Это разнородная группа бактерий вибриоидной или спиральной формы с числом витков от одного неполного до множества. Для многих видов характерно образование внутриклеточных гранул поли-β-гидроксибутирата. Газовые вакуоли, стебельки, простеки и эндоспоры отсутствуют. У некоторых видов часть клеток в старых культурах превращается в так называемые кокковидные тела с тонкой клеточной стенкой. Для некоторых видов (рода Azospirillum) характерно образование крупных окруженных капсулой цист, устойчивых к высыханию. Клетки молодых культур окрашиваются по Граму отрицательно, у одного рода (Azospirillum) в старых культурах могут присутствовать формы, окрашивающиеся по Граму положительно. Выращенные в жидких средах клетки подвижны за счет полярно расположенных жгутиков. Они перемещаются по прямой, совершая при этом характерное винтовое движение. Аэробы или микроаэрофилы, хотя некоторые виды могут также расти в анаэробных условиях, используя в качестве конечного акцептора электронов нитрат, фумарат или триметиламиноксид. Многие виды не способны катаболизировать углеводы. Представители некоторых видов (рода Azospirillum) могут подкислять в анаэробных условиях среды с некоторыми сахарами, но растут при этом очень слабо, и для них явно предпочтительны аэробные условия. Характерный признак – наличие оксидазной активности. Индол не образуют. Хемоорганотрофы, однако клетки некоторых видов могут расти и как факультативные автотрофы, используя в качестве донора электронов газообразный Н2. Фототрофных организмов в группе нет. Некоторые виды способны фиксировать N, но только в микроаэробных условиях. Один вид (Aquaspirillum magnelotacticum) обнаруживает таксис к магнитному полю. Встречаются в почве, пресной воде, сильно соленых или морских средах, в корнях растений либо репродуктивных органах, кишечном тракте и ротовой полости человека и животных. Бактерии трех родов – Bdellovibrio, Micavibrio и Vampirvvibrio – хищники по отношению к другим микроорганизмам.
Дифференциация родов Группы 2: См. табл. 2.1.
Таблица 2.1. Дифференцирующие признаки подвижных, аэробных или микрааэрофильных,
грамотрицательных, изогнутым бактерий1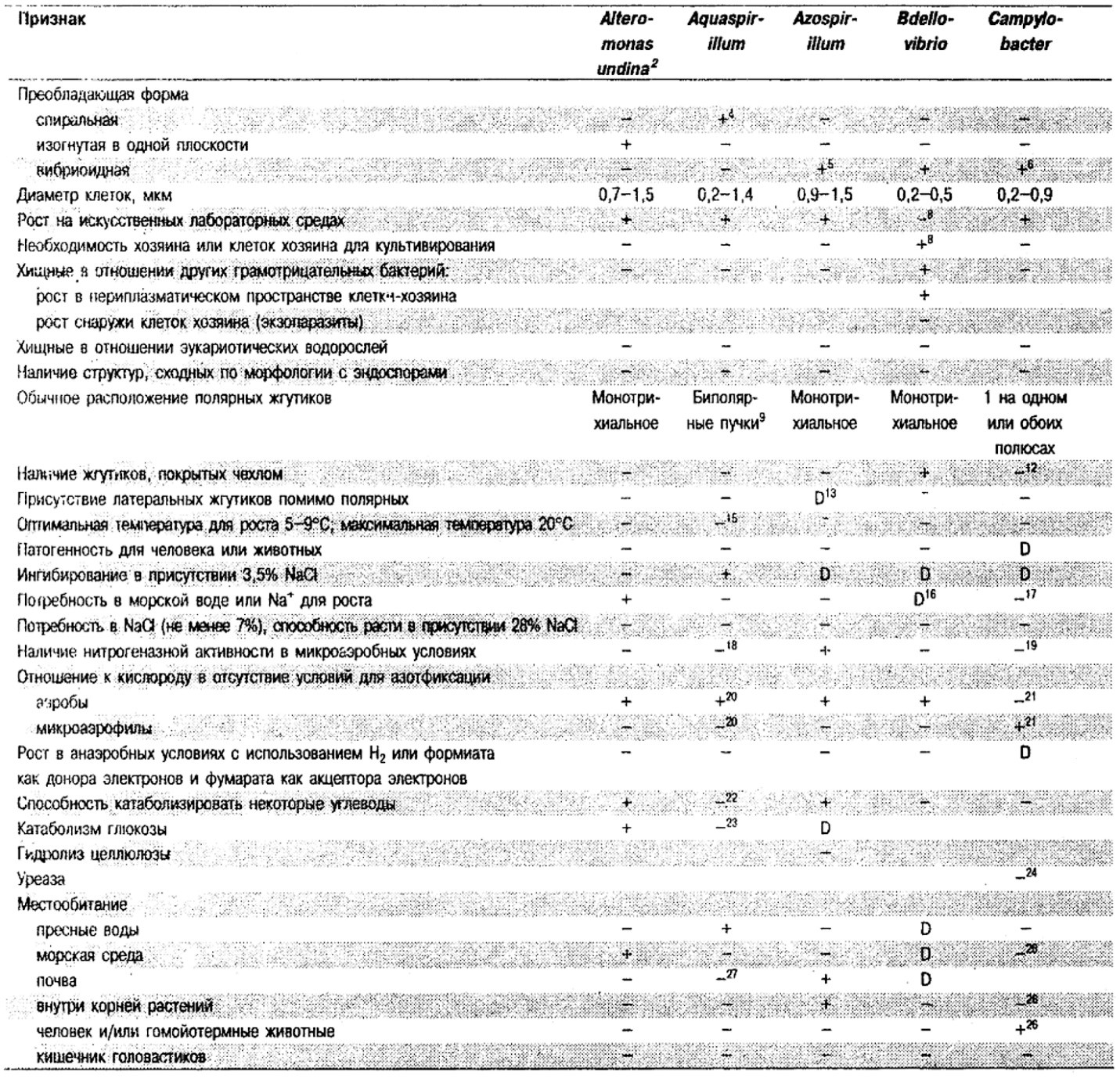
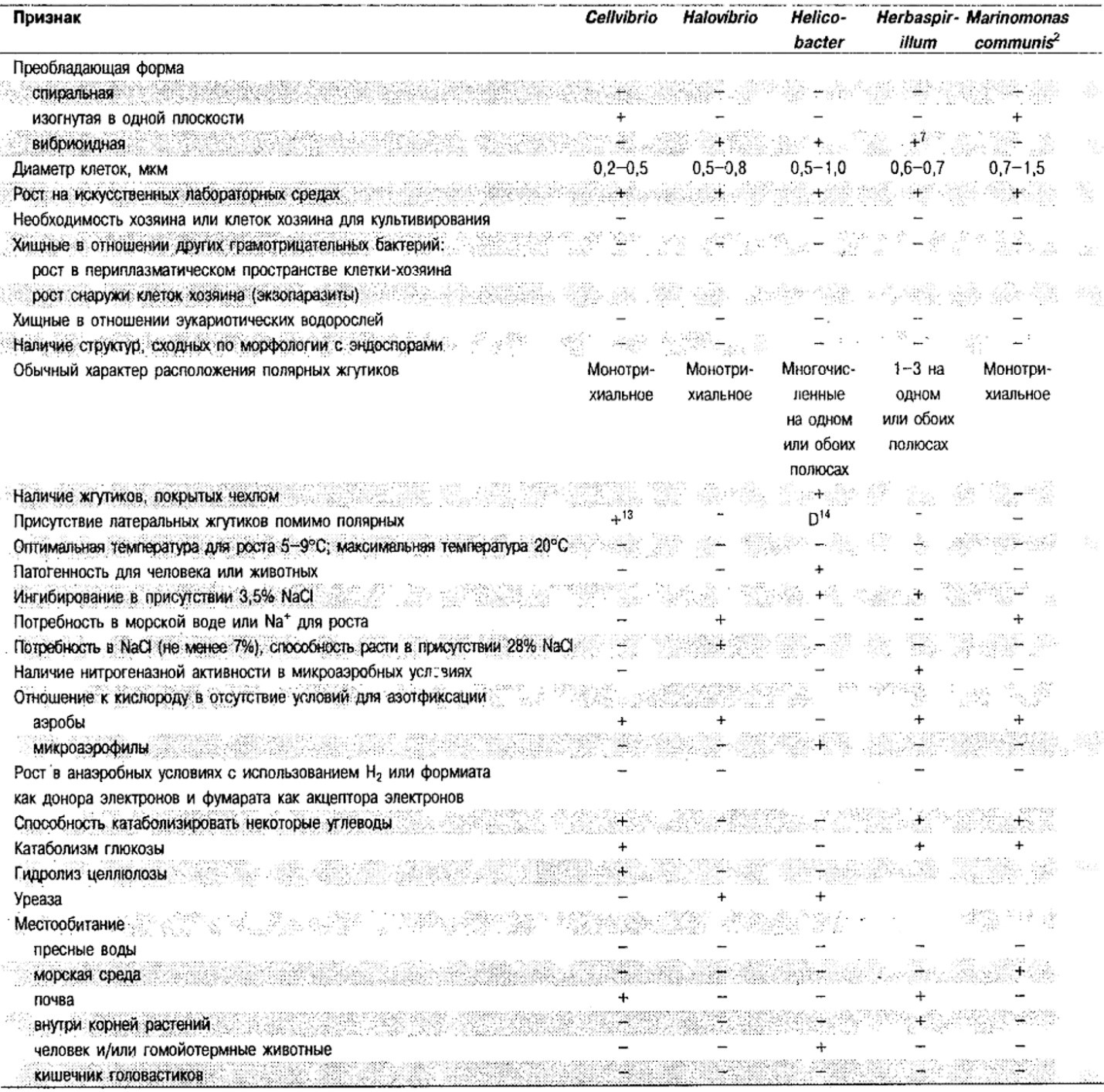
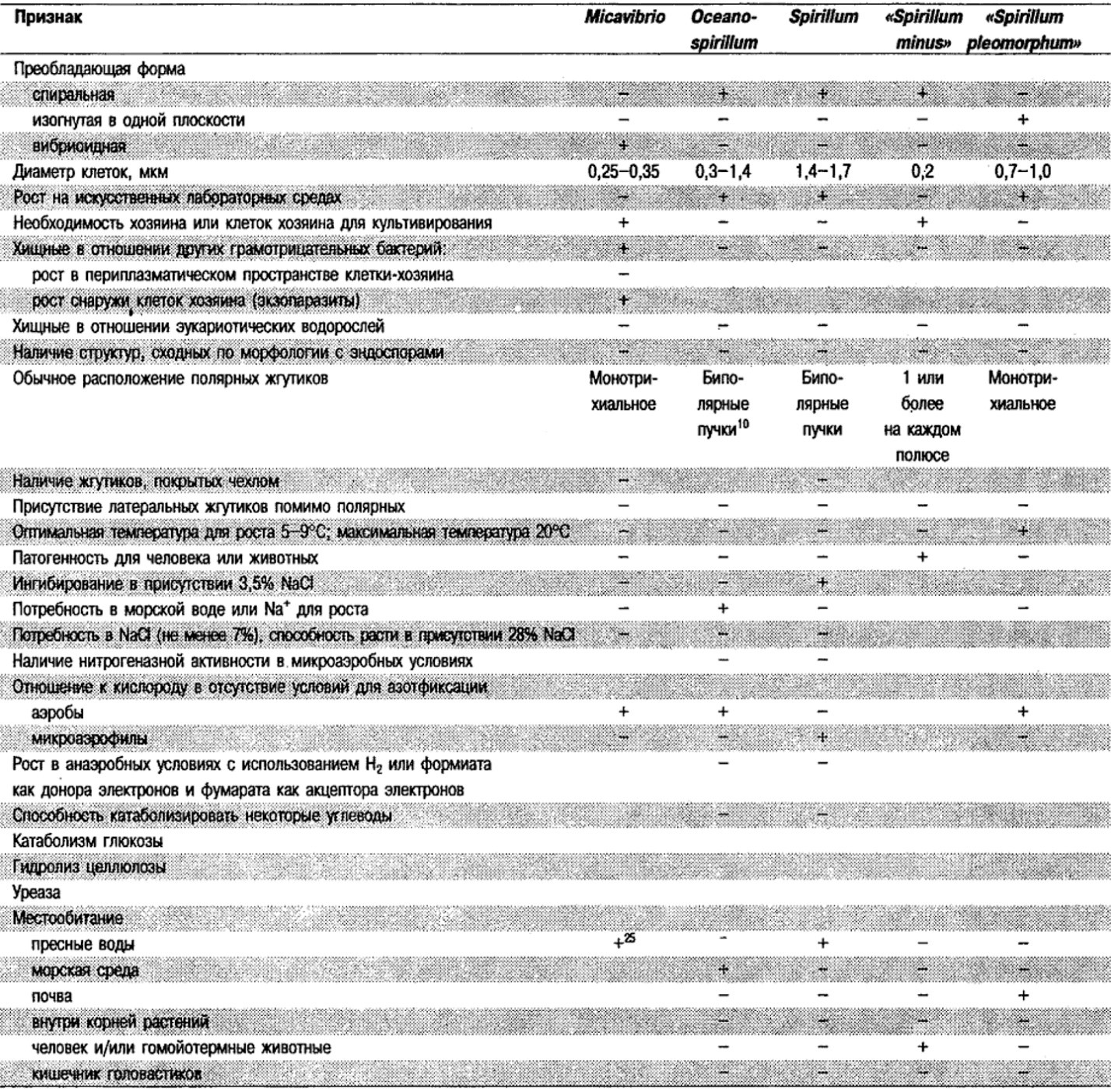
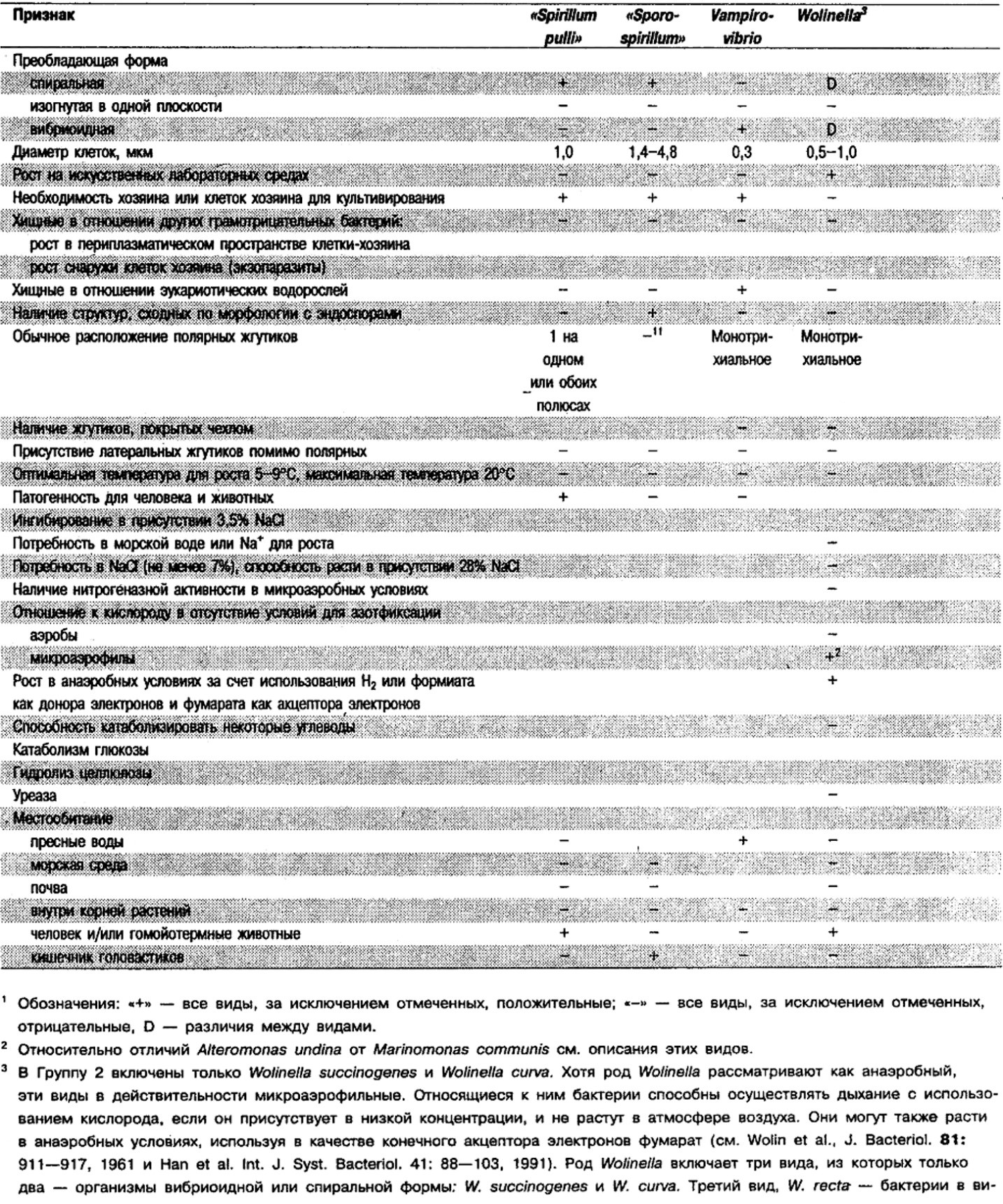

Род Alteromonas: Alteromonas undina
Клетки в форме изогнутых палочек, диаметром 0,7-1,5 и длиной 1,8-3,0 мкм. Поли-β-гидроксибутират в качестве внутриклеточного запасного вещества не накапливают. Микроцисты и эндоспоры не образуют. Грамотрицательные, Подвижные за счет одного полярно расположенного жгутика. Хемоорганотрофы; метаболизм чисто дыхательного типа с использованием О2 в качестве конечного акцептора электронов. Некоторые штаммы растут при 4°С, все – при 20°С, ни один – при 35°С. Колонии неокрашенные. Нитрат до нитрита не восстанавливают. Денитрификацию не осуществляют. Нуждаются в органических факторах роста. Основой для питательных сред должна быть морская вода. Образуют желатиназу, липазу, хитиназу, но не альгиназу; некоторые штаммы образуют амилазу. В качестве источника углерода используют сахарозу, мальтозу, ацетилглюкозамин, сукцинат, фумарат и тирозин. Не используют D-маннозу, D-галактозу, D-фруктозу, целлобиозу, мелибиозу, лактозу, эритритол, маннитол, глицерол, салицин, глюконат, DL-лактат, цитрат, D-сорбитол, DL-малат, α-кетоглутарат и м-гидроксибензоат. Аргининдигидролазную систему конститутивно не синтезируют. Встречаются в прибрежных водах и открытом океане.
Виды Alteromonas с клетками в форме прямых палочек рассмотрены в данном определителе в составе Группы 4.
Отличия Alteromonas undina от Marinomonas communis: Клетки A, undina в отличие от М. communis 1) обладают активностями желатиназы, липазы и хитиназы, 2) используют сахарозу, ацетил глюкозамин и тирозин и 3) не используют маннозу, фруктозу, глюконат, DL-лактат, маннитол, глицерол, сорбитол, DL-малат, α-кетоглутарат и м-гидроксибензоат.
Род Aquaspirillum (за исключением Aquaspirillum fasciculus)
Клетки ригидные, обычно спиральной формы, 0,2–1,4 мкм в диаметре, однако у одного вида вибриоидные. Как правило, содержат внутриклеточные гранулы поли-β-гидроксибутнрата. Некоторые виды образуют кокковндные тела с тонкой клеточной стенкой, преобладающие в старых культурах. Подвижные за счет полярных жгутиков, расположенных обычно пучками на обоих концах клетки, однако у одного вида жгутиков а ние монотрихиальное, а у некоторых жгутики расположены по одному на каждом конце. Один из видов обнаруживает таксис к магнитному полю. Аэробы или микроаэрофилы. Отдельные виды могут расти в анаэробных условиях, используя в качестве конечного акцептора электронов нитрат. Для роста большинства видов оптимальна температура 30-32°С. Хемоорганогетеротрофы, однако один вид – факультативный автотроф, использующий молекулярный водород. Оксндазоположительные. Обычно каталазо- и фосфатазоположительные, индол- и сульфатазоотрицательные. Казеин, крахмал и гиппурат не гидролизуют, В присутствии 3% NaCl не растут. Отдельные виды способны к денитрификации. Некоторые виды обладают нит- рогеназной активностью, но только в микроаэробных условиях. Источниками углерода служат аминокислоты и соли органических кислот. Углеводы обычно ие катаболизируют, однако некоторые виды могут разлагать небольшое число углеводов. В витаминах, как правило, не нуждаются. Обычно встречаются в стоячих пресноводных средах.
Типовой вид: Aquaspirillum serpens.
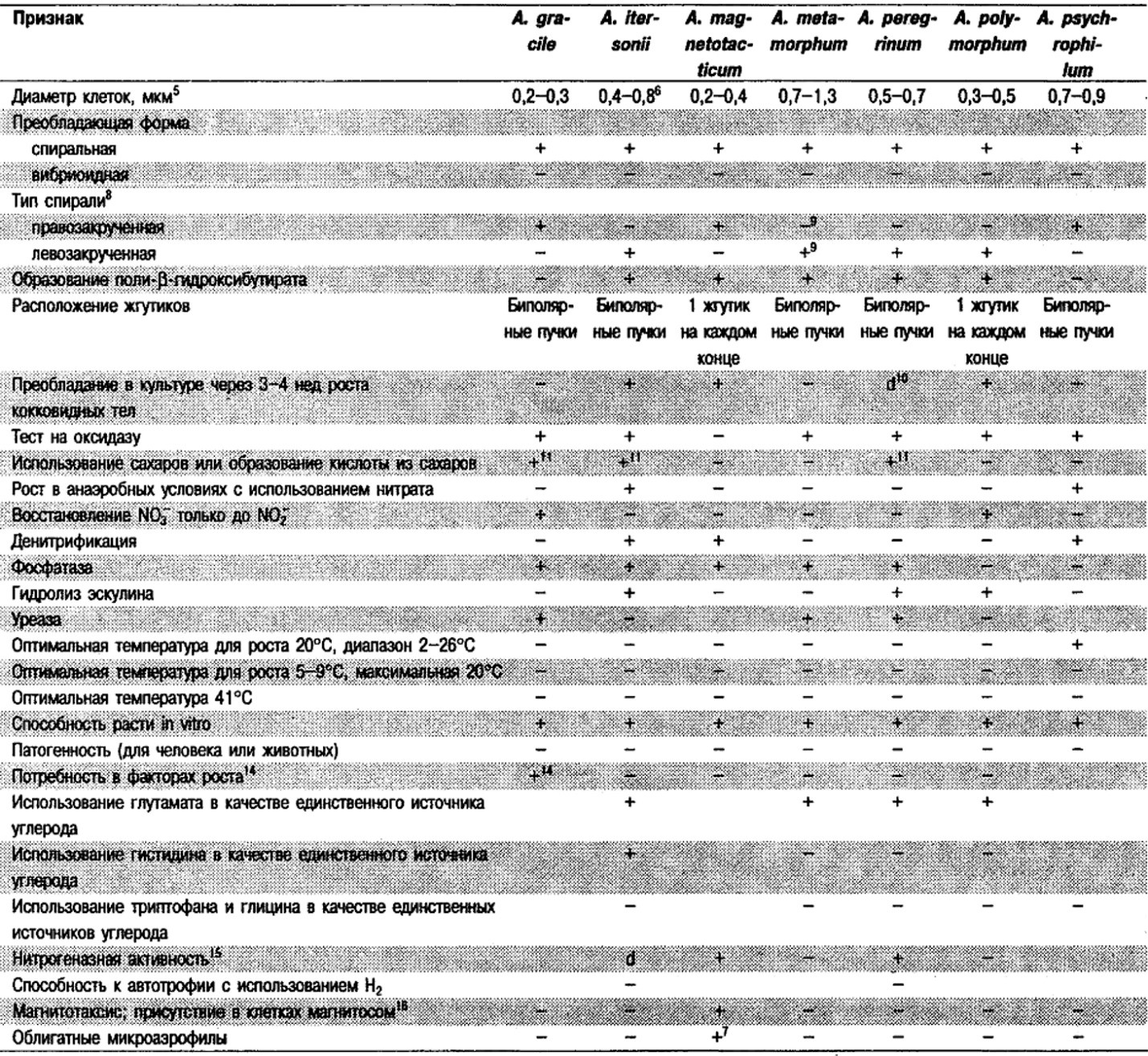
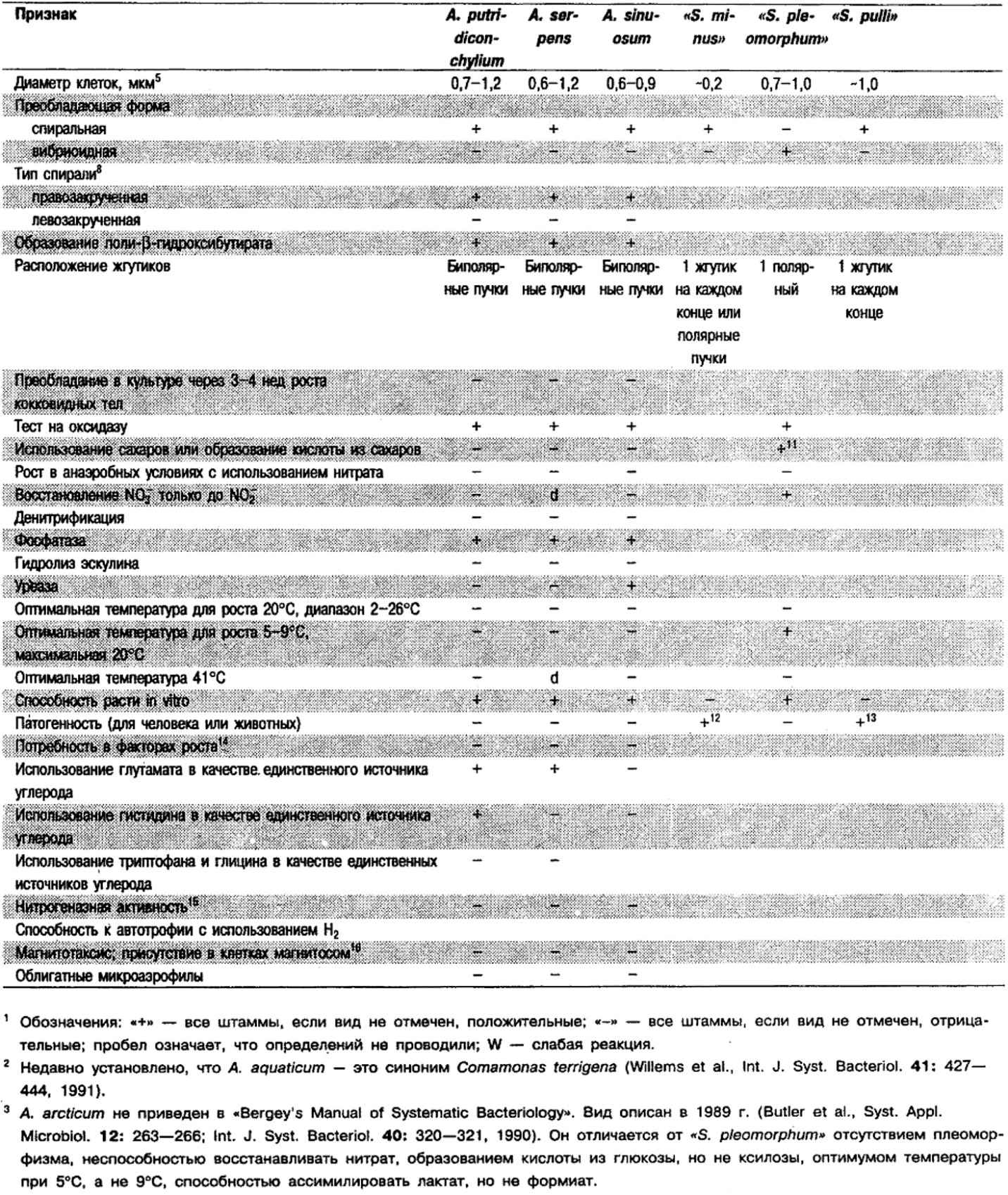
Дифференциация видов рода Aquaspirillum: См. табл. 2.2.
Таблица 2.2. Дифференцирующие признаки видов рода Aquaspirillum,
а также «Spirillum minus», «Spirillum pleomorphum» и «Spirillum pulii»1 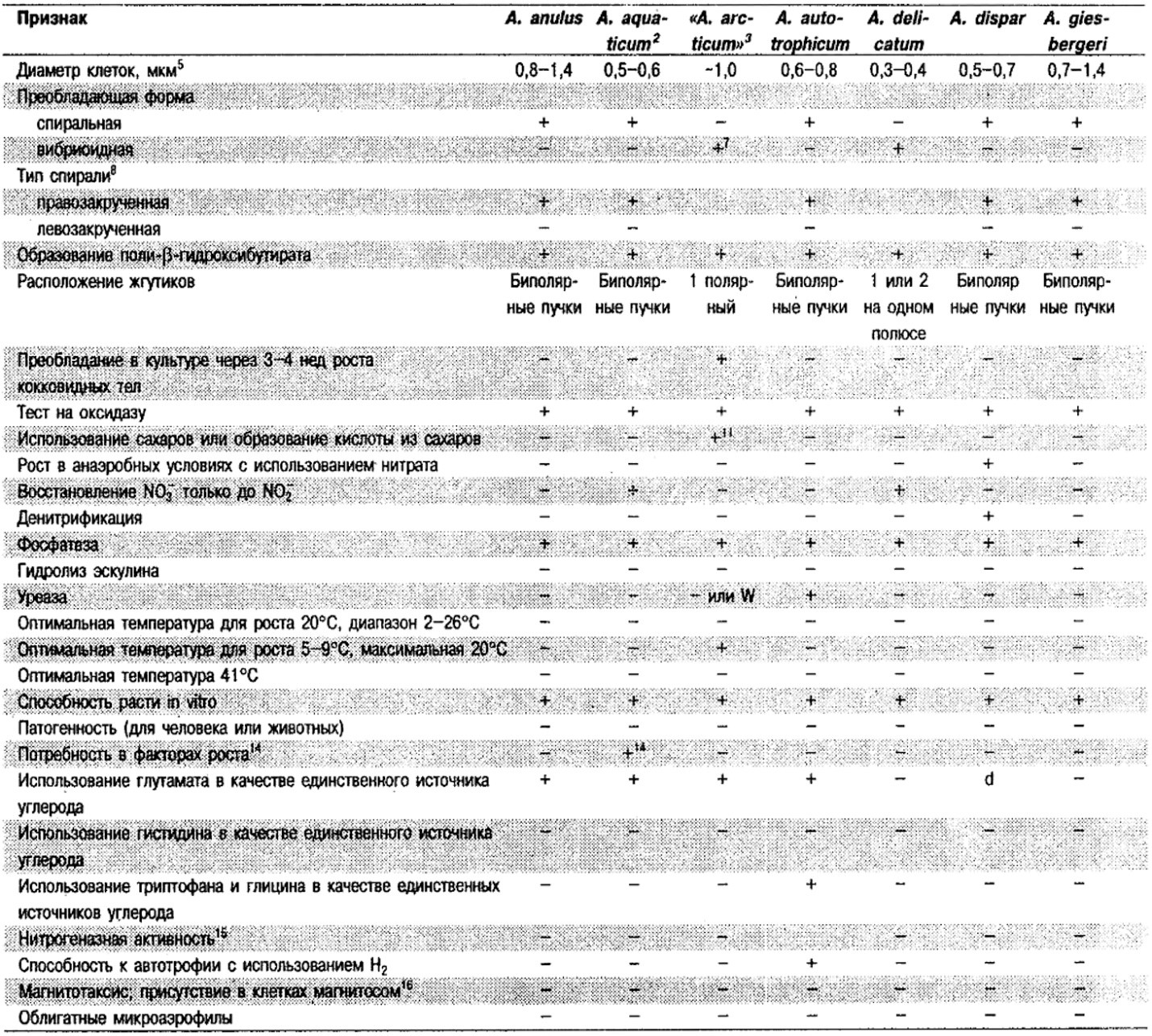

Род Azospirillum
Утолщенные вибрионы или прямые палочки, 0,9–1,2 мкм, часто с заостренными концами. Окрашивание но Граму от грамотрицателъного до грамвариабельиого. Содержат внутриклеточные гранулы поли-β-гидроксибутирата. Подвижные, в жидкой среде совершают характерное винтовое движение с помощью одного полярного жгутика. У некоторых видов при культивировании на плотной среде при 30°С помимо полярного жгутика образуются латеральные жгутики. Колонии некоторых штаммов на картофельном агаре имеют окраску от светло- до темно-розовой. Оптимальная температура для роста 34–37°С. Некоторые штаммы хорошо растут при рН 7, для других предпочтительны более кислые условия. Азотфиксаторы, способные к N2-зависимому росту в микроаэробных условиях. Хорошо растут в атмосфере воздуха в присутствии источника связанного азота, например, соли аммония. Метаболизм, как правило, дыхательного типа с использованием в качестве конечного акцептора электронов О2, а у некоторых штаммов и NO3–. Могут проявлять и слабую способность к брожению. В условиях сильного дефицита О2 некоторые штаммы могут осуществлять диссимиляционное восстановление NО3– до NО2– либо N2O и N2. Оксидазоположигельные. Хемоорганотрофы, однако некоторые виды – факультативные автотрофы, окисляющие молекулярный водород. Хорошо растут за счет использования солей органических кислот, таких как малат, сукцннат, лактат и пируват; могут использовать в качестве источника углерода и некоторые углеводы. Ряд штаммов нуждается в биотине. Встречаются в почве как свободножнвущне формы или к ассоциации с корнями трав, злаковых и клубневых растений. Образование корневых клубеньков не индуцируют.
Типовой вид: Azospirillum lipoferutn.
Дифференциация видов рода Azospirillum: См. табл. 2.3.
Таблица 2.3. Дифференцирующие признаки видов Azospirillum
и Herbaspirillum seropedicae1
Род Bdellovibrio
Бактерии – хищники по отношению к другим грамотрицательным бактериям. Клетки палочковидные, в форме запятой, диаметром 0,2–0,5 мкм, подвижные за счет одного полярного, покрытого чехлом жгутика. Имеют двухстадийный (по морфологическим и физиологическим параметрам) жизненный цикл, состоящий из стадии хищничества, во время которой клетки не размножаются, и репродуктивной стадии, протекающей внутри клеток-хозяев. Высокоподвижный бделловибрион, по-видимому, попадает в клетку-жертву в результате случайного столкновения: сильно ударяясь о клетку-жертву, как правило значительно превосходящую его по размерам, он прикрепляется к ней, а затем быстро проникает в периплазматическое пространство. Пораженная бделловибрионом клетка-жертва обычно округляется и раздувается, приобретая сферическую форму (бделлопласт). В самом начале нападения бделловибрион убивает свою жертву, после чего эта клетка уже служит субстратом для развития бделловибриона. Развиваясь за счет использования Протопласта жертвы, бделловибрион удлиняется, приобретая змеевидную форму. Неподвижная спиральная клетка распадается затем на подвижные, одинаковые по размерам вибрионы, которые покидают свою жертву, превратившуюся теперь в «тень» клетки, и начинают новый жизненный цикл.
Чтобы отнести бактерии к роду Bdellovibriot нужно убедиться в их способности расти в периплазматическом пространстве бактерий. Это главный признак, по которому данные разнородные бактерии, имеющие форму запятой, объединяют в одну группу. Установить такую способность можно с помощью фазово-контрастной световой или просвечивающей электронной микроскопии. Эти методы позволяют подтвердить локализацию бделловибрион о в внутри клетки-хозяина. Имеется описание соответствующей методики (Burnham et al., J. Bacteriol. 96: 1366-1368, 1968). Полезным для идентификации может также быть электронно-микроскопическое исследование негативно окрашенных препаратов, позволяющее подтвердить наличие покрытого чехлом жгутика.
Облигатные аэробы с чисто дыхательным типом метаболизма, использующие в качестве конечного акцептора электронов О2. Оптимальная для роста температура обычно 28-30°С, при температуре выше 37°С и ниже 10°С рост слабый. На начальных этапах выделения все природные изоляты растут только внутри периплазматического пространства клеток восприимчивой к ним жертвы. В отдельных случаях удается получить рост изолятов в отсутствие жертвы на среде с высокой концентрацией экстракта бактериальных клеток. Штаммы, выделенные из природных образцов, всегда представляют собой хищные формы бделловибрионов, однако впоследствии из хищных штаммов можно получить мутанты, способные к аксеничному росту («не зависящие от жертвы» штаммы). Некоторые штаммы – факультативно хищные, т, е. способны расти как в присутствии, так и в отсутствие клеток жертвы. Бделловибрионы встречаются в почве, сточных водах, а также в пресноводных и морских местообитаниях.
Типовой вид: Bdellovibrio bacteriovorus.
Дифференциация видов рода Bdellovibrio: См. табл. 2.4.
Таблица 2.4. Дифференцирующие признаки видов рода Bdeflovibrio1 
Помимо указанных трех видов описано большое число морских штаммов, форматно не отнесенных пока к определенным видам. Эти штаммы, выделенные из морской воды, отличаются от штаммов трех установленных видов своей потребностью в ионах Na+ (75 мМ или выше).
Чтобы более четко отличить В. bacteriovorus от В. stolpii и В. starrii, помимо признаков, указанных в табл. 2.4, иногда необходимо определить состав оснований ДНК у выделенных штаммов. Содержание G+C в ДНК у В. bacteriovorus составляет 50 мол.%, тогда как у В. stolpii – 42, а у В, starrii – 43,5 мол.%.
Род Campylobacter
Тонкие, вибриоидные клетки диаметром 0,2–0,5 мкм и длиной 0,5-5,0 мкм. Могут иметь один или больше витков спирали и достигать длины 8 мкм. Характерны также S-образная форма и форма типа крыльев чайки, возникающие при соединении двух клеток в короткую цепочку. Спор не образуют. В старых культурах часто присутствуют сферические формы (кокковидные тела). Грамотрицательные. Подвижные; совершают характерное винтовое движение с помощью одиночных жгутиков без чехла, расположенных на одном или обоих концах клетки. В большинстве микроаэрофнлы с дыхательным типом метаболизма. Как правило, растут при концентрации О2 3–15% и концентрации СО2 3-5%. Немногие штаммы слабо растут в аэробных условиях (21% О2). Один вид аэротолерантен и может расти в атмосфере воздуха. При росте в микроаэробных условиях некоторые штаммы нуждаются в Н2 или формнате. Некоторые виды растут также в анаэробных условиях в присутствии фумарата, либо фумарата + формиата, либо формиата + Н2. Хемоорганотрофы. Углеводы не сбраживают и не окисляют. Кислых или нейтральных конечных продуктов не образуют. В крови или сыворотке для роста не нуждаются. Для получения энергии используют аминокислоты или интермедиаты цикла трикарбоновых кислот, но не углеводы. Желатину не гидролизуют. Проба с метиловым красным и реакция Фогеса-Проскауэра отрицательные, Липазной активностью не обладают. Оксидазоположительные и уреазоотрицательные за исключением некоторых штаммов С. lari. Пигментов не образуют. В составе клеточных хинонов преобладают менахинон-6 и метил-замещенный менахинон-6. В составе жирных кислот у большинства видов доминирует гексадекановая кислота. Некоторые виды патогенны для человека и животных. Обнаруживаются в репродуктивных органах, кишечном тракте и ротовой полости человека н животных. Один вид – азотфикеирующие бактерии, выделенные из корней растений, растущих на соляных маршах.
Типовой вид: Campylobacter fetus.
Дифференциация видов рода Campylobacter: См. табл. 2.5.
Таблица 2.5. Дифференцирующие признаки видов рода Campylobacter,
а также Wolmella succinogenes и Wolinella curve1 



Род Cellvibrio
От редакции; Род Cellvibrio не приведен в «Вегgey's Manual of Systematic Bacteriology». Он описан в 1985 г. (Blackall et al. J. Appl. Bacteriol. 59: 81-97; Int. J, Syst. Bacteriol. 36: 354-356, 1986) и содержит единственный вид С. mixtus.
Слегка изогнутые палочки, 0,2-0,5 х 1,0-1,3 мкм. Грамотрицательные; подвижные. Имеют один полярно расположенный жгутик, а при росте на плотной среде образуют также до 11 более тонких латерально расположенных жгутиков. Аэробы; каталазо- и оксидазоположительные. Окисляют глюкозу, гидролизуют целлюлозу. Органические кислоты в качестве единственного источника углерода не используют. В факторах роста не нуждаются.
Типовой (и единственный) вид: Cellvibrio mixtus.
Признаки вида: Клетки растут на агаризованных средах: с пептоном, дрожжевым экстрактом и 5% сахарозы; с сахарозой и пептоном; с глюкозой и дрожжевым экстрактом; с глюкозой и сульфатом аммония, однако плохо растут или не растут на агаризованной среде только с пептоном и дрожжевым экстрактом. На агаризованной среде с сахарозой и пептоном образуют колонии диаметром 1–3 мм, от белых до кремовых, гладкие, от слабовыпуклых до выпуклых, просвечивающие, слизистые, округлые, с цельным краем или желто-оранжевые, матовые, плоские, просвечивающие, округлые, с краем неправильной формы. На синтетической минеральной среде с целлюлозой все штаммы образуют белые, гладкие, округлые колонии с цельным краем, окруженные узкой зоной просветления. Образуют кислоту из L-арабинозы, декстрина, галактозы, глюкозы, лактозы, мальтозы, маннозы, мелибиозы, рафинозы, сахарозы, трегалозы и ксилозы, но не из адонитола, арабитола, D-арабинозы, дулъцитола, эритритола, глицерола, инозитола, маннитола, рибозы, сорбитола, сорбозы или этанола. Гидролизуют хитин, крахмал, эскулин, а также щелочной или нейтральный пектиновый гель. Желатину, казеин, ДНК, Твин-80 и кислый пектиновый гель не гидролизуют. Образуют α-глюкозидазу, β-глюкозидазу и β-ксилозидазу. Уреазу, α-фукозидазу, аргининдигидролазу, арилсульфатазу, ациламидазу и фенилал аниндезаминазу не образуют. Из глюкозы синтезируют полисахарид курдлан. Не образуют: 3-кетолактозу из лактозы, Дигидроацетон из глицерола, индол из триптофана, H2S из цистеина и газ из нитрата. Результаты тестов с метиловым красным и Фогеса-Проскауэра отрицательные. Глюконат не окисляют. Не растут на цетримидном агаре, в анаэробных условиях, при 42°С, на безазотистой среде с глюкозой, минеральной среде с алкиламинами, минеральной среде с фенолом и в присутствии 0,0075% KCN. Подвид С. mixtus subsp. dextranolyticus гидролизует декстран, подвид С. mixtus subsp. mixtus – нет.
Род Halovibrio
От редакции: Род Halovibrio не приведен в «Bergey's Manual of Systematic Bacteriology». Он описан в 1988 г. (Fendrich, Syst. Appl. Microbiol. 11: 36-43; Int. J. Syst. Bacteriol., 39: 205-206, 1989) и содержит единственный вид H. variabilis.
Облнгатно аэробные, извитые, грамотрицател ь- ные, палочковидные клетки, растущие при концентрации NaCI в среде не менее 1,2 М (7,0%, масса/объем). Подвижные за счет одного полярного жгутика. Хемоорганотрофы. Углеводы не окисляют.
Типовой (и единственный) вид: Halovibrio variabilis.
Признаки вида: Клетки вибриоидной формы, 0,5-0,8 х 1,0-3,0 мкм, одиночные или в парах. В старых культурах могут присутствовать кокковидные тела. Галофилы, растут при концентрации NaCI oт 1,2 М (7,0%, масса/объем) до 4,9 М (28,6%, масса/объем); оптимальная концентрация NaCI 16 М (9,4%, масса/объем). Диапазон рН 6,5–8,4, оптимум 7,5; диапазон температуры 15-37, оптимум ЗЗсС. Колонии округлые, с ровным краем, слизистые, приподнятые, светло-коричневые. Субстратами для роста служат ацетат, капронат, глутамин, валерат, эскулин, глицерол, этанол, казаминовые кислоты, пептон, дрожжевой экстракт. Каталазо- и оксидазоположитель- ные. Желатину не разжижают. Образуют уреазу. Крахмал и целлюлозу не гидролизуют. Индол и H2S не образуют. Нитрат не восстанавливают. По аргининдигидролазе, лизинде карбоксилазе и орнитиндекарбоксилазе отрицательные. Обнаружены в Северном заливе Большого Соленого озера, шт. Юта, США.
Род Helicobacter
От редакции: Род Helicobacter не приведен в «Bergey's Manual of Systematic Bacteriology». Он описан в 1989 г. (Goodwin et al., Int. J. Syst. Bacteriol. 39: 397-405) и включает два вида: Н pyroli и Н. mustelae.
Клетки в форме спиральных, изогнутых или прямых палочек, диаметром 0,5–1,0 мкм и длиной 2,5–5,0 мкм, с закругленными концами. При росте в жидкой среде на качалке образуют внеклеточный гликокаликс. Движение клеток быстрое и стремительное, за счет многочисленных жгутиков с чехлом, расположенных на одном или на обоих полюсах клеток, а также латералъно и имеющих на концах вздутия. Микроаэрофилы. В атмосфере воздуха, обогащенного 10% СО2, и в анаэробных условиях рост вариабельный. Без перемешивания в жидкой среде с сердечно-мозговой вытяжкой (СМВ) и других жидких средах растут медленно, на СМВ-агаре и шоколадном агаре дают рост через 2–5 сут. Растут также на СМВ-агаре с древесным углем или кукурузным крахмалом. Колонии неокрашенные, просвечивающие, диаметром 1–2 мм. Оптимальная для роста температура – 37°С, рост происходит при 30, но не 25°С, при 42°С рост нестабильный. Растут в присутствии 0,5% глицина и 0,04% трифенилтетразолийхлорида. В присутствии 3,5% NaCl не растут. Каталазо- и оксидазоположительные. Быстро гидролизуют мочевину. Основные изопреноидные хиноны в составе клеток – МК-6 и неидентифицированный МК-6 (Un-МК-6). В составе жирных кислот преобладает гексадекановая кислота. Результаты теста на образование H2S отрицательные при постановке теста на тройном сахарном агаре с железом и вариабельные при использовании бумаги, пропитанной ацетатом свинца. По способности к восстановлению нитрата и гидролизу гиппурата вариабельные. По щелочной фосфатазе и ץ-глутамилтранспептидазе положительные, по лейцинариламидаэе вариабельные. Чувствительны к пенициллину, ампициллину, амоксициллину, эритромицину, гентамицину, канамицину, рифампицину и тетрациклину. Нечувствительны к ванкомицину, сульфамидам и триметоприму. По устойчивости к налидиксовой кислоте, цефалотину, метронидазолу и полимиксину вариабельные. Выделены из слизистой желудка приматов и африканских хорьков. Некоторые представители рода, возможно, ассоциированы с гастритом и язвой желудка.
Типовой вид: Helicobacter pylori.
Дифференциация видов рода Helicobacter: См. табл. 2.6.
Таблица 2.6. Дифференцирующие признаки видов рода Helicobacter1 
Род Herbaspirillum
От редакции: Род Herbaspirillum не приведен в «Bergey's Manual of Systematic Bacteriology». Он описан в 1986 г. (Baldani et al., int. J. Syst. Bacteriol. 36: 86-93) и содержит один вид H. seropedicae.
Клетки обычно извитой» иногда спиральной формы, диаметром 0,6–0,7 мкм. Грамотрицательные. Подвижные за счет 1–3 жгутиков, расположенных на одном или обоих полюсах клетки. Колонии на картофельном агаре коричневатые в центре, приподнятые, гладкие. Оптимальная температура для роста 34°С, диапазон рН 5,3-8,0. Азотфнксаторы, растущие за счет использования N2 в микроаэробных условиях. В присутствии источника связанного азота, например соли аммония, хорошо растут в атмосфере воздуха. Обладают дыхательным типом метаболизма. В анаэробных условия» в присутствии нитрата не растут. При сильном дефиците О2 осуществляют диссимиляционное восстановление NO3– до NO2–, а некоторые штаммы – до N2О. Оксидазоположительные. Хемоорганотрофы. Хорошо растут за счет использования солей органических кислот, таких как малат, фумарат, сукцинат, пнруват, цитрат; источниками углерода могут также служить некоторые углеводы. В витаминах не нуждаются. Встречаются в почве в свободноживу- щей форме или в ассоциации с корнями растений, относящихся к семейству Gramineae.
Типовой (и единственный) вид: Herbaspirilllum seropedicae.
Признаки вида: Клетки обычно имеют два полярных жгутика (в отдельных случаях от одного до трех) на одном или обоих полюсах. При 35°С на мягком агаре происходит выраженное роение, однако латеральные жгутики, подобные описанным у ряда видов рода Azospirillum, отсутствуют. Эффективность азотфиксации при росте на полужидкой среде без азота составляет 12-15 мг N на 1 г DL-малата или 13 мг N на 1 г маннитола. Скорость роста в условиях азотфиксации ниже, чем у видов Azospirilhim, однако в присутствии минерального азота или глутамата намного выше. В присутствии 2% NaCl не растут. В качестве источников углерода могут использовать глюкозу и α-кетоглутарат, но не сахарозу. Оптимальная температура для роста в условиях азотфиксации – 34°С; при 22 и 38°С не растут.
Род Marinomonas: Marinomonas communis
От редакции: Вид Marinomonas communis приведен в «Bergey's Manual of Systematic Bacteriology» под своим прежним названием Altervmonas communis. После пересмотра классификации в 1983 г. он перенесен в род Marinomonas (Van Landschoot, De Ley, J, Gen. Microbiol. 129: 3057-3074; Int. J. Syst. Bacteriol. 34: 91-92, 1984).
Изогнутые палочки диаметром 0,7–1,5 мкм и длиной 1,8-3,0 мкм. Поли-β-гидроксибутират не накапливают. Микроцист и эндоспор не образуют. Грамотрицательные. Подвижные за счет одиночных полярных жгутиков. Хемоорганотрофы; метаболизм чисто дыхательного типа; в качестве конечного акцептора электронов используют молекулярный кислород. Денитрифнкацию не осуществляют. В органических факторах роста не нуждаются. Нуждаются для роста в морской воде как основе питательной среды. Растут при 20 и 40°С и не растут при 4°С. Колонии не окрашены. Нитрат до нитрита не восстанавливают. По амилазе, желатиназе, липазе, альгиназе и хитиназе отрицательные. Используют следующие источники углерода: D-маннозу, D-фруктозуу D-глюконат, сахарат, мезо-инозитол, сукцинат, фумарат, DL-лактатт цитрат, аконитат, D-маннитол, глицерол, ץ-аминобутират, саркозин, путресцин, D-сорбитол, DL-малат, а-кетоглутарат, хиннат, D-аланин, L-орнитин, бетаин, м-гидроксибензоат и n-гидроксибснзоат; не используют сахарозу, целлобиозу, мелибиозу, лактозу, салицин, N-ацетилглкжозамин, эритритол, L-треонин и L-тирозин. Встречаются в прибрежных водах и открытом океане.
Клетки другого вида Marinomonas, М. vaga, имеют форму прямых палочек; этот вид помещен в Группу 4.
Отличия Marinomonas communis от Alterumonas undina. Представители Мcommunis в отличие от A. undina 1) не обладают желатиназной, липазной и хитиназной активностями; 2) не используют сахарозу, ацетил глюкозам ин и тирозин, 3) используют маннозу, фруктозу, глюконат, DL-лактат, маннитол, глицерол, сорбитол, DL-малат, α-кетоглутарат и м-гидроксибензоат.
Род Micavibrio
От редакции: Род Micavibrio не приведен в «Bergey's Manual of Systematic Bacteriology». Он описан в 1982 г. (Ламбина и др., Микробиология, 51: 114-117; Lambina et al., Int. J. Syst. Bacteriol. 38: 93-94; 1989) и содержит один вид М. admirandus.
Клетки палочковидные, в форме запятой, диаметром 0,2–0,3 мкм. Грамотрицательные. Движутся с помощью одного жгутика, не покрытого чехлом (в отличие от жгугика Bdellovibrio). Хищники по отношению к бактериям Xanthomonas maltophilia (единственный известный на сегодняшний день хозяин). Прикрепляясь снаружи к клеткам хозяина, теряют подвижность и развиваются как экзопаразиты, лизируя клетки хозяина. В отличие от бделловибрионов, не проникают в периплазматическое пространство клетки-хозяина и не имеют в жизненном цикле стадии образования удлиненной спиральной формы. Не чувствительны к виб- риостатическому птеридину О/129 и не могут расти в отсутствие бактерии-хозяина.
Типовой (и единственный вид): Micavibrio admirandus.
Признаки вида: Как для рода.
Род Oceanospirillum
От редакции: Клетки Oceanospirillum kriegii и Oceanospirillum jannaschii имеют форму прямых палочек. В данном определителе эти виды помещены в Группу 4. Приведенное ниже описание рода на эти виды не распространяется.
Ригидные клетки спиральной формы, диаметром 0,3–1,4 мкм. Накапливают поли-β-гидроксибутират. Большинство видов образует кокковидные тела с тонкой оболочкой, которые преобладают в старых культурах. Грамотрицательные, Подвижные за счет биполярных пучков жгутиков нлн двух жгутиков, по одному на каждом конце клетки. Аэробы; метаболизм чисто дыхательного типа с использованием в качестве конечного акцептора электронов О2. Нитратное дыхание не осуществляют. Оптимальная температура для роста 25–32°С. Оксидазоположительные; индол и арилсульфатазу не образуют. Казеин, крахмал, гипгтурат и эскулин не гидролизуют. Для роста нуждаются в морской воде. Углеводы не окисляют и не сбраживают. В качестве единственного источника углерода используют аминокислоты либо соли органических кислот. В факторах роста обычно не нуждаются. Выделены из прибрежной морской воды, разлагающихся морских водорослей и гниющих остатков морских моллюсков.
Типовой вид: Oceanospirillum linum.
Дифференциация видов рода Oceanosptrillum: См. табл. 2.7.
Таблица 2.7. Дифференцирующие признаки видов рода OceanospiriHum1 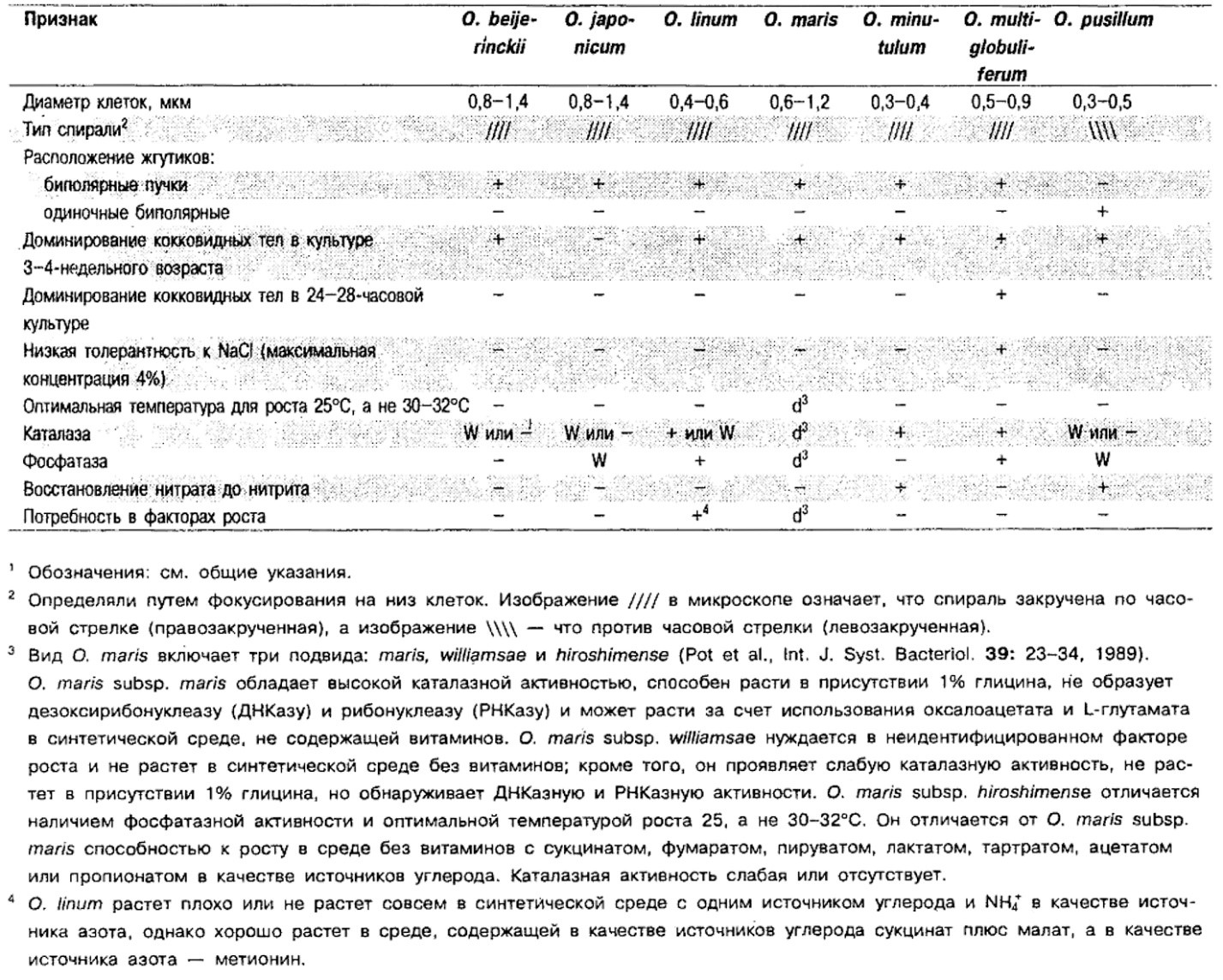
Род Spirillum
Ригидные клетки спиральной формы, диаметром 1,4–1,7 мкм и длиной 14–60 мкм. Содержат включения поли-β-гидроксибутирата. Кокковидных тел не образуют. Грамотрицательные. Подвижные за счет крупных биполярно расположенных пучков жгутиков, имеющих протяженную длину волны и образующих примерно один виток спирали; они хорошо различимы с помощью темнопольной и фазово-контрастной микроскопии. Обычно растут как микроаэрофилы, но могут расти аэробно в специальных жидких средах или средах с особыми добавками. Колонии на поверхности плотных сред можно получить только при использовании специальных сред и только в микроаэробных условиях. Метаболизм чисто дыхательного типа с использованием в качестве акцептора электронов О2. В анаэробных условиях при наличии нитрата в среде не растут. Оптимальная температура для роста 30°С. Оксндазо- и фосфатазоположительные. Катала зоотркцательные. Индол- и сульфатазоотрицательные. Казеин, крахмал, эскулин, желатину, ДНК и РНК не гидролизуют. Н2О2 в очень низких концентрациях, а также NaCl и фосфат в концентрациях выше 0,02% и выше 0,01 М соответственно ингибиру- ют рост. Углеводы не катаболизнруют, В качестве источников углерода используют соли ряда органических кислот, предпочтительно сукцинат калия. В витаминах не нуждаются. Встречаются в стоячей пресной воде.
Типовой вид: Spirillum volutans.
Признаки Spirillum volutans: Как для рода.
«Spirillum minus»
Ригидные клетки, обычно в виде спирали из двух или трех витков, хотя бегущая волна описана как плоская. Концы клеток тупые или заостренные. Диаметр клеток примерно 0,2 мкм, длина 3–5 мкм, длина волны 0,8–1,0 мкм. Активно подвижные за счет одного или более жгутиков, расположенных на обоих полюсах. У человека S. minus вызывает болезнь «содоку» – одну из двух форм лихорадки от укусов крыс. Лучше всего видны в свежих мазках крови или экссудатов больных с помощью темнопольной или фазово-контрастной микроскопии; можно также использовать окрашивание красителем Гимза или Райта либо импрегнацию серебром. Неясно, удавалось ли когда-нибудь культивировать этот организм на искусственных средах. Его можно выращивать in vivo, вводя внутрибрюшинно кровь больного, либо экссудат из пораженного участка, либо кровь зараженных естественным путем крыс не зараженным спириллами мышам или морским свинкам.
«Spirillum pleomorphum»
Спиральные клетки, изогнутые палочки, серповидные клетки, 0,7-1,0 х 2,0-4,5 мкм. На агаризованной среде с пептоном, дрожжевым экстрактом и глюкозой образуют U-образные клетки и почти кольцевые формы. Подвижные за счет одного полярно расположенного жгутика. Колонии на агаре с пептоном, дрожжевым экстрактом и глюкозой округлые, гладкие, выпуклые, непрозрачные, с цельным краем, бледно-коричневые. Оптимальная температура для роста 9°С, максимальная 20°С, минимальная – ниже 0°С. Аэробы; каталазо- и оксидазоположительные. Результаты тестов на образование индола, Фогеса-Проскауэра и с метиловым красным отрицательные. В присутствии 5% NaCl не растут. Образуют (в аэробных условиях) кислоту, но не газ из ксилозы. Из лактозы, сахарозы, мальтозы, арабинозы или глицерола ни в аэробных, ни в анаэробных условиях кислоту или газ не образуют. Цвет лакмусового молочка не изменяют. H2S не образуют. Крахмал и целлюлозу не разлагают. Восстанавливают нитрат до нитрита. В анаэробных условиях на среде с нитратом не растут. Ассимилируют сукцинат, формиат, ацетат, фумарат и пропионат, но не цитрат, лактат, протокатехоат, n-гидроксибензоат или гиппурат. Выделены из антарктической почвы.
«Spirillum pulli»
Ригидные спиральные клетки. При темнопольной микроскопии.диаметр клеток 1 мкм, длина 5–12 мкм. Активно подвижные за счет двух жгутиков, по одному на каждом конце клетки. Считаются возбудителями дифтериеподобного стоматита ротовой полости у кур. На искусственных питательных средах культивировать не удается. В эксперименте заражение кур происходит путем контакта и искусственной иноксуляции.
Род «Sporospirillum»
Ригидные спиралевидные бактерии огромных размеров, диаметром 1,8-4,8 мкм и длиной 40-100 мкм. Клетки содержат структуры, сходные по морфологии с эндоспорами. Эти спороподобные структуры могут вращаться и перемещаться внутри цитоплазмы бактерии. Вначале они формируются у полюсов клетки, а затем мигрируют к центру, откуда высвобождаются после разрыва и разрушения клетки. Об окраске по Граму данных нет. Клетки подвижные, однако обеспечивающие движение органеллы не обнаружены. Отношение к О2 неизвестно. Обнаружены в кишечнике головастиков; в культуру не выделены.
Типовой вид: не установлен.
Дифференциация видов рода Sporospirillum: Поскольку культур данных организмов не получено, идентификация видов проводится только по морфологическим признакам, указанным в табл. 2.8.
Таблица 2.8. Дифференцирующие признаки видов рода «Sporospirillum» 
Род Vampirovfbrio
Палочковидные клетки в форме запятой, диаметром 0,3 мкм. Описаны также более крупные (0,6 мкм) кокковидные формы. Грамотрицательные. Подвижные за счет одного полярного, не покрытого чехлом жгугика. Размножение происходит путем бинарного деления. Клетки прикрепляются к клеткам штаммов водорослей Chloretta; для роста нужны живые клетки водорослей. Растут снаружи клеток-хозяев; о проникновении внутрь клетки-хозяина сообщений нет. Стадия жизненного цикла с образованием удлиненной спиральной формы клеток отсутствует.
Типовой (и единственный) вид: Vampirovibrio chlorellavorus.
Признаки вида: Как для рода.
Род WolineJIa: Wolirteila succinogenes и Wolinella curva
Спиральные или изогнутые клетки, 0,5–1,0 х 2–6 мкм, на концах суженные. Спор не образуют. Подвижные за счет одного полярно расположенного жгутика. Хотя в «Beigey's Manual of Systematic Bacteriology», т. 1, виды Wolinella описаны как анаэробы, на самом деле они микроаэрофилы, использующие Н2 и формиат и способные к кислородному дыханию. В отсутствие альтернативных конечных акцепторов электронов, например фумарата, растут за счет потребления кислорода при низкой его концентрации, но не растут в аэробных или анаэробных условиях (относительно W. succinogenes см. Wolin et al., J. Bacteriol. 81: 911–917, 1961; относительно W. curva см. Han et al., Int. J. Syst. Bacteriol. 41: 218-222, 1991; см. также описание Wolinella recta – вида бактерий, имеющих форму прямых палочек – в Группах 4 и 6 данного определителя). В качестве акцепторов электронов при анаэробном дыхании используют фумарат и нитрат. Оксндазоположительные и каталазо отрицательные. Содержат цигохромы b и с. Углеводы не используют. Выделены из рубца жвачных, а также у человека – из десневой борозды, каналов зубных корней при инфекции и другого клинического материала.
Типовой вид: Wolinella succinogenes.
Различие между Wolinella succinogenes и Wolinella curva: См. табл. 2.5.
Необходимые указания для пользования данным определителем
Если в сносках под таблицей нет других пояснений, то используемые обозначения имеют следующий смысл:
«+» – 90% и более штаммов положительные
«–» – 90% и более штаммов отрицательные
«d» – 11-89% штаммов положительные
«v» – нестабильность на уровне штамма (не равнозначно «d»)
«D» – различные реакции у разных таксонов (видов одного рода или родов одного семейства)
Для других обозначений пояснения приведены в сносках под таблицами.
