Структурно-механические свойства
Качество сычужного сгустка прежде всего характеризуется механическими показателями: твердостью, упругостью, эластичностью, пластичностью, вязкостью. Твердость - это способность тела сопротивляться сжатию; упругость - способность быстро возвращаться в исходное состояние после снятия нагрузки; жесткость - сопротивляемость деформациям сдвига; эластичными называют тела, способные под действием внешних сил к большим остаточным деформациям, которые в отличие от упругих деформаций исчезают после снятия нагрузки не сразу, а в течение определенного, иногда довольно продолжительного времени; пластичность - способность к необратимым деформациям, т. е. к деформациям, которые остаются после снятия нагрузки; вязкость характеризует способность жидких и твердых тел сопротивляться перемещению одного слоя по отношению к другому под действием внешних сил; текучесть - свойство, противоположное вязкости, обладающее им тело при постоянном напряжении непрерывно деформируется; прочность определяется минимальными напряжениями, при которых структура разрушается.
Механические свойства определяются химическим составом геля, взаимосвязью компонентов геля друг с другом, количеством и прочностью связей между ними, т. е. составом и структурой геля, в связи с чем эти свойства обычно называют структурно-механическими. Количественно они характеризуются реологическими показателями. Реологические показатели характеризуют связь между напряжением (отношением внешних сил к площади поперечного вектору этих сил сечения) и величиной вызываемых этими напряжениями относительных деформаций. Так, сопротивляемость сдвигу характеризуется модулем сдвига (G), который равняется отношению напряжения сдвига (S) к деформации сдвига (у); вязкость - коэффициентом вязкости, равным отношению напряжения сдвига к скорости деформации (j). Сычужный сгусток обладает вязкоэластичными свойствами, т. е. обладает свойствами твердого и жидкого тела, поэтому для характеристики его свойств нужны несколько реологических показателей. Чаще всего для этого используют модуль сдвига и коэффициент вязкости. Существует корреляционная связь между эффективной вязкостью и модулем сдвига, поэтому по изменению этих величин можно судить об изменении прочности сгустка.
Изменение константы увеличения вязкости при сычужном свертывании молока, в зависимости от ряда факторов, показано в табл. 2.5.
Структурно-механические свойства молочных сгустков характеризуют модулем упругости G' (модуль запаса) и модулем потерь G", которые определяют с помощью динамического реометра с коаксиальным цилиндром как функцию вызываемой деформации от круговой частоты колебаний. G' - мера энергии, запасаемой и отдаваемой материалом в каждом цикле колебаний; G" - количество энергии, теряемой в процессе деформации в виде тепла. G' относится к эластичной, G"- к вязкостной части реакции геля на прикладываемое напряжение. G' можно назвать модулем эластичности. Тангенс потерь (tg<5 = G"/G') характеризует степень твердообразности материала: для упругого тела он равен нулю, для вязкой жидкости - бесконечности. Тангенс потерь - мера времени существования связей протеин-протеин в матрице геля. Чем выше тангенс потерь, тем с большей вязкостью реагирует гель на прикладываемое напряжение. По этим характеристикам можно вычислить динамическую вязкость ГГ = G"/w, время релаксации напряжений при данной частоте колебаний равно 2G'/G'-w.
Величины модулей, а следовательно, прочность сгустка зависит от количества, прочности и времени релаксации связей. Для очень маленьких и быстрых деформаций (область линейных упругих деформаций) зависимость модулей от главных факторов можно выразить уравнением:

Для гелей из дискретных частиц dF2/dx2 характеризует прочность связей, поскольку изменение энтропии для этих типов гелей незначительно. Такие факторы, как силы притяжения ван дер Ваальса, водородные связи, гидратационное отталкивание, конформационная энтропия, определенно играют роль в конформации казеиновых мицелл, но важность их пока не выявлена. Водородные связи образуются главным образом при связывании воды.
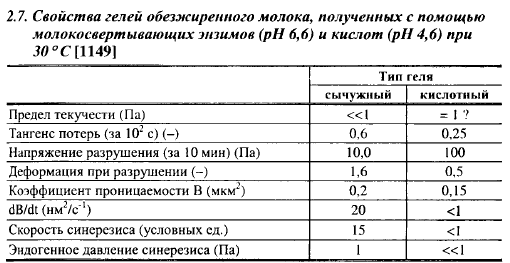
Исследования голландских ученых показали, что в сычужном сгустке относительная деформация пропорциональна прилагаемому напряжению только до тех пор, пока она не достигнет величины 0,03, что подтверждает мицеллярное строение сгустка, так как гели из макромолекул ведут себя по-иному. Предел текучести его намного меньше 1 Па, что свидетельствует о наличии остаточных деформаций при очень малых напряжениях.
В табл. 2.6 показаны основные связи, принимающие участие в формировании молочных сгустков (внутримицеллярные и межмицеллярные).
Изменения модуля G' и тангенса потерь молочных сгустков показаны на рис. 2.14, 2.15 и 2.16. Сразу после образования сгустков величина G' очень мала, что объясняется небольшим количеством связей между агрегированными параказеиновыми мицеллами в этот момент. По мере старения сгустка величина модуля растет почти линейно по отношению к логарифмической шкале времени вплоть до 6-9 ч, затем появляется плато (-180 Па), а после примерно 12 ч G' начинает снижаться. Параллельно с G' увеличивается и уменьшается G". Величина модулей возрастает вначале за счет увеличения числа связей, затем также за счет изменения характера связей: простые солевые мостики (электростатическое взаимодействие) заменяются мостиками с участием МФК. Однако незначительные изменения тангенса потерь, которые могут вызываться неоднородностью сгустка, свидетельствуют о небольшом изменении характера связей по мере старения сычужного сгустка, так как тангенс потерь характеризует природу связей, в особенности релаксацию.
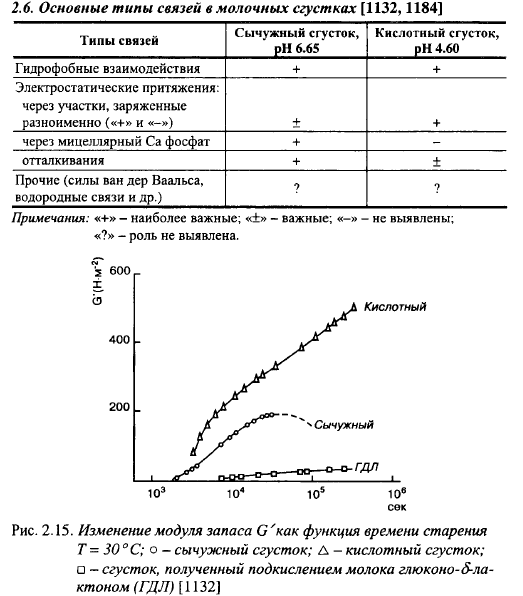
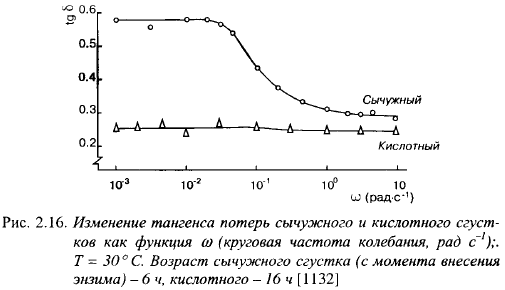
Повышение температуры сгустка после его образования снижает величины модулей G'n G", определяемых по достижению ими постоянных значений после выдержки сгустка при новой температуре (в пределах одного часа). Снижение тангенса потерь и повышение модулей свидетельствует о повышении упругости и прочности сгустка при снижении температуры. Эти изменения полностью обратимы.
Снижение прочности сгустка увеличивает его потери с сывороткой и уменьшает синерезис во время его обработки. Однако снижение прочности сгустка происходит не раньше, чем через полчаса после повышения температуры, а в начальный период модули увеличиваются. В связи с этим повышение температуры сгустка во время его обработки (II нагревание) не отражается на его технологических свойствах.
Снижение содержания мицеллярного фосфата кальция (МФК) уменьшает величины модулей, что подчеркивает важность осуществляемых с участием МФК внутримицеллярных связей в формировании прочности сгустка.
Тангенс потерь возрастал с увеличением времени деформации: он равнялся 0,28 при 0,1 с и 0,55 при 100 с. Это указывает на то, что при увеличении времени деформации сгусток становится более вязким (с увеличением времени деформации С и G" уменьшались).
Разрушение сычужного сгустка наступает при относительной деформации между 1,2 и 1,6 (табл. 2.7). Разрушающее напряжение для сычужного сгустка, характеризующее его прочность, увеличивается со снижением времени деформации: оно равно 2-104 Па при времени деформации 10'7 с, 160 Па - при 1 с, 85 Па - при 30 с и 17 Па при 10 мин.
Доза молокосвертывающего энзима
Обычно в молоко для выработки твердых сыров вносят такое количество молокосвертывающего энзима, чтобы свертывание произошло примерно за 30 мин. Необходимое для этого количество энзима в значительной степени зависит от физико-химических свойств молока. Методика его определения с помощью прибора «кружка ВНИИМС» изложена в Сборнике технологических инструкций по производству твердых сычужных сыров. Обычно на 100 кг молока вносят примерно 2,5 г препарата с активностью 100 тыс. условных ед. При необходимости ускорения этой фазы, например, при переработке сычужновялого молока, дозу энзима, по сравнению с указанной, стараются значительно не повышать. Это связано с тем, что чем выше доза сычужного порошка, тем больше химозина останется в сыре, тем интенсивнее будет протекать неспецифический протеолиз, от масштабов которого зависит качество сыра; в частности, это может способствовать появлению в сырах горечи. Так, сыр Чеддер, выработанный с применением повышенных доз сычужного порошка, на ранних стадиях созревания имел отличный вкус и эластичную консистенцию, а к концу созревания вкус его стал горьким, консистенция - мажущейся, пастообразной. Свертывание сычужновялого молока можно ускорить некоторым повышением степени зрелости молока, температуры свертывания, доз закваски и СаСП.
В отличие от скорости энзиматической фазы, скорость второй фазы не зависит от концентрации молокосвертывающего энзима. Это понятно, так как вторая фаза начинается после того, как почти весь æ-казеин уже расщеплен. Однако, если использовать низкую концентрацию молокосвертывающего энзима, то казеин выпадает в осадок без образования геля, или гель распространяется не на весь объем молока. По-видимому, это обусловлено незаконченностью первой фазы из-за низкой концентрации молокосвертывающих энзимов.
Скорость сычужного свертывания
Молоко должно быстро свертываться после внесения в него молокосвертывающих ферментов, так как именно в этом случае образуется прочный сгусток, хорошо удерживающий жир и отдающий сыворотку. Ованова обнаружила тесную корреляционную связь, близкую к функциональной (г = -0,96), между временем сычужного свертывания молока и прочностью образующегося сгустка, которую она оценивала по нарастанию предельного напряжения сдвига. Сгусток из молока с низкой сычужной свертываемостью не только медленно образуется, но и обладает малой прочностью, так как сычужновялое молоко имеет низкое содержание казеина и Са (содержание Са коррелирует с содержанием казеина), а чем ниже содержание казеина в молоке, тем меньше будет связей между параказеиновыми мицеллами в сгустке. Смешивание сычужновялого молока с молоком с нормальной свертываемостью и кислотностью в равных отношениях не полностью ликвидирует недостатки сычужновялого молока, несмотря на увеличение кислотности смеси. Следовательно, причины плохого сгустка в таком молоке лежат не просто в низкой его кислотности, а в изменении содержания и свойств казеина. Время свертывания сычужновялого молока в четыре раза выше, чем молока с нормальной сычужной свертываемостью.
Температура свертывания
Зависимость скоростей первичной и вторичной фаз сычужного свертывания молока от температуры показано на рис. 2.14. В диапазоне физиологических температур скорости увеличиваются с повышением температуры.
Скорость первичной фазы (Vеnz) равна Ккat [Е]/Кm (уравнение 3). В интервале температур от 25 до 40° С значение Ккат удваивается (Gamier, 1963), а значит, в два раза сокращается продолжительность первой фазы при прочих равных условиях. Повышение температуры с 20 до 42° С уменьшило продолжительность индукционного периода с 36 до 11 мин, или в 3,27 раза, а продолжительность стадии флокуляции - с 12,5 до 1,8 мин, или в 6,94 раза. При таком повышении температуры доля индукционного периода во времени сычужного свертывания молока (ВСС) увеличилась с 53,3 до 71,3%, доля стадии флокуляции снизилась с 18,5 до 11,7%. По Краюшкину, температурный коэффициент энзиматической фазы сычужного свертывания Qw равен примерно 2, а коагуляционной стадии - больше 10. Это означает, что температура гораздо сильнее влияет на скорость второй (неэнзиматической) фазы. В диапазоне температур от 30 до 44° С кривые продолжительности индукционного и флокуляционного периодов имеют точку перегиба при 42° С, соответствующую минимуму функций при использовании сычужного порошка и пепсинов. Наличие точки перегиба при 42° С свидетельствует о том, что влияние температуры на сычужное свертывание молока неоднозначно. Отрицательное влияние температур выше 42° С на сычужное свертывание молока вызвано постепенной инактивацией энзимов.
Скорость вторичной фазы (Vfloc) резко снижается, приближаясь к нижней температуре, при которой может происходить коагуляция молока, и при температуре ниже 15° С становится равной нулю. Раманаускас полагает, что агрегация параказеиновых мицелл идет и при температуре ниже 15° С, но с очень низкой скоростью из-за того, что эта стадия сычужного свертывания характеризуется наиболее высокой энергией активации . Если энергия активации первой фазы равна 12,9 ккал/моль, то для второй фазы она равняется 43,8 ккал/моль. Особенно быстро скорость формирования сгустка увеличивается при повышении температуры от 15 до 30° С. В опытах Раманаускаса и Урбене повышение температуры свертывания с 20 до 32° С сократило время сычужного свертывания в 4,8 раза. При температуре выше 50° С скорость агрегации приближается к теоретическому максимуму, рассчитываемому по уравнению Смолуховского, и становится независимой от температуры. Однако, по другим данным, максимальная скорость флокуляции так же, как и первой фазы реакции, наблюдалась при 42° С. Различия в этих данных, скорее всего, обусловлены разной продолжительностью наблюдений, так как при температурах выше 42° С, как указывалось выше, происходит постепенная инактивация молокосвертывающих энзимов. Минимальные продолжительности индукционного периода и стадии флокуляции при 42°С наблюдали и другие авторы.
В опытах Раманаускаса и Урбене прочность сгустка при повышении температуры свертывания с 20 до 32° С возрастала, что согласуется с возрастанием числа гидрофобных связей, играющих особо важную роль в строении сгустка (табл. 2.6). В опытах Шингаревой повышение температуры свертывания молока в интервале 30-42° С при использовании в качестве коагулянта сычужного порошка, ферментных препаратов ВНИИМС, ФП-6 и ФП-7 увеличивало прочность сгустка и скорость синерезиса, но уменьшало степень использования сухих веществ молока и жира.
Температура и ВСС молока во время выработки сыра устанавливаются с учетом обеспечения наиболее благоприятных условий для развития необходимой микрофлоры и получения требуемых органолептических показателей сыра, поэтому используют такие концентрации энзима, которые обеспечивают свертывание молока при производстве сычужных сыров при температуре 30-34° С в течение, примерно, 30 мин. Увеличение продолжительности свертывания, как указано выше, может привести к снижению прочности сгустка и скорости синерезиса. Повышение концентрации энзимов сверх установленной дозы вызывает горечь в сырах, повышение температуры свертывания интенсифицирует рост вредной и замедляет развитие необходимой микрофлоры. Однако в определенных пределах концентрацию энзимов и температуру свертывания используют для управления процессами выработки сыра. Несостоятельны попытки повысить температуры свертывания в производстве мелких сыров выше 42° С без изменения состава закваски, потому что максимальные температуры для лактококков и лейконостоков, используемых для выработки этих сыров, равны 37-40° С.
Роль кальция и фосфатов
Кальций не участвует в энзиматической фазе сычужного свертывания, однако изменение его содержания в среде может изменить физикохимические условия. Повышение количества вносимого в молоко СаС12 с 0 до 40г/100 л снизило продолжительность индукционного периода с 14,69 до 4,21 мин, т. е. в 3,5 раза, стадии флокуляции - в 3,7 раза, метастабильного равновесия - в 2,7 раза, т. е. СаС12 оказал влияние на все стадии сычужного свертывания. По Patel et al., внесение ионов Са в молоко уменьшает время сычужного свертывания и прочность сгустка через 20 мин (К2о), которые достигают минимума при 0,01 мМ Са; при высоком содержании Са (0,54мМ) значения этих величин повышаются. СаС12 может оказать влияние на сычужное свертывание молока через содержание Са2+ и изменение pH молока. При поддержании pH на постоянном уровне, добавление к молоку СаС12 в обычных дозах мало влияет на первую фазу сычужного свертывания, что хорошо видно на рис. 2.14. Следовательно, добавление к молоку СаС12 оказывает влияние на энзиматическую стадию через снижение pH молока.
Вторая фаза - флокуляция может проходить только при наличии в молоке определенного количества ионов Са. Нормальная концентрация Са2+ равна 11 мг/100 г; молоко с содержанием меньше 8 мг/100 г Са2+ является сычужновялым. При увеличении содержания Са2+ в молоке выше этого уровня время свертывания снижается пропорционально концентрации ионов.
Во время пастеризации концентрация ионов Са в молоке снижается за счет перехода части растворенного в молоке фосфата Са в нерастворимую форму и выпадения в осадок в составе так называемого «молочного камня». В молочный камень переходит из молока около 7,5% фосфатов, 16% Са и Mg. В молоке, пастеризованном при 72° С в течение 20 с, содержание Са2+ снижается по сравнению с его содержанием в исходном молоке на 15,2%. Это немного замедляет свертывание молока и скорость упрочения сгустка.
Влияние пастеризации на кинематические характеристики сычужного свертывания молока приведены в табл. 2.5 и 2.8. О скорости флокуляции в этих опытах судили по скорости увеличения вязкости. Из приведенных в табл. 2.8 данных видно, что пастеризация и хранение пастеризованного молока оказали отрицательное, а созревание, особенно пастеризованного молока с закваской, - положительное влияние на константы скорости увеличения вязкости. Положительный эффект, очевидно, связан с повышением активной кислотности молока во время созревания.
Для восстановления сычужной свертываемости пастеризованного молока в него вносят хлористый кальций. Зависимость времени сычужного свертывания молока от концентрации СаС12 Климовский выразил уравнением:
Уравнение справедливо при концентрации хлористого кальция от минимального уровня, при котором начинается коагуляция молока, до 56 г/100 л молока. На рис. 2.14 показано графически влияние количества добавляемого СаСl2 на скорость вторичной фазы сычужного свертывания молока, которая практически перестает увеличиваться при увеличении количества вносимого в молоко СаС12до 0,05%. Подобный результат получен и другими исследователями. Продолжительность свертывания пастеризованного по обычному режиму молока (72° С, 20 с) можно восстановить добавлением в него 0,056 г/л СаС12 (Humberi & Alais, 1975).
Роль ионов Са2+ в образовании сгустка состоит в том, что они осуществляют перекрестные связи мицелл друг с другом через фосфосерильные остатки казеинов. Возможно также, что положительно заряженные ионы Са2+ нейтрализуют остатки поверхностного заряда, а следовательно, способствуют снижению гидратного слоя, который частично сохраняют параказеиновые мицеллы (рис. 2.2). При внесении в молоко больше 50 мМ СаС12 поверхностный заряд мицелл казеина меняется на положительный и между ними снова возникают силы отталкивания, пропорциональные количеству внесенного в молоко СаС12. О неизбежности перехода изоточки казеина в щелочную зону при увеличении содержания ионов Са в молоке писал Глаголев еще в 1958 г.
Добавление СаС12 к молоку повышает модуль эластичности (С) (рис. 2.14) и прочность сгустка. Одна и та же эластичность сгустка была достигнута при добавлении в молоко 10, 20, 30 и 40 г/100 л СаС12 соответственно через 28, 21, 14 и 11 мин. Эластичность сгустка в конце его формирования сильно увеличивалась при добавлении к молоку от 0 до 30 г/100 л СаС12, но практически не изменилась при дальнейшем повышении его дозы до 40 г/100 л. По Раманаускасу и Урбене, эффективная вязкость сычужного сгустка повышается при добавлении к молоку до 0,08% СаС12. Дальнейшее повышение количества добавляемого к молоку СаС12 снижало эффективную вязкость и повышало тиксотропность системы.
Поскольку при пастеризации осаждается часть фосфата Са, для восстановления свертывающих свойств в пастеризованное молоко целесообразно добавлять фосфаты. Добавление фосфата Са вместо СаС12 или в качестве частичной его замены оказало существенное влияние на скорость сычужного свертывания и качество сыра. Добавление в молоко фосфата Са за 30 мин до внесения молокосвертывающего энзима снижало время сычужного свертывания; минимальным время свертывания было при внесении в молоко 0,01 М фосфата. Внесение в 100 л молока 10-20 г СаС12 и 50-70 г монофосфата натрия предотвращало излишнее снижение pH сырной массы, улучшало консистенцию сыра, снижало частоту появления горького вкуса.
Японские ученые полагают, что на структуру сыра большое влияние оказывает соотношение Са и Р, которое даже существеннее, чем влияние pH. Молоко с отношением содержания Са к содержанию Р меньше 1 свертывалось за 36,8 мин, с немного большим 1 отношением - через 34,2 мин, еще более высоким - за 27 мин.
Повышение содержания фосфата до 0,75 г/л и более немного увеличило время свертывания при незначительном изменении плотности сгустка.
Французские ученые установили, что в сычужновялом молоке отношение содержания Са к содержанию N менее 0,20, в нормальном молоке более 0,23 (Mocquot et al., 1954).
Солевое равновесие в молоке после добавления СаС12 или фосфатов наступает через определенное время. Поэтому рекомендуется их вносить в подготовленную для выработки сыра смесь с внесенным молокосвертывающим энзимом (около половины дозы) при 30-36° С, выдерживать смесь в течение 14-15 мин, а затем вносить остальную часть энзима; или вносить соли в пастеризованное молоко перед созреванием.
Коллоидный фосфат кальция (ККФ) играет важную роль в сычужном свертывании молока: снижение содержания ККФ в мицеллах уменьшает скорость агрегации мицелл казеина. При снижении содержания ККФ больше чем на 20% сычужное свертывание вообще не происходит, если не увеличить содержание ионов Са в молоке.
Влияние ионной силы, катионов и анионов
На обе фазы сычужного свертывания влияет ионная сила молока. Влияние ее на энзиматическую фазу обусловлено тем, что энзим и субстрат имеют отрицательные заряды и поэтому отталкиваются друг от друга. Изменяя ионную силу молока, можно уменьшить величину и даже знак заряда. По-видимому, по этой причине при выработке некоторых видов сыров в молоко вносят до 0,15% NaCl. При слишком высокой ионной силе молокосвертывающая активность энзимов будет снижаться, и слишком высокие дозы соли в молоке не уменьшают, а увеличивают время сычужного свертывания. Наличие максимума на кривой ВСС, как функции ионной силы, очевидно, вызвано сменой заряда казеиновых мицелл в точке перегиба.
При посолке Чеддера замена части NaCl на NaH2P04 или КН2Р04 не изменила влаго- и жироудерживающую способности, кислотность сырной массы, но заметно уменьшила степень протеолиза и выраженности сырного вкуса и аромата; консистенция сыра стала тверже, более зернистой, менее липкой и эластичной, появился самокол; цвет теста стал белесым.
Предполагают, что агрегация параказеиновых мицелл осуществляется путем электростатического взаимодействия между отрицательно заряженными участками мицеллы, освобождающимися в результате отделения от нее МП, и неидентифицированными положительно заряженными участками соседних параказеиновых мицелл. Поэтому факторы, влияющие на поверхностный заряд, влияют и на скорость сычужного свертывания молока.
С увеличением ионной силы среды скорость агрегации параказеиновых мицелл может увеличиваться и уменьшаться, что зависит от специфичности ионов [213]. Сычужное свертывание молока стимулировали Са , Mg2+, Mn2+, Cd2+, ингибировали Cu2+, Ni2+, Hg2+. Показано, что добавление в молоко Na+, К+, Li+ и Cs+ снижало концентрацию параказеиновых мицелл, необходимую для осуществления второй фазы сычужного свертывания.
Внесение в сырое молоко при 30° С FeCl3 в количестве 0,43 мМ ускоряет гидролиз æ-казеина химозином при производстве сыров, обогащенных железом.
Поликатионы могут устранить отрицательное влияние недостатка ККФ в мицелле на свертывание и даже вызвать коагуляцию казеина в отсутствие молокосвертывающих энзимов (Di Gregorio & Sisto, 1981). Добавление в молоко смеси Mn, Zn, Со и других микроэлементов сократило продолжительность сычужного свертывания молока, а также на 17% - время обработки зерна; содержание влаги в опытном Литовском сыре было ниже, чем в контрольных, на 1,7%.
Сычужное свертывание молока замедляется при внесении в него нитратов К или Na. По Раманаускасу, внесение в молоко до 0,7% поваренной соли понизило константу скорости увеличения истинной вязкости при агрегации параказеиновых частиц с 350 до 194 Па 1-с15, что свидетельствует о замедлении образования структурных связей и упрочения сгустка (табл. 2.5). В его опытах константа вязкости монотонно снижалась с увеличением количества добавляемой к молоку соли. Ионы Na обладают способностью заменять связанный с казеином Са и влиять на формирование и реологические показатели сычужного сгустка. В то же время Кайрюкштене нашла, что максимальной прочностью сычужный сгусток обладал при внесении в молоко 0,5% поваренной соли. По Patel et al., сычужное свертывание и упрочение сгустка замедляются с повышением содержания NaCl в молоке. Даже 0,5% NaCl замедляет сычужное свертывание молока.
Частичное дефосфорилирование казеина оказало небольшое влияние на первую фазу сычужного свертывания, но заметно снизило скорость второй фазы и скорость упрочения сгустка, что указывает на существенную роль фосфосерильных остатков казеинов в агрегации параказеиновых мицелл. Оноприйко для ускорения образования сгустка и синерезиса вносил в молоко после пастеризации фосфорную кислоту и фосфаты калия и натрия. К сожалению, в его опытах нет данных о влиянии этих добавок на микробиологические процессы и качество сыра.
Кислотность
Из рис. 2.14 видно, что скорости энзиматической и неэнзиматической фаз коагуляции молока снижаются при повышении pH. В опытах Раманаускаса повышение кислотности молока с 17 до 25° Т увеличило истинную константу скорости увеличения вязкости при сычужном свертывании молока с 291 до 606 Па 1-с'1’5, что свидетельствует об ускорении формирования сгустка при повышении кислотности молока (табл. 2.5).
Оптимальный pH для гидролиза <æказеина примерно равен 6,0; в интервале 5,6-6,4 скорость гидролиза изменяется незначительно. Если сычужное свертывание молока проводить при таком pH, то твердый сыр с традиционной консистенцией и вкусом не выработать из- за изменения состава и структуры мицеллы казеина и сыра (разд. 2.3). Сычужное свертывание молока проводят при pH 6,5-6,7 (17-20° Т), хотя скорость энзиматической реакции при этом намного ниже, чем при pH 6,0. В допустимом интервале кислотности молока лучше придерживаться ее верхней границы - как с точки зрения качества сыра, так и экономии молокосвертывающих энзимов. При повышении кислотности молока с 17 до 19,5° Т расход сычужного порошка снижается на 43%, свиного пепсина - на 65% при одном и том же времени свертывания; константа скорости увеличения вязкости при 20° Т в 1,34 раза выше, чем при 17° Т.
Снижение pH молока до 6,3 путем насыщения С02 уменьшает количество требуемого сычужного порошка на 75%, повышает прочность сгустка, образуемого свиным пепсином. Избыток С02 из молока можно удалить инертным газом, например азотом.
Во время выработки сыра pH снижается. Однако некоторое снижение pH после гидролиза æ-казеина для качества сыра неопасно, потому что па- раказеиновые мицеллы, в отличие от нативных, не теряют мицеллярный фосфат кальция и β-казеин и не изменяют свою структуру при снижении pH до определенного уровня. От pH зависит содержание в параказеиновых мицеллах органического Са, но, по данным Белоусова, содержание Са в сыворотке и в сгустке не меняется при снижении pH до 6,25, следовательно, в этих границах pH параказеиновые мицеллы не теряют кальция. При дальнейшем снижении pH содержание Са в сыворотке начинает быстро увеличиваться, а в сгустке и сыре - уменьшаться. Если же первую стадию сычужного свертывания молока проводить при pH 6,4-6,2 (21-25° Т), то переход Са в сыворотку резко возрастает за счет поступления в нее мицеллярного фосфата Са из нативных мицелл казеина. При внесении молокосвертывающих энзимов в молоко с кислотностью 21-25° Т сгусток уже в начале обработки будет содержать на 13-17% меньше Са, чем сгусток из молока с pH 6,5-6,6. Сыры голландской группы при нормальной скорости нарастания кислотности во время выработки содержали в среднем 0,85% Са и 0,53% Р, при слишком быстром нарастании кислотности - 0,63-0,75% Са и 0,35-0,50% Р, при оптимальном содержании Са 0,8-0,9%. Отсюда следует, что сычужное свертывание молока следует проводить при pH 6,5-6,7, а удалять сыворотку нужно тогда, когда pH сыров с низкими температурами II нагревания будет на уровне 6,25, но не ниже 6,15. Более высокая кислотность молока при проведении энзиматической фазы и в начале формования сгустка приводит к крошливой, колющейся консистенции, более низкая - к грубой, резинистой консистенции сыра.
Модули G' и G" при снижении pH после образования сгустка вначале немного возрастают (рис. 2.14), что, возможно, связано с увеличением количества положительно заряженных участков (рК гистидина ~ 6,4) и уменьшением отрицательно заряженных участков в параказеиновых мицеллах. Начиная с pH ~ 6,15, они довольно быстро снижаются с достижением минимума при pH 5,3. Таким образом, изменение модулей, в зависимости от изменения pH сгустка после его образования, согласуется с изменениями содержания в сгустке Са в опытах Белоусова. Влияние pH нельзя объяснить увеличением активности Са2+, которое не может происходить при снижении pH. По-видимому, это связано с выходом из мицелл МФК и казеинов. При pH 5,3-5,4 максимальна степень гидратации мицелл (рис. 2.2), что объясняется заполнением водой пространства, занимаемого вышедшими из мицеллы казеинами, большой пористостью мицелл. Возможно, что pH влияет на время сычужного свертывания путем воздействия на ККФ (Shaabi & Fox, 1982).
При переработке молока с высокой кислотностью увеличивается количество химозина, остающегося в сыре, что может стать причиной появления горечи в сыре. Таким образом, pH сыров, при котором проводят энзиматическую фазу, должен поддерживаться в довольно узких границах, нарушение которых ухудшает качество сыра.
Ультрафильтрация и обогащение молока СОМО
Скорость первичной фазы незначительно, а вторичной фазы - существенно нарастает при увеличении степени концентрирования молока ультрафильтрацией и при повышении дозы молокосвертывающего энзима (рис. 2.14). При постоянной дозе энзима концентрирование молока ультрафильтрацией немного замедляет сычужное свертывание и очень сильно ускоряет упрочение сгустка (модуль G' увеличивается) (Fox & Morrissey, 1972; Dalgleish, 1980; Green et al., 1983. Это можно объяснить двухфазностью свертывания: увеличение концентрации субстрата при одинаковой дозе энзима должно увеличить продолжительность энзиматической фазы; с другой стороны, увеличение концентрации казеина должно повысить число эффективных соударений параказеиновых мицелл и количество связей в сгустке. Тангенс потерь при этом не изменяется, следовательно, не меняется и характер связей в сгустке. Повышение содержания казеина с помощью добавления ретен- тата, полученного ультрафильтрацией, увеличивает максимальную плотность сгустка примерно на 15% и снижает образование сырной пыли примерно на 20% (Amram et al., 1982). В сгустке из концентрированного ультрафильтрацией молока особенно сильно увеличивается вязкость (Culioli & Sherman, 1978).
С увеличением степени концентрирования молока ультрафильтрацией ухудшаются показатели сгустка, характеризующие его способность к синерезису. Синерезис ухудшается за счет снижения проницаемости сгустка при неизменном давлении синерезиса. Однако, для сгустка из ультрафильтрованного молока, когда вся или большая часть влаги удаляется из молока во время ультрафильтрации, синерезис не играет заметной роли.
Казеиновый каркас сгустков из концентрированного УФ молока более жесткий и грубый, чем в сгустке из обычного молока, что обусловливает такие дефекты сыра, как крошливая, зернистая консистенция (Green, 1985). Он хуже удерживает жир, чем сгусток из нормального молока (Green et al. 1981), возможно, в связи с увеличением его содержания в результате ультрафильтрации. Структуру этого сгустка можно улучшить некоторым понижением температуры свертывания, а, следовательно, скорости агрегации параказеиновых мицелл. Сгусток из концентрированного молока начинает образовываться даже при 5° С.
Сычужная свертываемость концентрированного УФ молока лучше сохраняется при пастеризации его при 90-97° С по сравнению с обычным молоком. Высокотемпературная обработка такого молока повышает выход сыра, улучшает консистенцию мягких сыров (Prokopek et al., 1976).
Повышение содержания казеина в обычном молоке за счет добавления СОМО увеличивает время сычужного свертывания и плотность сгустка. В опытах Кузнецова с соавт. средняя продолжительность индукционного периода без дополнительного внесения в молоко СОМО составила 7,8 мин при средней вязкости 1,5710'3 Па с; средняя продолжительность стадии флокуляции составляла 1,9 мин. Внесение в молоко от 10 до 50 г /л СОМО удлиняло индукционный период на 0,27 мин и повышало вязкость на 0,001 Па с на каждые 10 г/л добавляемого СОМО; продолжительность стадии флокуляции повышалась на 0,26 мин с увеличением вязкости на 0,29-10'3 Па с. В этом случае при одинаковой дозе энзима снижалась его концентрация по отношению к непосредственному субстрату, что замедлило гидролиз Jf-казеина, но увеличило количество и прочность связей, участвующих в формировании сгустка.
Попытка улучшить технологические свойства сычужновялого молока путем добавления в него сухого молока низкотемпературной сушки не увенчалась успехом, так как в этом случае замедляется синерезис и образуется большое количество сырной пыли (Атгаш et al., 1962).
Состав и свойства молока
Главным показателем молока, характеризующим его сычужную свертываемость, является содержание казеина. Для получения хорошего сгустка, высокого выхода и качества сыра содержание казеина должно быть не ниже 2,5%; молоко с содержанием меньше 0,7% казеина сычужный сгусток не образует. При повышении содержания казеина в молоке выше 2,5% существует зависимость между прочностью сгустка и содержанием казеина, причем прочность растет быстрее, чем содержание казеина. Увеличение содержания казеина в молоке с 26 до 36 г/кг не оказывает влияния на время свертывания, но заметно повышает прочность сгустка. В сычужновялом молоке содержание казеина на 9-11% ниже , в нем больше у- и пара-æ-казеина и мало æ- и β-казеина. Смешивание с нормальным молоком не улучшает сычужную свертываемость сычужновялого молока. Хранение сычужновялого молока при 4° С приводит к появлению очень сильной горечи в вырабатываемом из него сыре.
С содержанием казеина коррелирует содержание в молоке Са и Р, кислотность свежего молока, т. е. показатели, играющие важную роль в сычужном свертывании молока. Все факторы, оказывающие отрицательное влияние на содержание казеина в молоке, ухудшают его сычужную свертываемость. К ним относятся заболевания коров маститами, недостаточное кормление, в частности, дефицит энергии в рационах и др. Так, например, при увеличении содержания соматических клеток в молоке (что чаще всего связано с заболеваниями коров маститами) с 500 до 1000 тыс. клеток/мл, содержание казеина снижается примерно на 15%, а время сычужного свертывания увеличивается на 28%. Еще сильнее снижается прочность сычужного сгустка.
На сычужную свертываемость молока оказывает влияние фракционный и генотипический состав казеинов. Факторы, от которых зависит содержание и состав казеинов в молоке, рассмотрены в гл. 7.
Повышение содержания жира в смеси с 0 до 4% не оказало влияния на продолжительность фаз сычужного свертывания и эластичность сгустка, немного увеличив эффективную вязкость.
Продолжительность энзиматической фазы зависит, а фазы флокуляции - не зависит от размеров мицелл казеина, хотя, по Нильсену с соавт., продолжительность фазы флокуляции более зависит от этого фактора. По их данным, если продолжительность первой фазы сычужного свертывания молока со средними размерами мицелл при 30° С принять за 1,00, то для молока с мелкими мицеллами она будет равняться 0,90, с крупными - 1,18; для второй фазы соответственно 0,72 и 1,72. Японские ученые разделили мицеллы центрифугированием на крупные и мелкие, затем подвергли сычужному свертыванию: в обоих вариантах содержание небелкового азота увеличивалось после добавления химозина в течение 20 мин, следовательно, скорость энзиматической фазы не зависела от размера мицелл. Противоречивость результатов этих опытов может объясняться тем, что наряду с размерами мицелл менялись и некоторые другие, неучтенные факторы, оказывающие большее влияние на процесс. Однако зависимость сычужной свертываемости молока от размеров мицелл вряд ли имеет существенное практическое значение для сыроделия, поскольку сортировка молока по размеру мицелл казеина на заводах невозможна. Этот вопрос может иметь значение для селекции скота в зонах сыроделия.
Тепловая обработка молока
В производстве большинства сыров молоко пастеризуют при 72-74° С в течение 15-20 с. Отрицательное влияние такой пастеризации на сычужную свертываемость молока в основном связано со снижением в нем содержания ионов Са и рассмотрено в разд. 2.5.4. Оно ликвидируется внесением в молоко СаС12, созреванием пастеризованного молока с закваской (за счет снижения pH), но не хранением при температурах меньше 7° С, во время которого pH молока обычно повышается за счет размножения психротрофов. Привлекательна идея ужесточения режима пастеризации, в том числе проведение тепловой обработки молока при температурах выше 100° С с целью повышения бактериального качества сыра, в частности, предотвращения порчи сыров маслянокислыми бактериями, споры которых не уничтожаются обычной пастеризацией.
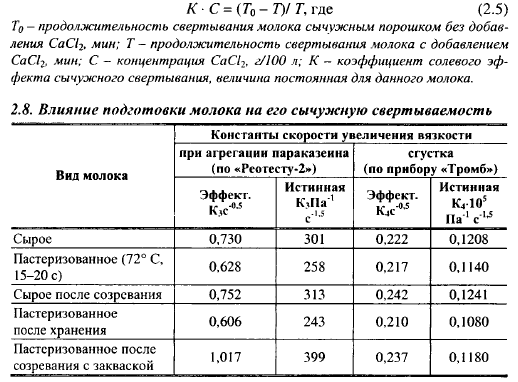
Режим тепловой обработки молока оказывает сильное влияние на первую фазу сычужного свертывания. Эффект незначителен до тех пор, пока не начнет денатурироваться β-лактоглобулин. Степень денатурации сывороточных белков зависит от температуры нагревания и времени выдержки при этой температуре (рис. 2.17). При обычно применяемой в сыроделии пастеризации молока денатурируется 7,5-9,0% сывороточных белков. Сгусток из такого молока с добавлением СаС12 имеет хорошие технологические свойства. Увеличение выдержки молока при 70° С до 15 мин приводит к получению очень слабого сгустка. При повышении температуры пастеризации с 72 до 80° С, с 15-секундной выдержкой, количество денатурированных сывороточных белков увеличивается с 7,5 до 27,5%. Сывороточные белки частично денатурируются в молоке, пастеризованном при 98° С с выдержкой в течение 0,5-1,87 мин, и в молоке, подвергнутом ультравысокой температурной обработке (140° С в течение 2-8 с); полностью - в молоке, нагреваемом до температуры 85° С и выдерживаемом при этой температуре 10-40 мин.
Жесткая температурная обработка молока ингибирует энзиматическую фазу сычужного свертывания. Ингибирование состоит в том, что в молоке, обработанном при высоких температурах, снижается скорость гидролиза æ-казеина, и часть его совсем не гидролизируется молокосвертывающими энзимами. При повышении температуры пастеризации с 70 до 95° С с выдержкой 15 с, скорость первичной фазы снизилась на 25%, дальнейшее повышение температуры при этой выдержке не оказывало на нее влияния. При такой обработке денатурировалось 25% сывороточных белков молока. Скорость первичной фазы снижалась при повышении температуры пастеризации линейно по отношению к количеству денатурированного β-лакто глобулина. Количество МП - продукта гидролиза ге-казеина - снизилось при тепловой обработке при 120° С в течение 5 мин на 10% (при температурах ниже 95° С количество отщепляемого МП не уменьшалось). При этом часть æказеина (~5%) была гидролизо- вана в результате тепловой обработки до внесения в молоко химозина.
В другом опыте обезжиренное молоко и мицеллы казеина, суспендированные в ультрафильтрате молока с добавлением или без добавления сывороточного альбумина (СА), βлактоглобулина (ЛГ) и иммуноглобулина (ИГ), нагревали при 80° С в течение 30 мин, охлаждали до 30° С и свертывали клонированным химозином. С А, ЛГ и ИГ существенно замедляли сычужное свертывание молока.
Причиной отрицательного действия жесткой тепловой обработки на энзиматическую фазу сычужного свертывания является то, что денатурированный β-лактоглобулин взаимодействует с æ-казеином через дисульфидные связи, что делает æ-казеин менее доступным для моло- косвертывющих энзимов. Однако это не единственная причина ухудшения технологических свойств сгустка, так как негативное действие увеличивается с повышением температуры даже в условиях постоянной степени денатурации сывороточных белков.
Высокотемпературная обработка молока оказывает более сильное негативное влияние на вторичную фазу сычужного свертывания, что, по-видимому, связано с осаждением на мицеллах Са3(Р04)2 и изменениями мицеллярного кальцийфосфатного комплекса.
Сычужный сгусток из молока, подвергнутого высокотемпературному воздействию, имеет низкую прочность и малопригоден для изготовления твердых сыров, так как при последующей обработке он плохо отдает сыворотку, что является одной из причин появления излишне кислого вкуса и горечи, колющейся или пастообразной консистенции в продукте. По данным Оноприйко, повышение температуры пастеризации молока (с выдержкой в течение 20 с) с 70 до 80° С увеличило продолжительность сычужного свертывания молока в 1,5 раза, уменьшило скорость синерезиса в 1,7 раза.
Часто высокотемпературную обработку молока пытаются применить для увеличения выхода и биологической ценности сыра за счет захвата денатурированного сывороточного белка сгустком. Выход сыра может быть увеличен воздействием высокой температуры, но не более, чем на 5%, и в основном за счет увеличения содержания влаги. Увеличение выхода за счет включения денатурированных сывороточных белков в сгусток без модернизации технологии нивелируется увеличением потерь казеина с сырной пылью, образующейся при обработке слабого сгустка.
В целях предотвращения денатурации β-лактоглобулина молоко для выработки сычужных сыров пастеризуют при температуре не выше 75° С или термизуют (при выработке твердых сыров с высокой температурой II нагревания) при 63-65° С в течение 15-25 с. Замена пастеризации молока термизацией при 65-70° С снижает продолжительность сычужного свертывания молока в производстве твердых сыров, в зависимости от подкласса, на 2-10%, а также повышает качество сыра: она особенно полезна зимой.
Высокотемпературную обработку молока можно применять только при использовании специальных технологических приемов, восстанавливающих свойства сгустка, прежде всего прочность и синеретическую способность, и предупреждающих появление горьких продуктов, которые в большем количестве образуются из сывороточных белков во время созревания сыра.
Нежелательные последствия высокотемпературной обработки молока можно почти полностью ликвидировать снижением pH до 5,8 с последующим повышением его перед добавлением молокосвертывающих энзимов до 6,3 (Davis & While, 1960). При снижении pH из мицеллы выходит часть мицеллярного фосфата Са, а после повышения pH происходит реформация мицелл с восстановлением нормальной структуры. Нежелательное влияние на сычужное свертывание молока нагревания до 85° С, с выдержкой при этой температуре в течение 5 мин, и последующего его нагревания до 135° С без выдержки можно уменьшить понижением pH, добавлением СаС12 и непрогретых мицелл казеина.
Созревание пастеризованного молока с 0,2-0,3% закваски, во время которого кислотность увеличивается на 1-2° Т (кислотность зрелого молока должна быть меньше 20° Т), восстанавливает его технологические свойства даже если пастеризацию проводят при температурах 75-79° С в течение 15-20 с. Хранение пастеризованною молока при температурах меньше 7° С не только не восстанавливает, но даже ухудшает его технологические свойства.
Получены положительные результаты по использованию высокотемпературной пастеризации в производстве сыров Моцарелла, Чешир.
Разработана технология сыра Волжский (твердый сыр с высоким уровнем молочнокислого брожения), включающая пастеризацию молока при 77-79° С в течение 15-25 с, созревание пастеризованного молока с добавлением закваски в течение 8-16 ч при 8-12° С, термизацию перед выработкой и применение специальной закваски, уменьшающей опасность появления горечи. Термизация молока после созревания нужна для уничтожения или дезактивации бактериофагов и вредной микрофлоры, размножившейся в молоке во время созревания. В результате применения этой технологии выход сыра увеличился ~ на 3%, содержание бактерий группы кишечной палочки и стафилококков уменьшилось в 6-10 раз по сравнению с контрольным сыром, вырабатываемым по технологии Российского сыра.
Макарян и др. показали, что в производстве рассольных сыров внесение в сырое молоко 0,03-0,05% триполифосфата натрия позволяет пастеризовать его при 85-90° С без снижения качества продукции.
Высокотемпературная обработка молока может найти широкое применение в производстве мягких сыров, в которых синерезис сгустка играет менее важную роль ввиду их высокой влажности и обработка сгустка проводится менее интенсивно, что предотвращает образование больших количеств сырной пыли. Кроме этого, значительное повышение кислотности молока - характерная черта технологии большинства мягких сыров - ликвидирует большую часть негативного влияния высокотемпературной обработки на технологические свойства сгустка. Применение ее в производстве этих сыров повышает выход сыра на 4-8%.
Хранение молока при низких температурах
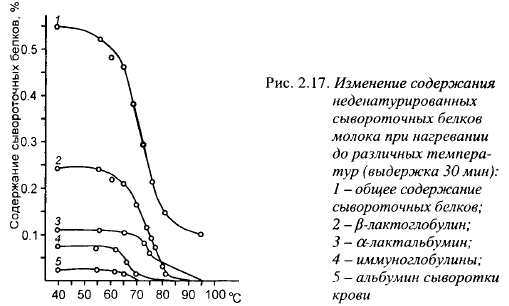
Широкое распространение получило хранение молока на фермах в течение нескольких суток при 3-4° С. Низкие температуры ингибируют рост микрофлоры в молоке, продолжительное хранение сокращает расходы на транспортировку. При низких температурах ослабляются гидрофобные связи в мицелле, часть казеинов, и прежде всего /9-казеина, выходит из мицеллы, определенное количество мицеллярного фосфата кальция также переходит в растворимое состояние, размеры мицелл уменьшаются (разд. 2.3). Кроме этого, увеличивается содержание у-казеина за счет гидролиза β-казеина и, в меньшей степени, продуктов гидролиза других казеинов плазмином (натуральная протеиназа молока) и энзимами, образуемыми размножающимися в молоке психротрофными бактериями. Эти изменения снижают выход сыра и ухудшают сычужную свертываемость молока: увеличивают время свертывания, уменьшают прочность сгустка и повышают количество сырной пыли, а также замедляют синерезис (табл. 2.9).
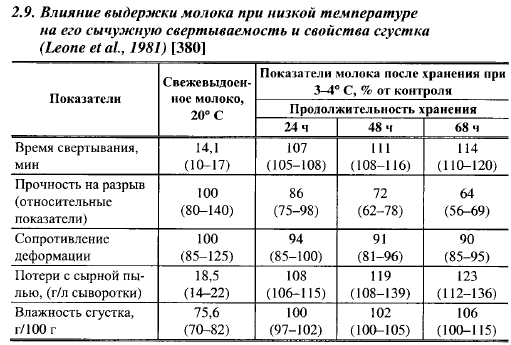
Изменения в мицелле, возникающие при низких температурах, обратимы, по крайней мере частично, при повышении температуры молока. При 20° С для частичного восстановления сычужной свертываемости требуется 18-24 ч. Скорость свертывания можно восстановить, хотя и не полностью, выдержкой молока при 45° С в течение 80 мин, при 60° С - 35 мин и пастеризацией молока при 12° С (15-20 с), однако реакция различных партий молока на эту обработку неодинакова (Qvist, 1979). Влияние различных факторов на восстановление технологических свойств молока, длительное время выдерживаемого при низких температурах, показано в табл. 2.10. Из приведенных в ней данных видно, что хорошие результаты по восстановлению сычужной свертываемости молока после холодильного хранения дает добавление в него 0,2 г/л СаС12 за час до внесения молокосвертывающего энзима. После добавления СаС12 молоко следует выдерживать при температуре 30-35° С.
Гомогенизация и другие механические воздействия на молоко
Гомогенизация молока может повлиять на энзиматическую фазу сычужного свертывания. Во время гомогенизации часть оболочек жировых шариков разрушается, а сами шарики дробятся на более мелкие. В рекомбинированные оболочки новых шариков включаются белки молока. В результате этого часть мицелл казеина связывается с компонентами оболочек жировых шариков (Oortwijn, Walstra & Mulder, 1977), что делает чувствительную связь в æ-казеине менее доступной для энзима и увеличивает время сычужного свертывания молока. Синерезис сгустков из гомогенизированного молока также замедляется (Emmons et al., 1980), что ведет к повышению влажности сыра из гомогенизированного молока. Сгусток из гомогенизированного молока обладает меньшей жесткостью, и это улучшает консистенцию вырабатываемых из него мягких сыров. В производстве твердых сыров гомогенизация может интенсифицировать липолиз и привести к появлению пороков вкуса и мажущейся консистенции.
Интенсивное механическое воздействие на молоко не оказывает прямого влияния на продолжительность энзиматической фазы. Скорость флокуляции существенно снижалась при повышении механического воздействия на молоко в ротационном вискозиметре с коаксиальными цилиндрами. Роль механических воздействий на образование сычужного сгустка, очевидно, связана с разрушением непрочных связей, участвующих в агрегации параказеиновых мицелл. Интенсивное механическое воздействие на молоко ингибирует размножение молочнокислых бактерий во время выработки сыра, что снижает скорость нарастания кислотности молока, которая влияет на его сычужную свертываемость, особенно на синерезис сгустка.
