Фотосинтез является ключевым процессом жизнедеятельности и осуществляется в основном в растениях. В простейшей форме он описывается реакцией
Растения
H2O 4- CO2 -------*- Органические вещества + O2.
Солнечная энергия
Кроме углерода, водорода и кислорода в ходе светозависимых реакций, протекающих в растениях, в состав органических веществ включаются также азот и сера.
Основные процессы фотосинтеза сегодня уже хорошо известны. Они протекают в хлоропластах (рис. 2.2), которые поглощают CO2, поступающий в растение путем диффузии. Первичный процесс карбоксилирования осуществляется в строме (части хлоропласта, содержащей мало мембран) и катализируется рибулозобисфосфат-карбоксилазой (РБФК). В результате образуются две молекулы трехуглеродной кислоты (фосфоглицерата, ФГ), которые затем восстанавливаются с образованием молекул трехуглеродного сахара — триозофосфата. Это вещество в хлоропластах служит предшественником крахмала, но может поступать и в цитоплазму, где оно используется при синтезе сахарозы. Часть связанного углерода повторно поступает в вое-
становительный пентозофосфатный цикл (ВПФ-цикл), поддерживая фонд акцептора CO2, рибулозобисфосфата.
ВПФ-цикл «работает» в направлении синтеза, используя энергию гидролиза ATP и окисления NADPH. ATP и NADPH образуются в ходе так называемых световых реакций фотосинтеза, которые идут в мембранных (тилакоидных) структурах хлоропластов.
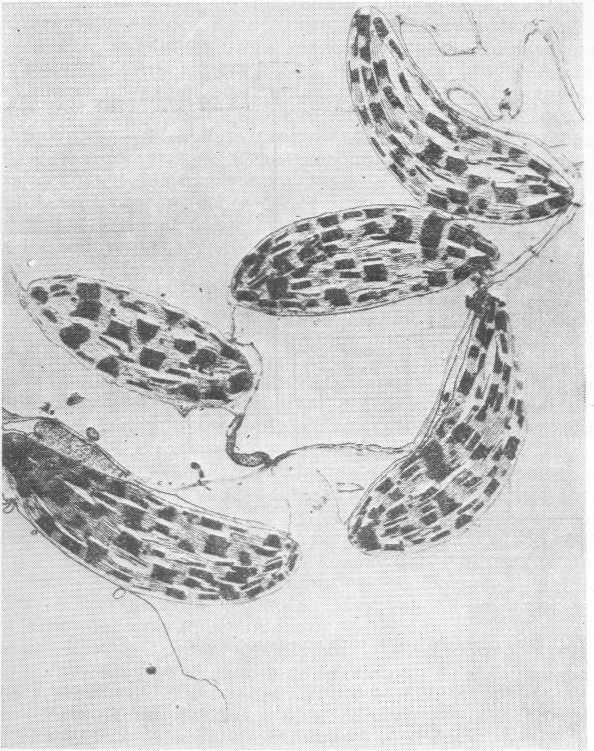
Рис. 2.2. Хлоропласты высшего растения.
Для восстановления CO2 до CH2O нужно не менее четырех электронов. Имеющиеся на сегодня данные говорят в пользу «Z-схемы» фотосинтеза, который протекает в резуль
тате кооперативного взаимодействия двух фотосистем (ФС I и ФС II, рис. 2.3), объединенных связующей цепью реакций переноса электрона. Восстановление NADP до NADPH происходит в ФС I при участии содержащего серу и железо белка ферредоксина.
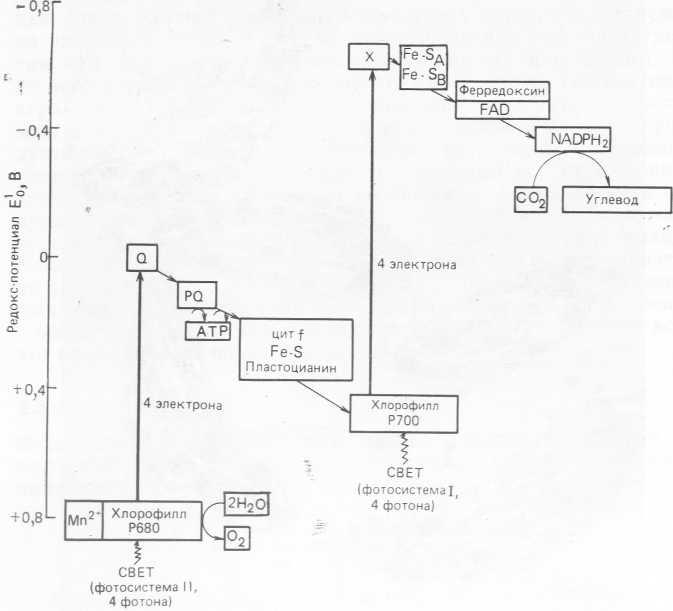
Рис. 2.3. Схема переноса электронов при фотосинтезе.
Свет улавливается своеобразными антенными хлорофилл- белковыми комплексами, и энергия его передается в реакционный центр (P700) ФС I, представляющий собой особым образом устроенный комплекс хлорофилла а и белка. Здесь происходит разделение заряда, и в результате восстанавливается ферредоксин. Электроны поступают от воды и передаются на P70O по связующей электрон-транспортной цепи от ФС II. В ходе идущей при участии ФС II реакции из воды образуется Ог.
Переносчики электронов располагаются внутри мембраны тилакоидов таким образом, что поперек нее формируется электрохимический градиент, сопряженный с переносом электронов.
Соответствующий электрохимический потенциал используется для синтеза ATP. Белок, участвующий в этом процессе, известен как сопрягающий фактор и обладает свойствами обращенной АТРазы.
Длина волны света, поглощаемого хлорофиллами и другими фотосинтетическими пигментами, определяется свойствами этих молекул; именно от этого зависит, какая часть солнечного спектра может быть использована растениями (фотосинтетически активная радиация, ФАР). Она составляет в пересчете на энергию 45—50% всего падающего на растение солнечного света. Способность улавливать свет — первый из ограничительных факторов, определяющий эффективность фотосинтеза, если исходить из количества падающего света.
При малой освещенности фотосинтез лимитируется световыми реакциями, а при более высокой интенсивности света или более низкой температуре — возможностями темновых реакций метаболизма углерода. У многих Сз-растений умеренного климата световое насыщение наблюдается при 100—150 Дж/м-2- -c^1, что соответствует 50 %-ной интенсивности солнечного света. При высоких интенсивности света и температуре влаги обычно не хватает (водный стресс). Стресс выражается в закрытии устьиц и подавлении метаболизма углерода. Таким образом, в полевых условиях взаимодействие световых и темновых реакций и их регуляция за счет диссипации (рассеивания) излишней световой энергии становятся определяющими.
В функционирующих хлоропластах каждая электрон-транспортная цепь может осуществлять акт передачи каждые 15 мс. На ярком свету молекула хлорофилла в среднем поглощает фотон один раз в 100 мс, а в условиях слабого или рассеянного освещения даже реже. Однако, поскольку каждый реакционный центр связан с несколькими сотнями светособирающих молекул пигмента, интенсивность переноса электронов увеличивается. Время с момента поглощения света до попадания возбуждения в ловушку измеряется пикосекундами. Однако перенос электрона осуществляется медленно, и лимитирующая реакция синтеза ATP протекает за время порядка 20 мс.
Как показано на рис. 2.3, в ходе фотосинтеза энергия электронов изменяется от потенциала воды (+0,8 В) до потенциала CH2O (—0,43 В). Эта разность потенциалов эквивалентна 480 кДж. Таким образом, ВПФ-цикл должен был бы работать с высокой эффективностью, чего в действительности не наблюдается.
Все дело в том, что фермент РБФК способен катализировать и окислительную реакцию между РБФ и O2, в результате которой образуются ФГ и фосфогликолат. Эта окислительная реакция конкурирует с реакцией карбоксилирования за 2ФГ, в результате чего углерод направляется по С-2-пути фотодыхания, в ходе которого образуется СОг, утрачиваемый растением.
В результате окислительных процессов и фотодыхания в Сз-растениях эффективность фотосинтеза уменьшается на 15— 50% в зависимости от вида растений и условий окружающей среды. Эти изменения обусловлены действием нескольких факторов: 1) потерей субстрата карбоксилирования; 2) потерей углерода из ВПФ-цикла; 3) потерей CO2 растением; 4) уменьшением диффузионного градиента, способствующего диффузии в растение.
Скорость фотосинтеза зависит от поступления CO2 в лист, что в свою очередь зависит от градиента концентрации CO2 и от ряда противодействующих факторов, связанных с наличием физических и биологических барьеров для диффузии и ассимиляции:
P-K (XCO2)/(ri + r2 -f- rs),
где К— коэффициент диффузии; ri соответствует разным типам противодействия. Варьирующее противодействие при диффузии r2 — это сопротивление устьиц. Сила его контролируется светом, водой и концентрацией CO2. В частности, при недостатке воды устьица смыкаются.
Противодействие гз (сопротивление мезофилла) зависит от наличия ферментных компонентов, способных к ассимиляции CO2. Сродство фермента РБФК можно охарактеризовать константой Михаэлиса (‰). Согласно большинству последних оценок, ‰ по CO2 для РБФК составляет 10—20 мкМ, что эквивалентно содержанию CO2 в атмосфере, находящемуся в равновесии с содержанием его в воде. Другими словами, если в клетке нет механизма концентрирования CO2, то этот фермент будет работать со скоростью, равной примерно половине оптимальной.
Механизмы концентрирования CO2 существуют у некоторых водорослей и ряда высших растений с C4-thπom фотосинтеза. Группа С4-растений включает несколько родов; в основном эта тропические растения. К их числу принадлежат такие коммерчески важные культуры, как сахарный тростник, кукуруза и сорго, а также тропическая «слоновья трава» (Pennisetum). В опытах с этими растениями была зарегистрирована максимальная- скорость образования сухого вещества: у них имеется1 дополнительный цикл карбоксилирования (C4-πyτb), который работает,, как насос, перекачивающий CO2 из атмосферы к месту восстановительной реассимиляции за счет ВПФ-цикла в клетках обкладки проводящих пучков. Вначале углекислый газ атмосферы ассимилируется с образованием четырехуглеродных кислот (малата и аспартата) в наружном (мезофильном) слое фотосинтезирующей ткани. Эти кислоты переносятся в слой клеток обкладки проводящих пучков, где СОг высвобождается и заново ассимилируется. В результате этого процесса фотодыхание в С4-растениях не так выражено, что увеличивает скорость фотосинтеза и продуктивность. Впрочем, хотя эти растения и ассимилируют углерод более эффективно, они потребляют дополнительную энергию для осуществления реакций С4-пути, и поэтому менее эффективно используют энергию света. С4-растения также с большим ycπexθιM используют воду и меньше страдают от повышенной температуры. Отметим, однако, что лишь немного видов С4-растений способны хорошо расти в местностях с умеренным климатом.
