самые важные для биотехнологов сведения по информации и самоорганизации.
Система жизнеобеспечения организмов состоит из трех основных подсистем: обмена веществ, энергообмена и информационных или управляющих процессов. Аналитическое выделение этих, физически неразделимых подсистем, позволяет выявить их соподчиненность и характерные особенности. Подсистема обмена веществ в сообществах организмов, экосистемах частично или полностью замкнута. Она принципиально не ограничивает развитие организмов.
В популяциях, способных к размножению, генетическая информация при благоприятных условиях может неограниченно долго циркулировать, переходя от поколения к поколению. В анабиозном состоянии генетическая информация может сохраняться очень долго. Подсистема информационных или управляющих процессов также принципиально не ограничивает развитие организмов.
Энергия, использованная организмом, большей частью рассеивается, деградирует и повторно не может использоваться подобными организмами. Подсистема энергообмена разомкнута, однонаправлена. Свободная энергия, доступная для организмов, принципиально ограничивает их развитие. Биотехнологу важно знать, что подсистемы информационная и обмена веществ подчинены подсистеме энергообмена организма. Эволюционное развитие информационной подсистемы и подсистемы обмена веществ направлены в основном на совершенствование подсистемы энергообмена.
Величины (понятия) количества информации, традиционно используемой в современной «теории информации», являющейся по существу теорией связи, недостаточно для характеристики биологической (генетической) информации. Наряду с количеством информации для применения биологической информации особо важно оценивать ее ценность (качество). Этот показатель информации определяют посредством представления о достижении определенной цели. Информация ценна в той мере, в какой она приближает к достижению желаемой (требуемой) цели. В связи с отмеченной выше соподчиненностью информационной подсистемы подсистеме энергообмена качество биологической информации в общем случае количественно целесообразно оценивать посредством учета изменения энергетической эффективности биологического объекта, которым получена определенная информация. Следует отметить, что ценность информации зависит от цели, которую преследует ее получатель (рецептор).
Ранее прилагательные «ценная» и «осмысленная» применительно к информации использовались без соответствующих определений в расчете на то, что они понятны интуитивно. Понятие ценность информации является весьма важным, центральным как в биотехнологии, так и в развиваемой современной теории информации. Ценность информации определяется мерой приближения к достижению цели при использовании полученной информации. Если преследуемая цель надежно достижима несколькими путями, то ценность информации можно определять уменьшением материальных, энергетических или временных затрат благодаря использованию данной информации.
В случае если достижение цели вероятно, но не обязательно, можно использовать следующий критерий ценности информации:
V = Log2(P/p) (9.1)
где р – вероятность достижения цели до получения информации; Р – тоже, после получения информации (Д.С. Чернавский, 2004).
В соответствии с уравнением (9.1), ценность информации зависит от величины р, т. е. от того, какой предварительной априорной информацией уже располагает рецептор. Если она отсутствует, то априорная вероятность во всех возможных вариантах (n) одинакова и равна p = 1/n. В этом случае величина р выполняет роль нормировочного множителя. Если после получения информации в этом случае цель достигается наверняка (Р = 1), то ценность этой информации максимальна и равна V= Vmax = log2n, при этом ценность информации совпадает с максимальным количеством информации, традиционно определяемой в теории связи, и при этом ценность информации можно понимать как количество ценной информации.
В традиционной теории информации (теории связи) величина энтропии, отображающая второе начало, получила необоснованное распространение под названием «информационная энтропия». На основе положений самоорганизации выявлена ошибочность термодинамической трактовки понятия (определения) информации посредством этого термина. Ошибочным также является использование термина «негэнтропия» и утверждение о том, что «информация есть негэнтропия».
Из большого количества понятий (определений) информации, используемых различными авторами, наиболее совершенным, исходя из положений самоорганизации, принято определение информации, предложенное Г. Касглером (Чернавский, 2004). Это определение следующее: информация есть случайный и запомненный выбор одного варианта из нескольких возможных и равноправных. Такое определение информации не имеет никакого отношения к понятиям «информационная энтропия» и «негэнтропия».
Наиболее важным достижением фундаментальной науки в последние десятилетия является развитие ее новой отрасли – самоорганизации (синергетика, неравновесная термодинамика, динамика сложных нелинейных систем). Исходя из самоорганизации, наряду с традиционным делением природы на живую и неживую, не менее важно делить ее на самоорганизующуюся (неравновесную) и несамоорганизующуюся (равновесную). Самоорганизующиеся физико-химические системы по своим важным свойствам, например энергопреобразующим, более близки к живым, чем к равновесным физико-химическим. Главным принципом этой новой отрасли науки является принцип энергетической экстремальности самоорганизации и прогрессивной эволюции. Этот принцип состоит из второго начала термодинамики и противоположного ему по сущности закона выживания. Второе начало приложимо только к равновесным (несамоорганизующимся) системам, а закон выживания – к самоорганизующимся (неравновесным).
Принцип энергетической экстремальности самоорганизации, представляющий диалектическое единство противоположной сущности двух названных законов, позволил решить столетние проблемы равновесной термодинамики, в том числе проблему «вопиющего» противоречия между эволюцией природы по второму началу термодинамики и теорий биологической эволюции (дарвиновской, синтетической). В самоорганизации выявлен и второй важный принцип – принцип подчинения синергетики. Сущность его в том, что из большого числа переменных, характеризующих систему, выбирают одну наиболее быстро изменяющуюся переменную (от которой в наибольшей мере зависит состояние системы), называемую переменной порядка. Затем определяют несколько (2–3) параметров управления и при помощи их и переменной порядка определяют состояние системы и управляют ею.
Самоорганизующиеся живые организмы избирательно потребляют из среды доступную свободную энергию, накапливают ее и используют на свои жизненные процессы. Самоорганизующиеся системы (структуры) называют еще «диссипативными», так как они живут за счет рассеяния, диссипации энергии. Все этапы эволюционного развития самоорганизующейся природы (физико-химический, биологический, социальный) имеют, в соответствии с принципом энергетической экстремальности самоорганизации, общую энергоэкономную (высоко энергоэффективную) направленность живых систем. Важным принципом биологической информации является максимум полезной (ценной) информации в минимуме массы и объема, а также минимум затрат энергии на сохранение и функционирование. Этот принцип позволяет биотехнологу составить представление о том, в каком направлении будет эволюционировать созданный им объект на основе генной инженерии.
9.2. БИОЭНЕРГЕТИКА НА МОЛЕКУЛЯРНОМ УРОВНЕ
Биоэнергетика изучает механизмы и законы преобразования энергии в процессах жизнедеятельности организмов. Слово «биоэнергетика» впервые использовал для названия своей книги американский биохимик (венгерского происхождения) А. Сент-Дьерди в 1956 г. Официально название «биоэнергетика» за этим разделом знаний было закреплено на симпозиуме биохимиков в Италии в 1967 г. Началом исследований в биоэнергетике являются работы немецкого врача Ю.Р. Майера (1841 г.), который открыл закон единства сохранения и превращения энергии в живой и неживой природе. Вначале энергообмен живой природы изучали на организменном и надорганизменном уровнях. С развитием биохимии биоэнергетику начали развивать на молекулярном уровне.
Живые организмы отличаются большой метаболической гибкостью и энергоэкономностью в соответствии с принципом энергетической экстремальности самоорганизации и прогрессивной эволюции. В известных пределах они могут изменять свой метаболизм в зависимости от того, какие питательные вещества (или иной вид доступной свободной энергии) имеются в окружающей срдде. Например, бактерия Е. coli, являющаяся хемоорганотрофом, может использовать в качестве единственного источника углерода не только глюкозу, но и другие сахара, глицерин, аминокислоты и даже такие простые соединения, как этиловый спирт и уксусная кислота.
Любое проявление жизни связано с затратами энергии. Организмы и их составляющие – самоорганизующиеся диссипативные системы, живущие за счет потребления из окружающей среды свободной энергии. Основные биоэнергетические процессы – фотосинтез (хемосинтез), дыхание, брожение – не случайно консервативны в эволюционном отношении. Возникнув у самых древних организмов, они в неизменном виде сохранились в современных (в высших растениях, организме человека). Это свидетельствует об их безальтернативности и фундаментальности, а также ведущей роли энергообмена в жизнеобеспечении организмов. Принято считать, что к превращениям энергии в организме применимы законы термодинамики, физики и химии.
Однако самоорганизующаяся сложность и специфичность биологических структур, а также протекающих в них самоорганизующихся процессов обусловливают ряд глубоких различий между биоэнергетикой и энергетикой не самоорганизующейся природы, в частности, технической энергетикой. Фундаментальная особенность биоэнергетики состоит втом, что организмы являются открытыми самоорганизующимися системами. Они функционируют только в условиях постоянного обмена веществом и энергией с окружающей средой. Преобразование энергии в таких системах принципиально отличается от преобразований ее в тепловых машинах. Основополагающее для равновесной термодинамики понятие о равновесном состоянии заменяется в биологии представлением неравновесной термодинамики о стационарном состоянии. Второе начало термодинамики (принцип возрастания энтропии) заменяется на противоположный ему по сущности закон выживания или принцип минимизации удельного производства внутренней энтропии Пригожина, в соответствии с которым энтропия в самоорганизующейся системе не возрастает, а уменьшается.
Вторая важнейшая особенность биоэнергетики связана с тем, что процессы преобразований энергии происходят при отсутствии перепадов температуры, изменений давления и объема. Из-за этого переход теплоты в работу в организме невозможен. В процессе эволюции организмы выработали специфические механизмы прямого преобразования одного вида свободной энергии в другой, минуя ее переход в теплоту. В организме лишь небольшая часть освобождающейся энергии превращается в теплоту и теряется. Большая ее часть преобразуется в форму свободной химической энергии особых соединений, в которых она чрезвычайно мобильна. Она может при постоянной температуре превращаться в иные формы, в частности, совершать работу или использоваться для биосинтеза, чем обеспечивается высокая эффективность биоэнергетических процессов. Например, КПД работы мышц достигает 30 %. Биоэнергетические процессы на молекулярном уровне протекают одновременно с биохимическими процессами метаболизма.
Обмен веществ состоит из двух процессов: катаболизма и анаболизма (В.П. Скулачев, 1989). При катаболизме происходит ферментативное расщепление сравнительно крупных молекул пищи – углеводов, жиров, белков, происходящее преимущественно за счет реакций окисления. При этом образуются более мелкие молекулы, например молочной кислоты, СО2, уксусной кислоты, аммиака или мочевины. Катаболизм сопровождается выделением свободной энергии изсложных структур крупных пищевых молекул и запасается в более доступной форме в виде фосфатных связей в аденозинтрифосфатах (АТФ). Катаболические процессы являются экзергоническими. Они идут с уменьшением свободной энергии. Анаболические процессы – эндергонические – протекают с увеличением свободной энергии.
Анаболизм представляет собой ферментативный синтез сравнительно крупных молекул клеточных компонентов (например, полисахаридов, нуклеиновых кислот, белков или жиров) из простых составляющих. Усложнение структур и увеличение размеров молекул при синтезе сопровождается затратами свободной энергии, запасенной в фосфатных связях АТФ. В клетках катаболизм и анаболизм протекают одновременно. В результате последовательных ферментативных реакций, при которых происходит соответственно разрушение (катаболизм) или синтез (анаболизм) ковалентного остова данной биомолекулы, образуются промежуточные продукты. Их называют метаболитами, а всю цепь превращений объединяют названием промежуточный метаболизм. Каждая из ферментативных реакций промежуточного метаболизма сопровождается соответствующим превращением энергии. На некоторых этапах катаболизма химическая энергия метаболитов запасается (в форме фосфатных связей), а на определенных этапах анаболизма она расходуется. Эту особенность метаболизма принято называть сопряжением энергии. Промежуточный метаболизм и сопряжение энергии – взаимосвязанные и взаимозависимые понятия. Поэтому при изучении метаболизма необходимо анализировать как реакции, в результате которых изменяются ковалентные структуры предшественников и образуется продукт, так и энергетические изменения, сопровождающие эти превращения.
АТФ называют «разменной энергетической монетой». Она – важнейшее вещество, играющее для всего живого мира роль почти единственного трансформатора и передатчика энергии. АТФ может расщепляться до аденозиндифосфорной кислоты (АДФ) или до аденозининмонофосфорной кислоты (АМФ). Отщепление от АТФ конечной фосфатной группы (гидролиз) протекает по формуле АТФ + Н2О → АДФ + фосфат. Эта реакция сопровождается выделением свободной энергии ∆F. Если эта реакция протекает при концентрации всех реагентов и продуктов в 1,0 моль при 25 °С и рН 7,0, то свободная энергия АДФ оказывается меньше свободной энергии АТФ на 7 ккал/моль. В клетке это изменение свободной энергии больше – 12 ккаЛ/моль.
Значения ∆F в клетке для реакции АТФ → АДФ выше, очевидно не случайно, чем у большинства реакций гидролиза. Это различие очевидно обусловлено энергоэффективным синергизмом процессов клетки в соответствии с принципом энергетической экстремальности самоорганизации и прогрессивной эволюции. Связи третьей (конечной) и второй фосфатных групп в молекуле АТФ называют макроэргическими.
Под энергией этих связей в биоэнергетике понимают не действительную энергию ковалентных связей между атомами фосфора и кислорода (или азота), как это принято в физической химии, а лишь разность между свободной энергией (∆F) исходных реагентов реакций гидролиза АТФ или других аналогичных реакций и конечных продуктов этих реакций. «Энергия связи», в точном понимании этого термина, не локализована в данной связи, а характеризует реакцию в целом как самоорганизующийся процесс. Энергия макроэргических связей АТФ – универсальная форма запасания свободной энергии для всей живой самоорганизующейся природы: от микроорганизмов до человека. Все преобразования энергии в процессах жизнедеятельности происходят посредством аккумуляции энергии в этих связях, а ее использование – при их разрыве. Порции энергии макроэргических связей обеспечивают высокую биоэнергетическую эффективность живой самоорганизующейся природы, тождественно этой роли кванта действии, открытого более ста лет назад М. Планком, в квантовой физике. Макроэргическую связь ∆F можно назвать биологическим квантом действия.
9.3. БИОТЕХНОЛОГИИ
И ЭНЕРГЕТИЧЕСКАЯ ПРОБЛЕМА
Начало 70-х годов XX столетия ознаменовалось выявлением мирового энергетического кризиса. За период с 1972 по 1982 гг. мировые цены ца нефть и нефтепродукты возросли в 10 раз. Этот рост продолжается и в настоящее время (И.И. Свентицкий, 1982, 2001). Практическая причина этого – приближение исчерпаемости невозобновляемых горючих ископаемых – основного первичного источника промышленной энергетики и транспорта. Теоретически эту причину позволяет объяснить принцип энергетической экстремальности самоорганизации. В РФ принята долгосрочная (до 2010 г.) федеральная программа «Энергоэффективная экономика», которой предусмотрено повышение эффективности использования энергоресурсов во всех министерствах и ведомствах. Приоритетным мировым направлением научных исследований стало выявление возобновляемых альтернативных источников энергии и высокоэффективное использование энергетических ресурсов (энергосбережение).
В чем естественнонаучная причина энергетической проблемы? Новейшие положения самоорганизации (неравновесной термодинамики, синергетики, динамики сложных нелинейных систем) позволяют однозначно ответить на этот вопрос. Наиболее важным достижением самоорганизации является обоснование принципа энергетической экстремальности самоорганизации и прогрессивной эволюции (И.И. Свентицкий, 2001). Этот принцип состоит из второго начала равновесной термодинамики и противоположного ему по сущности закона, названного законом выживания. Начало и закон логически концептуально объединены динамической зеркальной симметрией. Второе начало объясняет преобразование энергии в равновесных (несамоорганизующихся) замкнутых системах, например тепловых машинах, в которых энергия преобразуется благодаря использованию температурного градиента. Оно выполнило и выполняет важную роль в прогрессивном развитии традиционной градиентной энергетики.
Однако, необоснованное распространение приложения второго начала равновесной термодинамики к высоко энергоэффективным природным самоорганизующимся (открытым, неравновесным) системам обусловило ряд проблем фундаментальной науки. Этим обусловлен также своеобразный теоретический запрет на развитие нетрадиционной энергетики, основанной на использовании самоорганизующихся процессов, в частности фазовых переходов. Принцип энергетической экстремальности самоорганизации и входящий в него закон выживания позволили решить эти проблемы и выявить естественненаучную причину возникновения энергетической проблемы. В соответствии с этим принципом энергетической экстремальности самоорганизации и законом выживания все этапы эволюции природы (физико-химический, биологический, социальный) имеют общую энергоэффективную направленность. Природа на протяжении всей своей эволюции «решает проблему энергосбережения». В сознательной деятельности человек, не учитывая этот закон, допустил энерго- и ресурсорасточительство. Этим и обусловлено возникновение энергетической проблемы. Человек и его общество, как и все иные части природы, подчинены принципу энергетической экстремальности самоорганизации и прогрессивной эволюции. Проблема высокоэффективного использования доступной свободной энергии не преходяща. Человек неизбежно должен ее решать на протяжении всего своего существования.
В связи с этим не случайно наиболее важным фактором, определяющим уровень развития общества, является его энерговооруженность. Она определяет все стороны общественного производства. Интенсивное развитие всех отраслей народного хозяйства связано с неизбежной необходимостью увеличения потребления энергии. Ярким примером является интенсификация сельскохозяйственного производства. Получение продовольствия становится все более энергоемким. В настоящее время на 1 кал, запасенной в продовольствии, затрачивают более 10 кал техногенной энергии. Потребление энергии на одного человека в развитых странах за исторический период развития возросло более чем в 100 раз и продолжает расти.
В решении энергетической проблемы важную роль могут выполнить биотехнологии. Основной возобновляемый источник энергии – энергия оптического диапазона солнечного излучения. По прогнозам только техническое преобразование этой энергии к 2020 г. в общем энергетическом балансе США должно составить 26 %. Большие возможности в самообеспечении сельского хозяйства энергией открывает биоконверсия солнечной энергии растениями с последующим получением моторных топлив (жидкого, газообразного) из биомассы. В последние годы в ряде стран, например в Чехии, построены крупнотоннажные заводы по производству дизельного топлива из масла семян рапса. При урожае семян рапса 30 ц/га и выше в условиях Чехии такое топливо конкурентоспособно с дизельным топливом, получаемым из нефти. Биотоплива, получаемого с урожая рапса, выращенного на 1 га, достаточно для обеспечения машинных технологий по выращиванию растений на 8–10 га.
В энергетической культуре рапса используют сорта и гибриды, выведенные для пищевых и кормовых целей. Эффективность энергетических культур можно существенно повысить выведением специализированных сортов и гибридов на основе биотехнологий не только рапса, но и других видов растений, в том числе и микроводорослей, например хлореллы. Прогнозное расчетное определение эффективности выводимых энергетических сортов и гибридов можно надежно проводить, используя компьютерную технологию (см. 9.7).
Потребительская стоимость энергии зависит от ее формы. На рис. 9.1 приведены способы конверсии различных форм энергии (в последовательности их ориентировочной иерархической ценности). Существующие технологические способы изображены сплошными линиями, разрабатываемые и возможные в будущем – пунктирными. Каждый этап преобразования энергии охарактеризован ориентировочным значением КПД. Отметим, что КПД преобразования энергии в процессе фотосинтеза растений в 2,5 раза на схеме занижен в связи с неточным определением эксергии солнечного излучения в отношении фотосинтеза растений (И.И. Свентицкий, 1982).
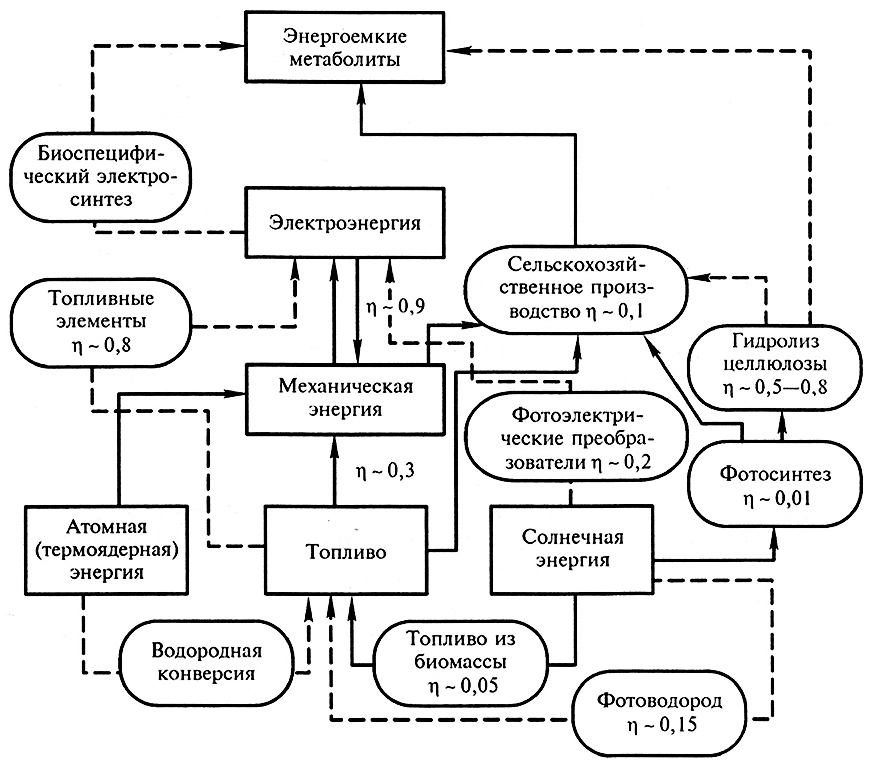
Рис. 9.1. Схема процессов преобразований энергии:
сплошные линии – реализованные; пунктирные – разрабатываемые и возможные в будущем
(из книги С.Д. Варфоломеева «Конверсия энергии биокаталитическими системами». М. Изд-во МГУ, 1981).
Электрическая энергия по многим показателям является наиболее удобной и чистой формой энергии (ее работоспособность – эксергия – составляет практически 100 %). В то же время наибольшую ценность представляют энергоемкие метаболиты, обеспечивающие локальные биоэнергетические процессы живых организмов. Значительная, если не большая часть всей техногенной энергии затрачивается человечеством на получение, транспортирование и распределение продовольствия.
Роль топлива как источника и накопителя (аккумулятора) энергии будет очевидно возрастать. В связи с этим привлекательна концепция водородной энергетики. Водород по оценкам многих экспертов является наиболее перспективным веществом для использования в качестве синтетического топлива. Он обладает достаточно высокой энергоемкостью. По этому показателю он превосходит все другие соединения, которые могут служить в качестве топлива: природный газ – в 2,5 раза, жидкие углеводороды нефти – в 3,3 раза, метанол – в 6,6 раза, целлюлозу – в 8,3 раза. Водород экологически чистое топливо, не вызывающее загрязнений окружающей среды. Основной, практически единственный продукт его сгорания – вода. Побочным экологически негативным продуктом сгорания водорода в воздухе является оксид азота, который образуется в ничтожных количествах.
Принципиальное значение для успешного применения водородной энергетики имеет выбор первичного источника энергии для получения водорода, а также его безопасное хранение и транспортирование. Положительной особенностью водорода является возможность высокоэффективного преобразования его энергии в электроэнергию при помощи топливных элементов (электрохимических генераторов), КПД которых достигает 80 %. Такой высокий КПД обусловлен тем, что в топливных элементах химическая энергия непосредственно преобразуется в электрическую. Аналогично преобразованию ее в организмах в другие необходимые виды энергии, в том числе и в электрическую. Альтернативны для получения водорода в настоящее время два вида первичной энергии: атомная и солнечная.
Недостатками использования для этой цели солнечной энергии являются: низкая плотность потоков энергии, отсутствие в настоящее время экономически конкурентоспособной технологии ее использования. Большие надежды в исследованиях по водородной энергетике возлагают на возможность принципиального (в несколько раз) уменьшения затрат первичной энергии на получение водорода при электролизе воды. Предполагается, что воздействие на воду током высокой (сверхвысокой) частоты, обеспечивающей резонанс с частотой собственных колебаний молекул воды, во много раз может уменьшить затраты энергии на выделение водорода из воды в процессе электролиза. Явления резонанса играют важную роль в процессах самоорганизации, обеспечивая высокую эффективность преобразований энергии. Практическая возможность резонансного уменьшения затрат энергии на выделение водорода вполне реальна.
9.4. НЕОБХОДИМОСТЬ УЧЕТА
БИОЭНЕРГЕТИЧЕСКИХ ПРОЦЕССОВ В БИОТЕХНОЛОГИЯХ
Из общего количества энергии, используемой при производстве сельскохозяйственной продукции, техногенная (промышленная) энергия составляет не более 3–4 %. Главная ее часть приходится на природную энергию – электромагнитную энергию солнечного света (И.И. Свентицкий, 1982,2001). Эту энергию первоначально преобразуют и запасают растения, а затем используют другие организмы (человек, животные, микроорганизмы) и их сообщества. Большие количества природной энергии накоплены в органическом веществе почвы, где она также преобразуется живыми почвенными организмами, которые формируют и поддерживают почвенное плодородие.
Конечная цель агро- и биотехнологий – получение определенного вида высококачественной продукции при низких энергетических и материальных затратах, а также негативном минимальном воздействии на природную среду (B.C. Шевелуха, 1992).
Сущность общебиологического закон биоэнергетической общей направленности структур и функций живых систем, названного законом выживания, состоит в том, что все элементы живой природы в своем развитии самопроизвольно направлены к наиболее полному (эффективному) использованию доступной свободной энергии. Под свободной энергией понимается та часть общей энергии, которая потенциально может быть использована данным элементом природы на свои процессы или преобразована в требуемый вид энергии. Важное практическое следствие этого закона – необходимость количественной оценки этой энергии на входе в любой ее преобразователь (потребитель), а также на выходе из него.
В селекции возникло и развивается направление по выведению энергоэкономных сортов (гибридов) растений, пород животных, штаммов микроорганизмов (B.C. Шевелуха, 1992). При такой селекции важно иметь прогнозную (расчетную) оценку на энергоэкономность исходного селекционного материала. Эта оценка должна учитывать конкретные экологические условия, в которых будут выращиваться новые сорта, гибриды, породы, штаммы.
Надежный анализ энергопреобразующих процессов как в технических, так и в биологических системах можно проводить на основе законов термодинамики и закона выживания. Законы термодинамики разрабатывались применительно к техническим преобразователям энергии – тепловым машинам. Эти законы применимы и для анализа преобразований энергии живыми системами, но их недостаточно для объяснения особенностей их развития и функционирования. Эти особенности можно объяснить на основе закона выживания – общей биоэнергетической направленности структур и функций самоорганизующихся живых систем.
9.5. ЗАКОНЫ И ОСНОВНЫЕ ПОНЯТИЯ ТЕРМОДИНАМИКИ
Обмен веществ в живых системах происходит благодаря энергетическим и информационным процессам. Он слагается из химических и физических реакций и подчиняется законам термодинамики.
Наиболее важными понятиями термодинамики являются: система, процесс, параметры состояния системы. Подсистемой понимают комплекс взаимосвязанных элементов (природных или созданных человеком), который способен потреблять, преобразовывать и накапливать энергию. Системы бывают открытыми, закрытыми и замкнутыми. Открытые системы обмениваются с внешней средой и веществом, и энергией. Закрытые системы обмениваются с внешней средой только энергией и не обмениваются веществом. У замкнутых систем нет обмена с внешней средой ни веществом, ни энергией. Природные системы, в том числе живые, являются открытыми. Различают самоорганизующиеся (неравновесные) и несамоорганизующиеся (равновесные, косные) системы.
Живые самоорганизующиеся системы способны самопроизвольно потреблять из внешней среды свободную энергию и за счет этого поддерживать свои внутренние интенсивные параметры (температура, давление, уровень концентрации, градиент вещества и энергии и др.), отличные от таковых внешней среды. Из-за этого их называют также неравновесными (диссипативными) системами. Косные системы не обладают такой способностью. Их интенсивные параметры самопроизвольно сравниваются с аналогичными параметрами внешней среды. Такие системы называют равновесными.
Необходимо учитывать первый и второй законы термодинамики. Согласно первому закону термодинамики, закону сохранения и превращения энергии, ее общее количество в замкнутой системе сохраняется неизменным. Энергия не исчезает и не возникает, а только преобразуется из одного вида в другой.
Второй закон термодинамики налагает определенные ограничения на возможные самопроизвольные превращения разных видов энергии. Все процессы в природе самопроизвольно протекают в одном направлении, а именно, в направлении снижения качества энергии, уменьшения ее работоспособности. Все градиенты (температура, давление и др.) самопроизвольно уменьшаются. Снижение качества энергии характеризуют ростом энтропии. Применительно к тепловым машинам эту величину называют приведенной теплотой. Она равна отношению количества теплоты к температуре, при которой эта теплота находится (содержится в системе или теплоносителе). Чем выше энтропия, тем ниже работоспособность этого вида энергии и тем меньше в этой общей энергии содержится свободной энергии, или эксергии.
9.6. СПОСОБЫ АНАЛИЗА ПРЕОБРАЗОВАНИЙ ЭНЕРГИИ
Известны три способа анализа энергопреобразующих процессов: по величине общей энергии, по величине энтропии и по величине свободной энергии, или эксергии. При анализе (балансе) общей энергии учитывается только первый закон термодинамики и не принимается во внимание второй ее закон, отсутствует учет качества (потенциальной превратимости) различных видов энергии. Этот метод анализа не точный и может привести к серьезным ошибкам.
В агроэнергетике необходимо проводить совместный анализ как технических преобразований энергии, так и ее биоконверсию – преобразование ее живыми организмами. Сельскохозяйственному производству основные энергетические ресурсы поставляет промышленная энергетика, в которой используют эксергетический анализ. Это благоприятствует применению эксергетического анализа как в агротехнологиях, так и в биотехнологиях. Аграрное производство – один из основных пользователей экологических ресурсов. В экологии до настоящего времени энергетический анализ проводят на основе баланса общей энергии, но уже разработаны положения эксергетического анализа для экологии и есть надежда, что в этой отрасли знаний будет применяться эксергетический анализ.
В экологии и сельскохозяйственном производстве первичными преобразователями основного вида энергии являются фотоавтотрофные растения. Они включают электромагнитную энергию оптического солнечного излучения во все жизненные процессы природы и общества. Рассмотрим эксергетический анализ при энергосберегающей оптимизации производства продукции растениеводства посредством агротехнологий и биотехнологий.
9.7. ЭНЕРГОСБЕРЕГАЮЩАЯ ОПТИМИЗАЦИЯ
ПРОИЗВОДСТВА ПРОДУКЦИИ РАСТЕНИЕВОДСТВА
НА ОСНОВЕ ЭКСЕРГЕТИЧЕСКОГО АНАЛИЗА
Анализ удобно начать с определения эксергии оптического излучения (света) в отношении фотосинтеза и формирования продуктивности растениями. Под этой величиной понимается та часть общей энергии излучения, поступающей к растениям, которая потенциально, при всех других благоприятных условиях, может быть использована ими на фотосинтез. Эту величину называют также свободной энергией в отношении фотосинтеза растений или фотосинтезной энергией.
Фотофизические и химические реакции фотосинтеза растений протекают при практически постоянных температуре и давлении. Эти параметры состояния, интенсивные для тепловых преобразователей, не оказывают существенного влияния на энергопреобразующие процессы живых организмов. Интенсивным параметром энергоносителя (излучения) в данном случае является энергия фотона – кванта излучения, которая определяется длиной его волны (А,). Чем больше длина волны излучения, тем меньше величина энергии его кванта – фотона (е), которая определяется выражением
ɛ = hγ, (9.2)
где h – постоянная Планка, равная 6,625 • 10-34 Дж • с;
γ – частота излучения.
В соответствии с законом квантовой эквивалентности число осуществленных фотохимических актов пропорционально не количеству подведенной общей энергии излучения, а числу поглощенных фотонов. Фотохимические элементарные акты в фотосинтезе растений могут осуществлять только фотоны определенной энергии, которые соответствуют длинам волн от 300 до 750 нм. Энергия фотонов излучения с длинами волн больше 750 нм уже недостаточна для осуществления элементарного фотохимического акта фотосинтеза, а энергия фотонов с длинами волн меньше 300 нм настолько велика, что вызывает разрушение белковых молекул фотосинтезирующих структур.
Зеленые растения избирательно поглощают излучения разных длин волн. Больше всего они отражают и пропускают их в зеленой области спектра. Пропущенная и отраженная части падающей на растения энергии излучения являются потерями и потенциально не могут быть использованы на фотосинтез. На каждый элементарный фотохимический акт используется только часть энергии фотона (исключение составляют только фотоны с длиной волны 680 нм, которой соответствует максимальная спектральная эффективность фотосинтеза). Неиспользованная часть фотона деградирует в теплоту и принципиально не может быть использована на процесс фотосинтеза.
Определить эксергию (свободную энергию) излучения применительно к фотосинтезу растений можно путем вычитания из общей приходящей энергии излучения рассмотренные две составляющие потенциальных потерь: отраженную и пропущенную, а также избыточную энергию фотонов, неизбежно превращаемую в теплоту.
Так как эти потенциальные потери различны для излучений разных длин волн, то учитывать их необходимо на основе спектрального состава излучения, который характеризуют функцией распределения энергии солнечного излучения по спектру (ϕ(λ)с), и функции спектральной эффективности фотосинтеза (K(λ)ф). Первую функцию определяют при помощи спектрофотометра. Вторую можно рассчитывать по спектрам поглощения фотосинтетических пигментов (хлорофиллы, каротиноиды) и их концентрации в листе растений на основе закона квантовой эквивалентности или установить экспериментально. Численные ее значения приведены в отечественных отраслевых стандартах и немецких национальных нормах DIN. Эксергия солнечного излучения в отношении фотосинтеза растений (ес) определяется по зависимости
![]() (9.3)
(9.3)
где 0,95 – максимальная спектральная эффективность излучения с длиной волны 680 нм.
По выражению (9.3) методом графического интегрирования можно рассчитать величины ес за одну секунду (мощность эксергии). Умножив ее на время облучения можно получить значение ес за этот промежуток времени. При разработке и исследовании объектов биотехнологий часто используют облучение искусственными источниками. По формуле (9.3) можно рассчитать эксергию и для излучения искусственных источников любого спектрального состава, заменив ϕ(λ)с на функцию распределения излучения по спектру для примененного источника.
Спектральный состав прямого солнечного излучения у поверхности Земли может существенно изменяться в зависимости от высоты Солнца и состояния атмосферы. Однако спектральный состав суммарного излучения (прямое + рассеянное) постоянен и не зависит от этих условий. По расчетам и измерениям эксергия суммарного солнечного излучения у поверхности Земли равна 20 % от общей суммарной энергии излучения. Это позволяет рассчитывать величину ес по простой зависимости:
ес = 0,2Q (9.4)
где Q энергия общего суммарного солнечного излучения у поверхности Земли, измеряемая существующей сетью метеостанций.
Возможности биотехнологии во многом определяются способностями человека изменить генетический аппарат организма, заставить его клетки работать в десятки, сотни и тысячи раз более эффективно для получения требуемого продукта. Но полученный биотехнологами организм, тратя энергию на бесполезный (с точки зрения этого организма) продукт, может замедлять рост, развитие. В случае микроорганизмов высокопродуктивные штаммы могут легко вытеснять из биореактора (ферментера) дикие собратья. Необходимо улучшать условия культивирования.
Высокопродуктивные сорта и гибриды, полученные методами биотехнологий, как правило, более требовательны к экологическим условиям, режимам питания. Для них в значительно большей мере, чем для традиционных сортов и гибридов, необходимо более точно проводить районирование, мелиорацию земель и совершенствовать агротехнологии. Следовательно, требуется количественно оценивать соответствие агроклиматического потенциала земли и ее плодородия потребностям (характеристикам) новых сортов и гибридов, а также требуемый для них вид и уровень мелиорации, совершенствовать сортовую и зональную агротехнологии. Названные величины количественно можно определять на эксергетической основе. Исходной величиной при этом может служить величина эксергии солнечного излучения в отношении фотосинтеза и продуктивности растений.
Для определения на количественной основе соответствия погодно-климатических условий земельного угодья эколого-физиологическим характеристикам растений данного вида, сорта, гибрида можно воспользоваться понятием эксергии агроклиматического потенциала (еап), под которой понимается то количество приходящей на поверхность Земли энергии солнечного излучения, которое при существующих погодно-климатических условиях и благоприятных других факторах может быть потенциально использовано на фотосинтез и формирование продуктивности растений. Значение этой величины можно определить по выражению
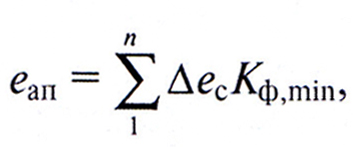 (9.5)
(9.5)
где ∆ес – эксергия солнечного излучения за промежуток времени, в течение которого значение Kф.min остается постоянным;
Kф.min – коэффициент оптимальности фактора, находящегося в относительном минимуме за этот промежуток времени;
n – число промежутков времени, за которые учитывается значение агроклиматического потенциалу.
Значение Kф можно определить по выражению
Kф= сф,Д/с ф,0, (9.6)
где сф,д – скорость фотосинтеза или формирования продуктивности данным видом (сортом, гибридом) растений при существующем значении фактора; сф,0 – та же величина, но при оптимальном значении этого же фактора.
Коэффициент оптимальности определяют для всех учитываемых погодно-климатических факторов. Сопоставляя эти значения, определяют фактор, находящийся в данный промежуток времени в относительном минимуме.
По выражению (9.6) можно определить эксергию плодородия земли (епз), если в число учитываемых факторов, наряду с погодно-климатическими, включить и факторы, характеризующие свойства плодородия почвы. При этом можно воспользоваться бонитировочной оценкой плодородия по отдельным свойствам почв, но бонитетные баллы (от 1 до 100) перевести в относительное исчисление (от 0 до 1). Под эксергией плодородия Земли понимается то количество солнечной энергии, приходящей на поверхность земли, которое при существующих погодно-климатических условиях и свойствах почвы может быть использовано данным видом (сортом, гибридом) на фотосинтез и формирование продуктивности.
По эксергии плодородия земли можно определить потенциальную (максимальную) продуктивность растений данного вида, сорта, гибрида в заданных экологических условиях. Значение этой величины можно получить умножением эксергии плодородия на коэффициент (Кд), который учитывает затраты энергии на дыхание. Численное значение этого коэффициента при отсутствии экспериментальных данных для нормальных условий выращивания можно принимать от 0,2 до 0,3 в зависимости от особенностей вида (сорта, гибрида).
Можно определить эксергию мелиоративного потенциала земельного угодия (бМп)> пользуясь выражением
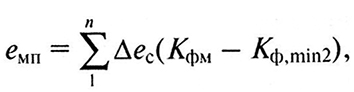 (9.7)
(9.7)
где ∆ес – значение эксергии солнечного излучения за учитываемый промежуток времени;
Кфм – значение коэффициента оптимальности мелиорируемого фактора;
Кф.min2 – коэффициент оптимальности фактора, находящегося во втором относительном минимуме после мелиорируемого.
Эта общая методика обеспечивает возможность количественного взаимно согласованного определения ключевых величин растениеводства (агроклиматический и мелиоративные потенциалы земли, ее плодородие, максимальная продуктивность растений) и выражения их в абсолютных значениях в одинаковых энергетических (точнее в эксергетических) единицах. Определение мелиоративного потенциала земли [см. формулу (9.7)] возможно в отношении любого экологического фактора (погодно-климатический, свойства почвы). При таком определении учитываются: особенности свойств конкретных видов (сортов, гибридов) растений, а также динамика влияния каждого экологического фактора как на величину мелиоративного потенциала, так и на другие рассмотренные величины.Только при определении их на единой методической основе возможно расчетное установление их при помощи ЭВМ.
Для характеристики эффективности биотехнологии или агротехнологии любой культуры целесообразно использовать три основных критерия (показателя): коэффициент полезного действия эксергетический по использованию плодородия земли (Ƞепз); показатель полезного действия эксергетический по затратам техногенной энергии (Пст); показатель эксергетический технико-экономический (Петэ). Чтобы определить эти показатели, требуется установить продуктивность (урожай), которую обеспечивает данная биотехнология (У); приведенные затраты на единицу продукции (Зп); затраты техногенной эксергии на получение единицы продукции (ет). Эксергию продукции (еп) определяют по энергии содержащихся в продукции питательных веществ (белки, жиры, углеводы). В первом приближении за эту величину можно принять энергосодержание данной продукции, которое приводится в справочниках.
Показатели эффективности биотехнологии можно определить по зависимостям:
Ƞепз = еп/епз, Пет = еn/ет, Петэ = 3п/еп. (9.8)
Используя выражения (9.1) – (9.7), можно разработать основные алгоритмы для программы расчета на ПЭВМ основных показателей эффективности альтернативных биотехнологий (агротехнологий) и выбрать наиболее результативную из них. Базовая блок-схема такой программы приведена на рис. 9.2.
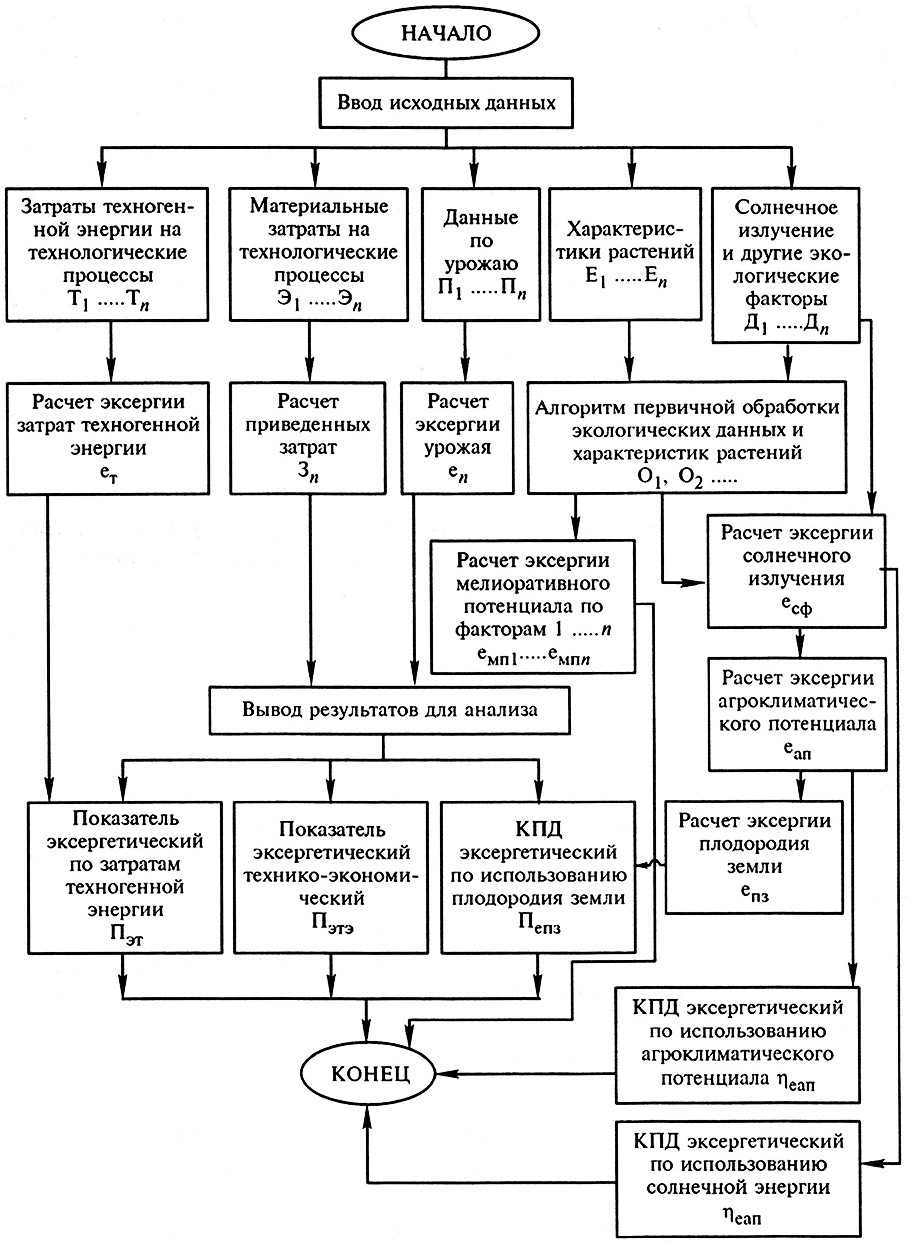
Рис. 9.2. Базовая схема компьютерной программы энергосберегающей оптимизации производства продукции растениеводства
По такой программе рассчитали показатели энергетической эффективности производства зерна яровой пшеницы (сорт Московская-35) для почвенных и метеорологических условий Московской области (данные Метеорологической обсерватории МГУ, средние значения за период 1971–1978). При урожае 40 ц/га, полученном Полевой опытной станцией Института почвоведения и фотосинтеза АН СССР, накопленная в зерне эксергия равна 5,6 МДж/м2, затрат эксергии техногенной энергии – 1,56 МДж/м2. Природная эксергия – эксергия плодородия земли – составила 162,4 МДж/м2. При этом показатель полезного действия эксергетический по затратам техногенной энергии составил 3,6, а коэффициент полезного действия по использованию эксергии плодородия земли равен 3,4 %, что свидетельствует о больших возможностях повышения урожая.
На практике во многих случаях в Московской области урожай этой культуры существенно ниже. Соответственно меньше и показатели энергетической эффективности. Например, в таких же почвенно-климатических условиях в совхозе «Заокский» Московской области, земли которого расположены рядом с Полевой станцией, в этот же период при одинаковом значении эксергии плодородия земли получен урожай зерна яровой пшеницы сорта Московская-35 18,5 ц/га. Затраты эксергии техногенной энергии на агротехнологии (которая была принята без прогнозных расчетов на соответствие экологическим условиям земли) составила 1,98 МДж/м2. В этом случае коэффициент полезного действия эксергетический по использованию эксергии плодородия земли составил примерно 1,4, а показатель полезного действия по использованию техногенной энергии–только 1,1.
Данные влияния сортовых особенностей на энергетическую эффективность разных сортов яровой пшеницы и ячменя, рекомендованных для возделывания в Центральном регионе РФ, представлены в табл. 9.1.
Таблица 9.1. Показатели оценки энергетической эффективности сортов яровой пшеницы
и ячменя, рекомендованных для возделывания в Центральном регионе РФ,
рассчитанные по результатам определения урожайности в конкурсном сортоиспытании НИИСХ ЦРНЗ
(Московская область) в 1954–2000 гг. (Э.Д. Неттевич, 2001)
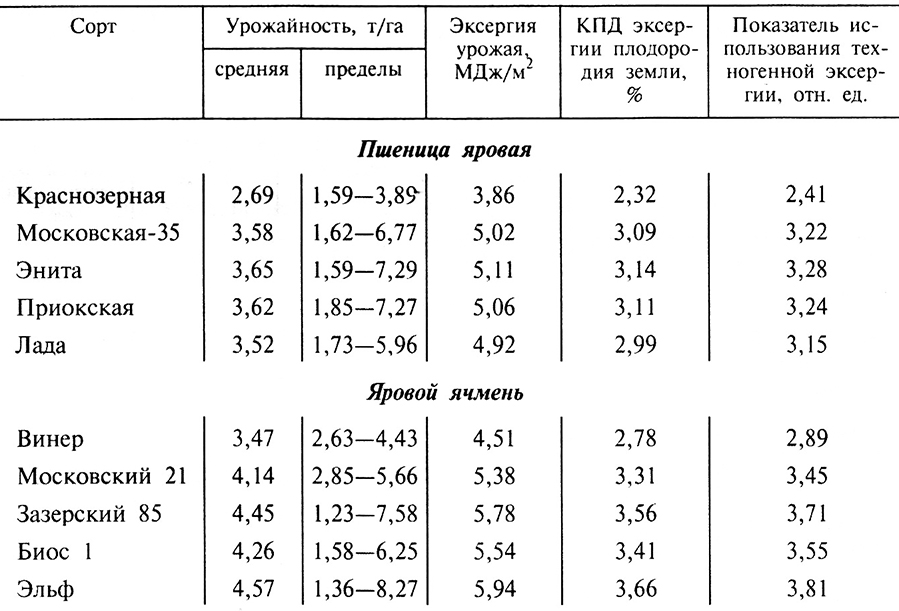
Наиболее высокие энергетические показатели по сортам пшеницы обеспечил сорт Энита: КПД использования эксергии плодородия земли – 3,14 %; показатель использования техногенной эксергии – 3,28. Самые низкие эти показатели у сорта Краснозерная (соответственно 2,32 и 2,41 %). У четырех сортов ячменя эти показатели выше, чем у пшеницы. У сорта Эльф самый высокий КПД использования эксергии плодородия земли (3,66) и показатель использования эксергии техногенной энергии (3,81), а самый низкий у сорта Винер (соответственно 3,31 и 3,45 %). Однако этот сорт, районированный еще в 1929 г., имеет самый малый разброс урожайности по годам (менее 1,7), а у самого высокоурожайного сорта Эльф этот показатель превышает 7. Для большинства сортов пшеницы и ячменя этот показатель имеет высокие значения, что свидетельствует о высокой зависимости их урожайности от погодных условий.
Влияние сортовых особенностей пшеницы на показатель использования эксергии техногенной энергии невелико (не превышает 32 %), а по КПД использования эксергии плодородия земли лишь незначительно больше – 35 %. Для сортов ячменя эти различия соответственно составили 33 и 31 %.
О влиянии сортовых особенностей на энергетическую эффективность этой культуры можно судить также по результатам испытаний новых сортов яровой твердой пшеницы, проведенных в НИИСХ ЦЧП им. В.В. Докучаева. Поданным четырехлетних полевых опытов (1988–1992) из пяти сортов наибольшую среднюю урожайность обеспечил сорт Светлана (25,2 ц/га), а самую низкую – Харьковская-46 улучшенная (20,7 ц/га). Различия в урожайности и соответственно показателях энергетической эффективности составили около 22 %, В вариантах опыта использовалась одинаковая агротехнология. При использовании сортовых агротехнологий, приспособленных к характеристикам испытываемых сортов, это различие очевидно могло быть существенно большим.
Использование рассмотренной компьютерной технологии позволяет достичь более высоких результатов при выборе альтернативных культур, например при выращивании кормов (табл. 9.2).
Таблица 9.2. Показатели энергетической эффективности культур
в кормовом пятипольном севообороте в Московской области на серых лесных почвах
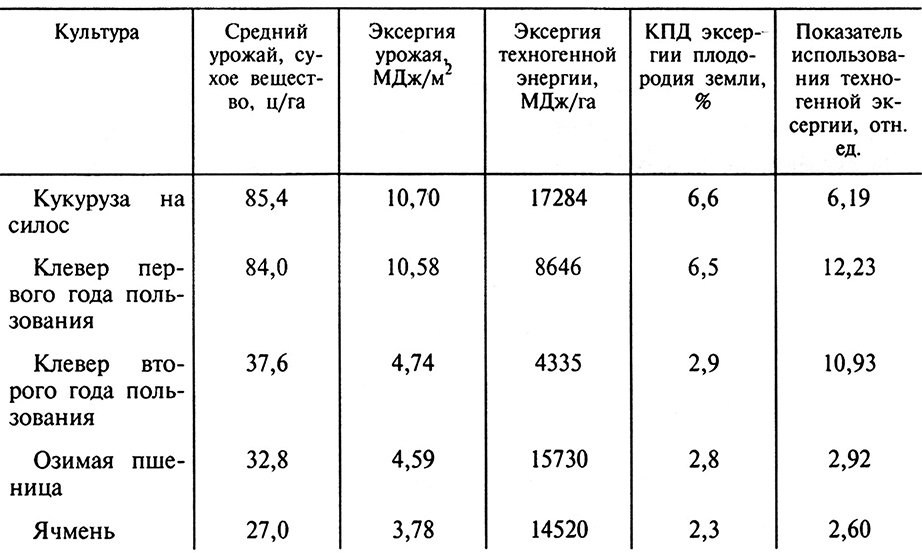
Наибольший КПД использования эксергии плодородия земли обеспечивала кукуруза (6,6 %). Близкое значение этого показателя получено и для клевера первого года пользования (6,5 %). Самое низкое значение КПД у ячменя (2,3 %). Различия почти в 2,9 раза. Самый высокий показатель использования эксергии техногенной энергии обеспечил клевер первого года пользования (12,23). Высокое значение этого показателя и у клевера второго года пользования (10,93), а самое низкое – у ячменя (2,60). Различие значений этого показателя у культур, использованных в севообороте, более чем в 2,5 раза. Зерновые культуры, как видно из этого анализа, малопригодны для использования в кормовом севообороте.
Социально-экономическое значение
биоэнергетического-эксергетического анализа
преобразований энергии
Представленный эксергетический анализ биоконверсии энергии растениями и преобразований техногенной энергии в агротехнодогиях имеет важное теоретическое и практическое значение. Это видно из следующего примера. Поданным директора института энергетической стратегии В.В. Бушуева, в 1970 г. энергоемкость отечественной внутренней валовой продукции (ВВП) была на уровне среднемирового значения. К 2005 г. этот показатель оказался примерно в 3 раза выше среднемирового и в 5,5 раза выше этого показателя в западноевропейских странах. Известно, что себестоимость сельскохозяйственной продукции, прямо пропорциональна ее энергоемкости. Ущербное состояние по энергоемкости ВВП Российской Федерации обрекает ее на роль сырьевого придатка мирового сообщества, если не принять необходимых мер.
Учеными выявлено, что высокая энергоемкость ВВП РФ обусловлена низкой эффетивностью использования первичных энергоносителей в так называемой малой энергетике – котельных и теплогенераторах (без выработки электроэнергии), используемых для обогрева бытовых, производственных помещений и в технологических процессах. В сельском хозяйстве в этих устройствах используют примерно 50 % энергоносителей потребляемой в этой отрасли.
Замена устаревших котельных и теплогенераторов на современные мини-ТЭЦ и тепловые насосы позволяет повысить КПД этих систем обогрева в 3–4 раза и приблизить энергоемкость сельскохозяйственной продукции к среднемировому значению этого показателя, повысить ее конкурентоспособность на мировом рынке.
