Как правило, кинетика роста аэробных культур микроорганизмов зависит не от одного субстрата, а от двух. Вторым субстратом, влияние которого часто преобладающее, является кислород. Общая зависимость может быть представлена в виде альтернативного уравнения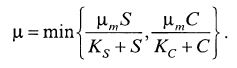 (9.43)
(9.43)
Если ферментационное оборудование недостаточно интенсивное, эта зависимость вырождается в однофакторную зависимость от концентрации растворенного кислорода С:![]() (9.44)
(9.44)
Уравнение баланса биомассы в непрерывном процессе с лимитированием кислородом такое же, как при лимитировании обычным субстратом, только вместо концентрации негазообразного субстрата S в уравнении использована концентрация растворенного кислорода С:![]() (9-45)
(9-45)
Из условия стационарности dX/dt = 0 можно найти значение установившейся концентрации растворенного кислорода: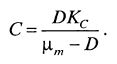 (9.46)
(9.46)
Уравнение материального баланса для субстрата дополняется поступлением кислорода за счет массопередачи в жидкую фазу: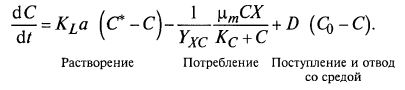 (9.47)
(9.47)
Последним членом уравнения в связи с низкой растворимостью кислорода в жидкости можно пренебречь.
Для стационарного состояния (dC/dt = 0) можно записать:![]() (9.48)
(9.48)
Отсюда![]() (9.49)
(9.49)
Концентрация негазообразного субстрата S, хотя и не лимитирует процесс, также зависит от D:
S = S0 – Х/YXS, (9.50)
или![]() (9.51)
(9.51)
Производительность хемостата определяется формулой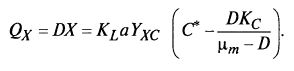 (9.52)
(9.52)
Следовательно, производительность существенно зависит от величин KLα и С*.
Критическая скорость разбавления DKP определяется по выражению![]() (9.53)
(9.53)
Полученные зависимости представлены на рис. 9.10. На рис. 9.11 показано, что в данном случае с возрастанием коэффициента разбавления продуктивность все время снижается, так как снижается скорость сорбции кислорода. Однако при некотором достаточно малом значении скорости разбавления концентрация негазообразного субстрата в соответствии с (9.50) снижается настолько, что начинает лимитировать рост биомассы. При этом зависимость QX(D) приобретает вид, характерный для хемостата. Этот участок изображен пунктирной линией.
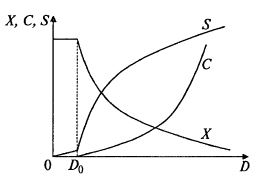
Рис. 9.10. Зависимость стационарных концентраций биомассы X,
растворенного кислорода С и нелимитирующего негазообразного субстрата S
от скорости разбавления в хемостате при лимитировании кислородом; D0 –
скорость разбавления, при которой происходит переход от лимитирования
негазообразным субстратом к лимитированию кислородом
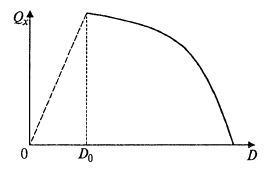
Рис. 9.11. Зависимость продуктивности по биомассе
от скорости разбавления в хемостате с лимитированием по кислороду
Таким образом, в этом случае оптимальной скоростью разбавления является как раз точка переключения лимитирования с растворенного в жидкости субстрата S на кислород.
Повышение величины KLa (т. е. массообменных возможностей аппарата) позволяет сдвинуть эту точку вправо и получить более высокую производительность процесса по биомассе.
