Объектами молекулярной биотехнологии являются самые разнообразные биологические системы: микроорганизмы, клеточные линии насекомых, растений и млекопитающих, вирусы насекомых, растений и млекопитающих, многоклеточные организмы (растения, мыши, домашние животные и т. д.) – выбор системы зависит от целей эксперимента. Характер биологической системы исключительно важен для биотехнологического процесса. Во многих случаях именно генетически модифицированная самовоспроизводящая биологическая единица – микроорганизм, вирус, растение или животное – является конечным коммерческим продуктом. Среди множеств? биологических объектов, использующихся в молекулярной биотехнологии, основными «рабочими лошадками, являются бактерии Escherichia coli, одноклеточные дрожжи Saccharomyces cerevisiae и различные клеточные линии животного происхождения. Все они играют важную роль в получении белков, кодируемых клонированными генам]
В последующих главах мы детально опишем различные высокоспециализированные биологические системы. В частности, в гл. 7 будет рассмотрена система «вирус насекомых -клетки насекомых», которая используется для продукции аутентичных белков, кодируемых клонированными генами, а в гл. 19 – генетическая модификация домашних животных (коров, овец, свиней). В настоящей главе мы дадим краткое описание наиболее значимых для молекулярной биотехнологии систем, которые также будут рассматриваться в последующих главах.
ПРОКАРИОТЫ И ЭУКАРИОТЫ
Все живые организмы делятся на две основные группы: прокариоты и эукариоты. В основе этой классификации лежат многочисленные структурные различия, на которых мы остановимся более детально в последующих главах, а здесь укажем лишь основные из них: 1) наличие или отсутствие ядра, содержащего хромосомную ДНК; 2) строение и химический состав клеточной стенки и 3) наличие или отсутствие субклеточных цитоплазматических органелл. В прокариотической клетке, например бактериальной, хромосомная ДНК находится непосредственно в цитоплазме, клетка окружена ригидной клеточной стенкой, в состав которой часто входит пептидогликан, но не хитин или целлюлоза; в клетке нет субклеточных цитоплазматических органелл. В эукариотической клетке имеется ядро, отделенное от цитоплазмы ядерной мембраной, хромосомная ДНК находится в ядре; клеточная стенка, если она есть, может сод г ржать хитин или целлюлозу, но не пептидогликан; в цитоплазме содержатся различные субклеточные органеллы (митохондрии, аппарат Гольджи, хлоропласты в клетках растений) (рис. 2.1).
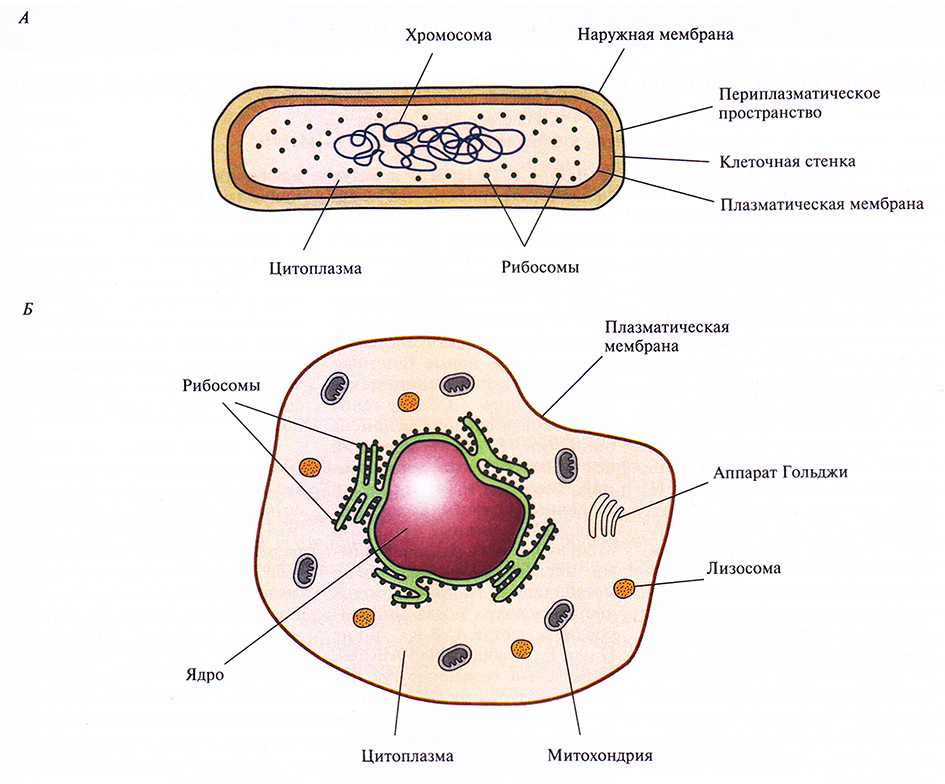
Рис. 2.1. Схематическое представление прокариотической бактериальной клетки (А) и эукариотической животной клетки (Б).
Escherichia coli
Бактерия Escherichia coli – один из наиболее хорошо изученных организмов. За последние пятьдесят лет удалось получить исчерпывающую информацию о ее генетике, молекулярной биологии, биохимии, физиологии и общей биологии. Это грамотрицательная непатогенная подвижная палочка длиной менее 1 мкм. Ее средой обитания является кишечник человека, но она также может высеваться из почвы и воды. Благодаря способности размножаться простым делением на средах, содержащих только ионы Na+, К+, Mg2+, Са2+, NH4+, С1~, НР042- и S042-, микроэлементы и источник углерода (например, глюкозу), Е. coli стала излюбленным объектом научных исследований. При культивировании E.coli на обогащенных жидких питательных средах, содержащих аминокислота, витамины, соли, микроэлементы и источник углерода, время генерации (т. е. время между образованием бактерии и ее делением) в логарифмической фазе роста при температуре 37 °С составляет примерно 22 мин.
Для каждого живого организма существует определенный температурный интервал, оптимальный для его роста и размножения. При слишком высоких температурах происходит денатурация белков и разрушение других важных клеточных компонентов, что ведет к гибели клетки. При низких температурах биологические процессы существенно замедляются или останавливаются совсем вследствие структурных изменений, которые претерпевают белковые молекулы. Исходя из температурного режима, который предпочитают те или иные микроорганизмы, их можно подразделить на термофилы (от 45 до 90 ° С и выше), мезофилы (от 10 до 47 °С) и психрофилы, или психротрофы (от –5 до 35 °С). Микроорганизмы, активно размножающиеся лишь в определенном диапазоне температур, могут быть полезным инструментом для решения различных биотехнологических задач. Например, термофилы часто служат источником генов, кодирующих термостабильные ферменты, которые применяются в промышленных или в лабораторных процессах, а генетически видоизмененные психротрофы используют для биодеградации токсичных отходов, содержащихся в почве и воде, при пониженных температурах.
Е. coli можно культивировать как в аэробных (в присутствии кислорода), так и в анаэробных (без кислорода) условиях. Однако для оптимальной продукции рекомбинантных белков Е. coli и другие микроорганизмы обычно выращивают в аэробных условиях. Если целью культивирования бактерий в лаборатории является синтез и выделение определенного белка, то культуры выращивают на сложных жидких питательных средах в колбах. Для поддержания нужной температуры и обеспечения достаточной аэрации культуральной среды колбы помещают в водяную баню или термостатируемую комнату и непрерывно встряхивают. Такой аэрации достаточно для размножения клеток, но не всегда – для синтеза белка. Рост клеточной массы и продукция белка лимитируются не содержанием в питательной среде источников углерода или азота, а содержанием растворенного кислорода: при 20 °С оно равно примерно девяти миллионным долям. Это становится особенно важно при промышленном получении рекомбинантных белков с помощью микроорганизмов. Для обеспечения условий, оптимальных для максимальной продукции белков, конструируют специальные ферментеры и создают системы аэрации.
__________________________________________________________________
Таблица 2.1. Некоторые генетически модифицированные
микроорганизмы, использующиеся в биотехнологии
Acremonium chrysogenum
Bacillus brevis
Bacillus subtilis
Bacillus thuringiensis
Corynebacterium glutamicum
Erwinia herbicola
Escherichia coli
Pseudomonas spp.
Rhizobium spp.
Streptomyces spp.
Trichoderma reesei
Xanlhomonas campestris
Zymomonas mobihs
__________________________________________________________________
Помимо E. coli, в молекулярной биотехнологии используют множество других микроорганизмов (табл. 2.1). Их можно разделить на две группы: микроорганизмы как источники специфических генов и микроорганизмы, созданные генноинженерными методами для решения определенных задач. К специфическим генам относится, например, ген, кодирующий термостабильную ДНК-полимеразу, которая используется в широко применяемой полимеразной цепной реакции (ПЦР). Этот ген был выделен из термофильных бактерий и клонирован в Е. coli. Ко второй группе микроорганизмов относятся, например, различные штаммы Corynebacterium glutamicum, которые были генетически модифицированы с целью повышения продукции промышленно важных аминокислот.
ВАЖНАЯ ВЕХА
Появление новых генотипов в смешанной культуре биохимических мутантов бактерий
J. Lederberg, E.L. Tatum Cold Spring Harbor Symp Quant. Biol. 11: 113-114, 1946.
Почти до середины XX в. среди бактериологов господствовало мнение, что в отличие от других живых организмов бактерии при неблагоприятных внешних воздействиях выживают не благодаря случайным генетическим изменениям (мутациям), а вследствие того, что именно эти воздействия в большинстве случаев запускают физиологические процессы, которые и позволяют бактериям выжить. Эта теория была опровергнута исследованиями С.Е. Лурия и М. Дельбрюка (Luria S.E., Delbruck М., Genetics 28: 491-511, 1943), которые доказали, что устойчивость Е. coli к бактериальным вирусам (бактериофагам) обусловлена именно произошедшими в них мутациями, а не реакцией бактерий на воздействие со стороны бактериофага. Эти данные нашли подтверждение в работах других авторов, изучавших последствие других неблагоприятных внешних воздействий. Исследования Лурия–Дельбрюка положили начало современной генетике микроорганизмов.
В 1946 г. Ледерберг и Татум продемонстрировали, что между членами генетически неоднородной популяции Е. coli может происходить обмен генетической информацией и что при этом, как и у двуполых организмов, в результате физического обмена между хромосомами могут возникать новые генетические комбинации (генетическая рекомбинация). Обнаружение этого замечательного феномена выдвинуло Е. coli на передний фронт генетических исследований. Ледерберг и Татум создали различные штаммы Е. coli, индуцируя те или иные мутации и отбирая клетки с разнообразными наследуемыми дефектами метаболизма. Например, одна чистая культура могла синтезировать вещества А, В и С, но не D или Е (ее обозначили ABCde). Параллельно создали другую чистую культуру, abcDE. Смешав эти культуры, получили бактерии ABCDE. Ни в одной из чистых культур тип ABCDE не возникал спонтанно. Экспериментально исключив остальные возможные причины образования комбинации ABCDE, Ледерберг и Татум пришли к выводу, что она возникает в результате физического обмена генетическим материалом между двумя хромосомами. Другими словами, у Е. coli существует некое подобие аппарата полового размножения, который, как и у других организмов, обеспечивает создание новых комбинаций генов.
Как заявил Лурия, это открытие может стать «одним из наиболее великих достижений за всю историю развития бактериологии», что впоследствии и подтвердилось. Вскоре появилась целая серия блестящих работ самого Ледерберга и других авторов, которые позволили установить, как работает генетическая система Е. coli. Этот микроорганизм широко использовался как модельная система для изучения молекулярных основ синтеза нуклеиновых кислот и белков, а также других важнейших биологических процессов.
Saccharomyces cerevisiae
Дрожжи Saccharomyces cerevisiae – это непатогенные одноклеточные микроорганизмы с диаметром клетки примерно 5 мкм, которые во многих отношениях представляют собой эукариотический аналог Е. coli. Их генетика, молекулярная биология и метаболизм детально изучены. S. cerevisiae размножаются почкованием и хорошо растут на такой же простой среде, как и Е. coli. Их способность к превращению сахара в этанол и углекислый газ издавна использовалась для изготовления алкогольных напитков и хлеба. В настоящее время ежегодно во всем мире расходуется более 1 млн. тонн S. cerevisiae. Дрожжи S. cerevisiae представляют также большой научный интерес. В частности, они являются наиболее удобной моделью для исследования других эукариот, в том числе человека, поскольку многие гены, ответственные за регуляцию клеточного деления S. cerevisiae, сходны с таковыми у человека. Это открытие способствовало идентификации и характеристике генов человека, отвечающих за развитие новообразований. Широко используемая генетическая система дрожжей (искусственная хромосома) является непременным участником всех исследований по изучению ДНК человека. В 1996 г. была определена полная нуклеотидная последовательность всего набора хромосом S. cerevisiae, что еще более повысило ценность этого микроорганизма для научных исследований. Такая работа на эукариотах была выполнена впервые.
Синтезированный бактериальной клеткой эукариотический белок часто приходится подвергать ферментативной модификации, присоединяя к белковой молекуле низкомолекулярные соединения – во многих случаях это необходимо для правильного функционирования белка. К сожалению, Е. coli и другие прокариоты не способны осуществлять эти модификации, поэтому для получения полноценных эукариотических белков используют S. cerevisiae, а также другие виды дрожжей: Kluyveromyces lactis, Saccharomyces diastaticus, Schizisaccharomyces pombe, Yarrowia lipolytica, Pichia pastoris, Hansenula polymorpha. Наиболее эффективными продуцентами полноценных рекомбинантных белков являются P. pastoris и Н. polymorpha.
КУЛЬТУРЫ ЭУКАРИОТИЧЕСКИХ КЛЕТОК
При всех различиях между типами эукариот методические подходы к культивированию клеток насекомых, растений и млекопитающих имеют много общего. Сначала берут небольшой кусочек ткани данного организма и обрабатывают его протеолитическими ферментами, расщепляющими белки межклеточного материала (при работе с растительными клетками добавляют специальные ферменты, разрушающие клеточную стенку). Высвободившиеся клетки помещают в сложную питательную среду, содержащую аминокислоты, антибиотики, витамины, соли, глюкозу и факторы роста. В этих условиях клетки делятся до тех пор, пока на стенках емкости с культурой не образуется клеточный монослой. Если после этого не перенести клетки в емкости со свежей питательной средой, то рост прекратится. Обычно удается переносить (перевивать, субкультивировать) и поддерживать до 50–100 клеточных генераций исходной (первичной) клеточной культуры, затем клетки начинают терять способность к делению и гибнут. Культивируемые клетки сохраняют некоторые свойства исходного клеточного материала, поэтому их можно использовать для изучения биохимических свойств различных тканей.
Часто некоторые клетки перевиваемых первичных клеточных культур претерпевают генетические изменения, в результате которых ускоряется их рост. Культуры клеток, которые при этом приобретают селективные преимущества, оказываются способными к неограниченному росту in vitro и называются устойчивыми клеточными линиями. Одни клеточные линии сохраняют основные биохимические свойства исходных клеток, другие нет. У большинства клеток, способных к неограниченному росту, имеются значительные хромосомные изменения, в частности отмечается увеличение числа одних хромосом и потеря других. В молекулярной биотехнологии устойчивые клеточные линии иногда используют для размножения вирусов и для выявления белков, которые кодируются клонированными последовательностями ДНК. Кроме того, они применяются для крупномасштабного производства вакцин и рекомбинантных белков.
ЗАКЛЮЧЕНИЕ
В молекулярной биотехнологии используется множество различных биологических систем – как для осуществления генетических манипуляций, так и для производства важных в коммерческом отношении продуктов. Наиболее значимыми из них являются бактерия Escherichia coli, дрожжи Saccharomyces cerevisiae и клеточные культуры насекомых, растений и млекопитающих.
