Классификация
Лейконостоки - второй компонент заквасок для производства сыров с низкими температурами II нагревания и правильным рисунком, сыров типа Рокфор и кисломолочных сыров. Это не противоречит высказанному в предыдущем разделе положению о том, что лактококки являются необходимой и достаточной микрофлорой для производства твердых сыров с низкими температурами II нагревания, так как при наличии диацетильного лактококка лейконостоки не являются необходимой микрофлорой для выработки этих сыров. Справедливо и обратное заключение: при включении лейконостоков в состав заквасок для твердых сыров с низкими температурами II нагревания наряду со сливочным или молочным лактококками нет необходимости вводить в них диацетильный лактококк. Диацетильный лактококк и лейконостоки - два компонента газообразующей микрофлоры мезофильных заквасок для сыра, которые могут быть использованы в составе заквасок совместно и порознь.
Лейконостоки - факультативные анаэробы. Главным отличием лейконостоков от лактококков является гетероферментативность сбраживания глюкозы и лактозы с образованием D(-)-лактата, этанола и СОг в качестве основных продуктов (рис. 3.1), а также медленный рост в молоке. Возможность взаимозамены лейконостоков и Lc. lactis subsp. diacetylactis обусловлена тем, что обе группы этих микроорганизмов образуют диацетил и СО2, т.е. относятся к газообразующей микрофлоре заквасок, однако диацетильный лактококк образует эти соединения только из цитратов, а лейконостоки из лактозы и цитратов (в присутствии сбраживаемого углевода). У диацетильного лактококка способность образовывать диацетил и С02 может быть утрачена, у лейконостоков она является стабильным свойством, так как закодирована в хромосомах. Лейконостоки трансформируют в молочную кислоту около 65% сброженной глюкозы или лактозы, лактококки - до 95%.
Лейконостоки филогенетически принадлежат к группе Lactobacilli, куда, кроме них, входят два рода: Lactobacillus и Pediococcus. В 8-ом издании Берги род лейконостоков был разделен на шесть видов: Leuc. mesenteroides, Leuc. dextranicum, Leuc. cremoris. Leuc. lactis, Leuc. paramesenteroides и Leuc. oenes. Последующие генетические исследования показали столь тесное родство первых трех видов, что их решили объединить в один вид Leuc. mesenteroides с тремя подвидами: mesenteroides, dextranicum и cremoris. Генетически лейконостоки находятся в наиболее тесном родстве не с лактококками, с которыми они схожи морфологически, а с некоторыми гетерофер- ментативными молочнокислыми палочками (Lbc. confusis, Lbc. viridescens и др.), от которых они отличаются по форме клеток. Однако часто лейконостоки имеют удлиненную форму и их трудно отличить от палочек. В 9-ом издании Берджи род Leuconostoc входит в группу 17 «Грамположительные кокки» и насчитывает 9 видов. Дифференцирующие характеристики Leuc. mesenteroides и Leuc. lactis показаны в табл. 3.9 (другие виды роли в сыроделии не играют).
В производстве сыров обычно используют Leuconostoc mesenteroides subsp. cremoris (синонимы Betacoccus cremoris, Leuc. citrovorum), далее «сливочный лейконосток». От других видов и подвидов его отличает низкая метаболитическая активность, повышенная чувствительность к внешним факторам и очень сложные питательные потребности, особенно в аминокислотах, отсутствие хотя бы одной из них делает рост его невозможным. Дополнительным отличием его является расположение большого количества клеток в виде длинных цепочек, тогда как клетки других лейконостоков расположены по одиночке и в виде коротких цепочек.
Производственные штаммы лейконостоков Угличской экспериментальной биофабрики не свертывали молоко или свертывали его при посеве петлей через 36—48 ч без обесцвечивания лакмуса; предельный прирост кислотности молока при их культивировании при 30° С составлял 20-42° Т. В летнем молоке они росли лучше, чем в весеннем и зимнем.
Рост в молоке и сыре, влияние на качество сыра
Лейконостоки медленно растут в молоке и обычно его не свертывают, лакмус не восстанавливают. Только отдельные штаммы лейконостоков свертывают молоко с восстановлением лакмуса после свертывания.
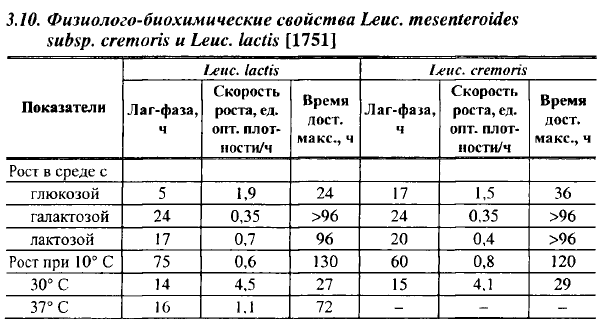
Часть клеток всех штаммов лейконостоков выживает при температуре 65° С с выдержкой в течение 30 мин. Исследования производственных штаммов лейконостоков, применяемых на Угличской экспериментальной биофабрике для производства бактериальных заквасок для сыров (ранее они фигурировали под названиями Sir. citrovoriis и St г. paracitrovorus), показали, что 16,3% их принадлежит к Leuc. lactis (далее молочный лейконосток), 63,7% - к сливочному лейконостоку и остальные имели промежуточные свойства между Leuc. lactis и Leuc. dextranicum. Следует отметить, что Str. paracitrovorus нельзя считать синонимом молочного лейконостока, выделенного из рода лейконостоков в самостоятельный вид Гарви в 1960 г. Культуры, описанные в 1928 г. Hammer и названные им Str. paracitrovorus, Гарви отнесла к педиококкам или Leuc. dextranicum. От сливочного лейконостока он отличается способностью сбраживать большее количество углеводов и более высокой устойчивостью к температуре, соли и pH, менее сложными питательными потребностями. Некоторые отличия молочного лейконостока от сливочного показаны в табл. 3.10. В средах с глюкозой и лактозой продолжительность лаг-фазы его была в 3,4 и 1,2 раза ниже, а средняя скорость роста в 1,3 и 1,75 раза выше, чем у сливочного лейконостока. В то же время в молоке при 10° С несколько быстрее развивался сливочный лейконосток.
Казалось бы, сливочный лейконосток можно считать вариантом, адаптировавшимся к молоку и молочным продуктам, поскольку в природе ему трудно конкурировать с другими представителями лейконостоков, однако и в молоке он растет очень медленно. Основной природной средой их обитания являются растительные массы, в частности силос; большой ущерб они наносят в сахарной промышленности {Leuc. те- senteroides сохраняет жизнеспособность в средах с 60% сахарозы) и виноделии. В человеческом организме они не растут.
Изучение американскими учеными роста Leuc. citrovorum в обезжиренном молоке при 22 и 30° С показало, что при 22° С максимальный уровень биомассы достигался через 22 ч, при 30° С - через 18 ч. Урожай клеток был, в зависимости от штамма, одинаковым при обеих температурах или более высоким при 22° С. Стационарная фаза продолжалась до 100 ч и более при 22° С и 26-72 ч при 30° С. Среднее время генерации при 30° С равнялось 3,2 ч, при 22° С - 3,8 ч, для некоторых штаммов оно равнялось 2,4 и 3,1 ч соответственно. Урожай биомассы лейконостоков в молоке составляет (3,3-8,4)108/мл.
Получены положительные результаты по увеличению кислотообразующей активности сливочного лейконостока путем трансформации хромосомной ДНК из молочного лактококка. Если родительский штамм сливочного лейконостока снижал pH обезжиренного молока при 30° С за 48 ч до 6,2, то генетически преобразованный - до 5,6, при этом гетероферментативный путь сбраживания лактозы сохранился.
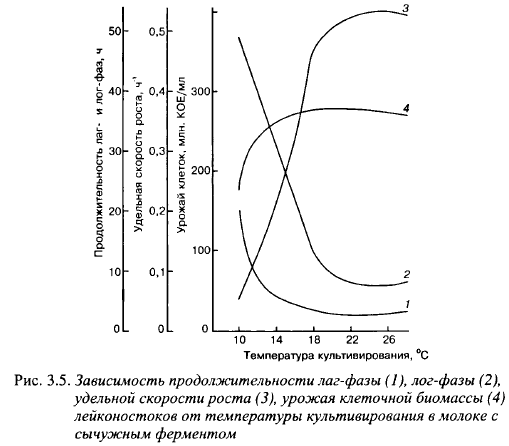
В отделе микробиологии ВНИИМС изучен рост производственных штаммов лейконостоков из коллекции Угличской биофабрики в молоке с реннетом. Параметры их роста (средние данные) приведены на рис. 3.5.
Оптимальная температура роста для изученных штаммов равнялась 22-26° С. Удельная скорость роста, продолжительность лаг- и лог- фаз роста, урожаи клеток изученных штаммов практически не менялись в интервале температуры от 20 до 30° С. Средняя продолжительность лаг-фазы равнялась 2,6 ч, удельная скорость роста - 0,51 ч_, время удвоения биомассы - 1,3 ч. Сравнивая эти данные с результатами изучения американскими учеными роста лейконостоков в молоке без добавления сычужного фермента, приведенными выше, можно сделать вывод о сильном стимулировании роста лейконостоков в молоке химозином, а следовательно, о нехватке низкомолекулярных азотистых соединений как одной из причин слабого роста лейконостоков в молоке. Рост лейконостоков в молоке во время выработки сыра также стимулирует лактококковая микрофлора закваски путем образования низкомолекулярных азотистых соединений и, возможно, других факторов роста. В то же время в жидких незабуференных средах лактококки ингибируют рост лейконостоков за счет быстрого снижения pH.
Несмотря на стимуляцию сычужным энзимом и лактококками, лейконостоки делают за время выработки сыров с низкими температурами II нагревания (от момента заквашивания до конца прессования), в среднем только 0,6 генерации при 3 генерациях лактококков. Это обусловлено слишком высокими для их роста температурами, применяемыми при выработке сыров, и длительной лаг-фазой, зависимость числа генераций от которой выражается уравнением:
![]()
У производственных штаммов лейконостоков продолжительность лаг-фазы варьировала в пределах от 1 до 3,7 ч. Штаммы с минимальной продолжительностью лаг-фазы развития в молоке успевают совершить в течение выработки сыра 1,9 генерации, штаммы с максимальной продолжительностью лаг-фазы во время выработки сыра не размножаются. Следовательно, для того чтобы оказать влияние на качество сыра, они должны размножаться во время созревания сыра.
Максимальная температура для роста сливочного лейконостока ниже 37° С, некоторые штаммы молочного лейконостока растут при 38° С.
Изучено развитие 9 штаммов Str. paracitrovorus в пастеризованном (74-75° С) молоке с сычужным порошком при температурном режиме, имитирующем режим выработки сыров с низкими температурами II нагревания (табл. 3.6) . В этом опыте в первые 2 ч инкубации при температуре 30-32° С, соответствующей температуре свертывания молока молокосвертывающими энзимами, из 9 штаммов лейконостоков только один повысил кислотность молока на 1° Т. На втором этапе культуры нагревали до 40-41° С и выдерживали при этой температуре 2 ч, что имитировало II нагревание во время выработки сыра. В конце этого этапа уже в двух культурах лейконостоков кислотность на 2° Т превышала исходную. На третьем этапе культуры выдерживались при комнатной температуре в течение 5 ч, что соответствует условиям самопрессования и прессования сыров. Температура среды к концу этапа снизилась до 25° С. Во всех культурах лейконостоков кислотность молока на этом этапе повышалась, и к концу этапа она на 1-10° Т превышала исходную. Следовательно, лейконостоки не погибают и не теряют активность при температурах II нагревания, характерных для производства мелких сычужных сыров. Устойчивость к температурам II нагревания в молоке с реннетом у лейконостоков выше, чем у лактококков, и поэтому при их повышении они могут стать преобладающими среди газообразующей микрофлоры сыров, вырабатываемых на заквасках с диацетильным лактококком и лейконостоками.
На третьем этапе культуры охлаждались до температуры 8-10° С, которая часто применяется в созревании сыров с низкими температурами II нагревания, и выдерживались при этой температуре 7 сут. Кислотность молока за этот период всеми штаммами была повышена, в т. ч. 33% на 10-40° Т. На основании этого опыта можно с высокой степенью достоверности предполагать способность лейконостоков размножаться в сыре при 8-10° С, хотя интенсивность их роста при этих температурах в значительной степени зависит от штамма.
Снижение температуры до 14° С - верхней границы созревания отечественных сыров с низкими температурами II нагревания - уменьшает удельную скорость роста лейконостоков по сравнению со скоростью роста при оптимальной температуре в 2,2-7,3 раза, при 10° С она снижается в 10 раз. Минимальная температура для роста лейконостоков, по данным других авторов, зависит от штамма и при прочих оптимальных условиях равняется 4-10° С.
Лейконостоки в сырах с высокими температурами II нагревания роли не играют, потому что до II нагревания они не размножаются, а во время II нагревания большая часть их клеток погибает.
Во время созревания сыра лейконостоки могут размножаться, пока есть лактоза. После прессования в сырах с низкой температурой II нагревания остается более 1 % лактозы, что вполне достаточно для накопления лейконостоками количества газа требуемого для формирования характерного для этих сыров рисунка.
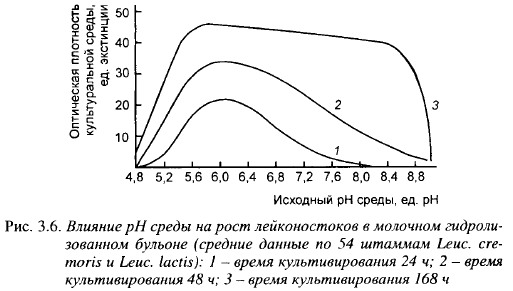
На рост лейконостоков может оказать сильное влияние активная кислотность среды. Влияние исходного pH среды на накопление биомассы лейконостоков показано на рис. 3.6. Лейконостоки активно размножались в широком диапазоне pH - от 4,8 до ~ 9,0. Наиболее активно накопление их биомассы шло при pH 5,6-6,5, что выше pH твердых сыров во время созревания. Выход биомассы при исходном pH 4,8 составлял 5-10% от ее выхода при pH 5,5. При активной кислотности, характерной для созревания мелких сычужных сыров (pH 5,2-5,4), накопление лейконостоками биомассы шло с достаточно высокой скоростью.
Исследовано образование С02 диацетильным лактококком и лейконостоками. При росте в молоке экспоненциальная фаза развития лактококков заканчивается при увеличении содержания молочной кислоты на 0,5% (pH 4,5-4,85), рост прекращается при увеличении содержания кислоты на 0,6-0,7%, что происходит примерно через 11ч инкубации при 30° С и 2 %-ной посевной дозе. Утилизация цитратов диацетильным лактококком начинается с середины экспоненциальной фазы после увеличения содержания молочной кислоты более чем на 0,2% (pH меньше 5,5). Позднее начало утилизации цитратов обусловлено тем, что цитрат пермеаза, осуществляющая перенос цитратов через клеточную мембрану, имеет низкий оптимальный pH.
В опытах А. Гудкова с соавт. в полусинтетической среде при оптимальных условиях скорость образования С02 диацетильным лактококком достигала максимума через 6 ч инкубации. Результаты этих исследований свидетельствуют о том, что диацетильный лактококк может начать образовывать газ при производстве сыров с низкими температурами II нагревания в конце выработки (прессования), а следовательно, основное образование им газа в сыре происходит во время по- солки и на начальном этапе созревания.
Лейконостоки не размножаются во время выработки и могут образовывать газ во время созревания, когда температура и другие условия в сыре далеки от оптимальных, но не подавляют их рост.
На образование газа лактококками и лейконостоками в условиях, имитирующих условия в мелких сычужных сырах во время посолки и начале созревания, сильное влияние оказывает содержание цитратов. Снижение содержания цитратов в среде на 0,08% (с 0,17 до 0,09%) уменьшило количество образованного обоими видами газа в зависимости от штамма на 66-74%. Изменение содержания в среде лактозы с 1 до 0,1% не оказало влияния на образование газа диацетильным лактококком, но уменьшило его количество в культурах лейконостоков. Изменения pH в характерном для этого периода интервале от 5,7 до 5,2 не оказало влияния на газообразующую активность лактококка, уменьшив ее у лейконостоков.
Очень большое негативное влияние на образование газа лактококками и лейконостоками оказало снижение температуры с 14 до 10° С. Скорость образования С02 диацетильным лактококком, в зависимости от штамма, снижалась при понижении на 2° С температуры в этом интервале на 31-44%, при снижении содержания цитратов на 0,08% в диапазоне от 0,17 до 0,01% она снижалась на 77-80%. Таким образом, во время посолки и в начале созревания лимитирующими образование газа диацетильным лактококком факторами являются содержание цитратов и температура. Эти факторы вкупе с содержанием лактозы лимитируют также газообразование лейконостоками, но ингибирующий эффект их в отношении этих микроорганизмов менее выражен. При снижении в среде концентрации цитратов на 0,08%, температуры на 2° С, содержания лактозы на 0,45% (в диапазоне от 1 до 0,1%) скорость газообразования лейконостоками уменьшается соответственно на 54, 20 и 13%. Скорость образования газа диацетильным лактококком в условиях, характерных для начала созревания сыра, в 1,45 раза выше, чем у лейконостоков, что, очевидно, обусловлено их более быстрым ростом. Лактококки перестают образовывать газ в сыре после утилизации цитратов, лейконостоки - после утилизации цитратов и сбраживания лактозы. Общее количество образуемого лейконостоками С02 в сыре может значительно превышать его количество, образуемое диацетильным лактококком.
Образование рисунка различными видами газообразующей молочнокислой микрофлоры проверено непосредственно в сыре. Результаты этого опыта представлены в табл. 3.11. В сырах, выработанных с диацетильным лактококком, рисунок был более мелким, но более частым, чем в сырах, выработанных с лейконостоками; в сырах с заквасками, включающими оба вида газообразователей, увеличились и число, и диаметр глазков. Во всех сырах рисунок соответствовал предъявляемым к нему требованиям и получил высшую оценку. Таким образом, нормальный рисунок в сырах с низкими температурами II нагревания можно получить с заквасками, включающими из газообразующих молочнокислых бактерий диаце- тильный лактококк (D-закваски), лейконосток (В-, или Л-закваски) и оба эти вида газообразующих бактерий (BD-, или ДЛ-закваски).

Выработки сыров в предыдущем опыте проводились в экспериментальном цехе ВНИИМС с точным соблюдением технологических и гигиенических требований. В производственных условиях могут происходить отклонения от нормального хода процесса. Так, содержание цитратов в молоке изменяется по сезонам года. Максимальным оно бывает в июле ( - 2 г/л), минимальным - в октябре, ноябре и апреле (1,33-1,53 г/л). В периоды низкого содержания цитратов в молоке часто рисунок сыров бывает редким, что можно объяснить недостатком субстрата для образования газа диацетильным лактококком, обычно представляющим газообразующую микрофлору в отечественных заквасках.
Низкое содержание цитратов в молоке может быть следствием их утилизации микрофлорой молока до переработки на сыр, так как многие микроорганизмы ферментируют цитраты. Это может быть причиной отсутствия рисунка в сырах, вырабатываемых на D-заквасках из молока с высокой бактериальной обсемененностью, в частности с повышенной кислотностью. В сырах, вырабатываемых на В- или BD-заквасках, сезонные изменения в содержании цитратов в молоке не оказывают влияния на рисунок.
Рост лейконостоков зависит от содержания в среде марганца. Дефицит Мn в среде снижает выход биомассы сливочного лейконостока на 40,7%, оказывая незначительное влияние на развитие лактококков, за исключением штаммов диацетильного лактококка с низкой кислотообразующей активностью в молоке (снижение выхода биомассы на 24,1%). Содержание Мn в молоке также зависит от сезона: в весеннем молоке среднее содержание Мn равно 23,9 мкг/кг (73% от его среднего содержания в осеннем молоке, что значительно ниже оптимального уровня для сбраживания цитратов лейконостоками (50 мкг/кг). Это приводит к менее интенсивному развитию лейконостоков и ухудшению рисунка в сырах, выработанных из весеннего молока на В-заквасках. BD-закваски весной могут превратиться в D-закваски, осенью - в В-закваски.
Рост лейконостоков в сырах прекращается после сбраживания лактозы. Чем больше остается лактозы в сырах после прессования, тем дольше продолжается развитие лейконостоков. При нормальной скорости сбраживания лактозы лейконостоки за время выработки и созревания сыра дают до 6 генераций. При избытке лактозы в сырах, например, в результате действия бактериофага на лактококки, лейконостоки могут вызвать раннее вспучивание сыра. Оно отличается от вспучивания, вызываемого бактериями группы кишечной палочки, тем, что вспученные сыры сохраняют вполне удовлетворительный вкус и аромат. Диацетильный стрептококк вспучивания твердых сыров не вызывает, так как содержание цитратов в сырной массе не бывает слишком высоким. Однако, при выработке кисломолочных сыров и творога высокое содержание в закваске диацетильного лактококка может привести к всплыванию сгустка вскоре после образования.
Бактериофаги лейконостоков менее распространены на сыродельных заводах, чем бактериофаги диацетильного лактококка (табл. 3.6). Это понятно, так как лейконостоки начинают размножаться после свертывания молока, когда условия для репродукции бактериофагов неблагоприятны. В связи с этим, В- и BD-закваски более стабильно обеспечивают формирование рисунка в сыре, чем D-закваски.
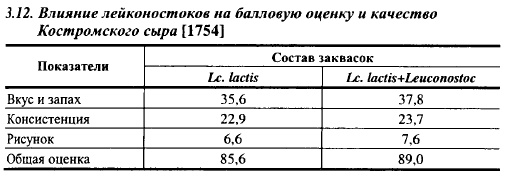
Лейконостоки оказывают сильное влияние не только на рисунок, но и на другие органолептические показатели сыров. Влияние их на качество Костромского сыра показано в табл. 3.12. Сыры, выработанные на лактококковых заквасках без газообразующих микроорганизмов, получили невысокую оценку. Введение в закваску лейконостоков повысило оценку за вкус и запах на 2,2 балла, за консистенцию - на 0,8 балла, за рисунок - на 1 балл. Сыры с лейконостоками имели более высокое содержание летучих жирных кислот (в 2,1 раза), растворимых белков (на 16%), меньше пептидов (на 38,1%), более высокий pH (5,28 по сравнению с 5,19), чем контрольные сыры. Повышенное содержание летучих жирных кислот усилило выраженность сырного вкуса, более низкое содержание пептидов могло быть причиной того, что горечь была обнаружена только в 34% опытных сыров, а в контрольных - в 75%, слишком низкий pH контрольных сыров мог быть причиной более крошливой, менее связной их консистенции. В табл. 3.7. показано, что штаммы лейконостоков не дают горечи, что обусловлено особенностями их протеолитических систем (низким уровнем образования в молоке пептидов и способностью разрушать горькие пептиды, образуемые другими компонентами заквасок). По характеру протеолиза в молоке они близки к диацетильному лактококку с низкой скоростью кислотообразования. Более высокое содержание летучих кислот и более высокий pH в сырах с лейконостоками является следствием сбраживания ими части лактозы гетероферментативным путем.
Сыры с заквасками, содержащими только молочный лактококк, не имели рисунка или имели отдельные рваные глазки, по-видимому, образованные бактериями группы кишечных палочек, которые, наряду с С02, образуют водород, плохо растворимый в сырной массе.
В других опытах средняя балловая оценка Голландского круглого и Костромского сыров, выработанных с заквасками без газообразующих бактерий, D-заквасками, В-заквасками и BD-заквасками, равнялась соответственно 85,6; 88,00; 89,3 и 90,1 балла.
Таким образом, лейконостоки в производстве мелких сычужных сыров с правильным рисунком играют важную роль. Однако слишком интенсивное развитие лейконостоков в сыре может ухудшить его качество не только за счет образования переразвитого рисунка или вспучивания головок. Продукты гетероферментативного сбраживания лактозы, в частности этиловый спирт, могут образовывать эфиры с продуктами липолиза, обусловленного развитием в сырах психротрофов, вызывающие различные пороки вкуса и запаха. Наивысшую оценку получали сыры, в которых содержалось в период максимума (2-4)-108 клеток/г лейконостоков, что обеспечивается внесением с закваской (1-3)-105 клеток на 1 мл молока (с учетом 10-кратного увеличения концентрации клеток в сгустке в результате механического захвата их коагулирующим белком). Следует, однако, учесть, что влияние лейконостоков на качество сыра в большой степени зависит от индивидуальных свойств штаммов.
Многие штаммы лейконостоков обладают выраженной специфической антибиотической активностью, в частности, по отношению к психротрофам при температуре 7° С.
По мнению Collins, диацетильный лактококк, с точки зрения образования вкуса и аромата сыра, непредсказуем и неустойчив, способен вызвать в сырах порочный «йогуртный» или «зеленый» привкус за счет образования ацетальдегида. В присутствии лейконостоков этот привкус не образуется, потому что лейконостоки трансформируют ацетальдегид в этанол.
Проведенный анализ не дает однозначного ответа на вопрос, какой вид мезофильных газообразующих молочнокислых бактерий лучше использовать для производства твердых сыров с низкими температурами II нагревания. Несомненные преимущества лейконостоков - устойчивость к бактериофагам, способность снижать опасность появления горечи и излишней кислотности в сырах, образовывать рисунок независимо от содержания цитратов в молоке. Среди лейконостоков часто обнаруживаются штаммы, обладающие специфической (обусловленной не только действием молочной кислоты) антагонистической активностью по отношению к посторонней микрофлоре. Недостатками лейконостоков являются низкая скорость роста и кислотообразова- ния в молоке и способность вызывать пороки сыров при передозировке.
Несомненным достоинством диацетильного лактококка является более высокая скорость роста и кислотообразования в молоке, хотя из зарубежных заквасок чаще выделяются штаммы Lc. diacetylactis с низкой скоростью роста. Эти штаммы в смешанной культуре со сливочным или молочным лактококком и в присутствии молокосвертывающих энзимов достаточно быстро размножаются, но, в отличие от активных по кислотообразованию штаммов, реже дают горечь.
В мировой практике применяют закваски всех трех типов: BD-, D- и В-закваски. По качеству сыра Гауда, вырабатываемого на этих заквасках, они располагались в следующем порядке: BD > D > В, однако различия были несущественны. При выборе типа закваски нужно учитывать изменения качества молока по сезонам года и гигиенические условия производства, в частности фаговую ситуацию. Весной лучше использовать D- или BD-закваски, осенью и при загрязнении закваски бактериофагами - В- или BD-закваски. В BD-заквасках очень важно соотношение культур: диацетильного лактококка в них должно быть не более 15%, лейконостоков - около 5.
Лейконостоки должны обязательно входить в закваски для производства сыра типа Рокфор для обеспечения пористости сырной массы, необходимой для развития в ней плесневых грибов. Обогащение молока лейконостоками путем непосредственного внесения в молоко их концентрата уменьшило число сыров с замкнутой структурой, при наличии которой плесень не может размножиться до нужных для формирования требуемых органолептических показателей сыра количеств, с 22,6 до 1,7%. Пустоты в Рокфоре могут образовать дрожжи, но они дают побочные привкусы в сыре.
