Эти грамотрицательные бактерии образуют весьма гетерогенную по морфологическим и физиологическим признакам группу. Однако все они обладают чисто дыхательным типом метаболизма и используют в качестве конечного акцептора электронов кислород. Почти все представители (за исключением микроаэрофилов) растут в атмосфере воздуха (21% О2). Для некоторых родов характерна способность к анаэробному дыханию с использованием нитрата, фумарата или других конечных акцепторов электронов. Бактерии нескольких родов способны фиксировать молекулярный азот.
Ключ для дифференциации основных подгрупп в Группе 4:
I. Аэробы. Палочки и кокки, способные расти в атмосфере воздуха (21% О2). Представители некоторых родов в условиях азотфиксации растут как микроаэрофилы, однако при наличии в среде источника связанного азота – как аэробы. Подгруппа 4А
II. Микроаэрофилы. Прямые палочки, не способные расти в атмосфере воздуха (21% О2). При пониженной концентрации О2 в газовой фазе и в отсутствие альтернативных акцепторов электронов растут за счет использования кислорода. Некоторые могут также осуществлять анаэробное дыхание с нитратом, фумаратом или другими конечными акцепторами электронов. Подгруппа 4В
ПОДГРУППА 4А
Дифференциация родов в Подгруппе 4А. Большинство дифференцирующих признаков родов представлено в табл. 4.1. Некоторые дополнительные дифференцирующие признаки для некоторых физиологических групп приведены в табл. 4.2-4.9.
Таблица 4.1. Дифференцирующие признаки родов
грамотрицательных аэробных палочковидных и корковидных эубактерий1 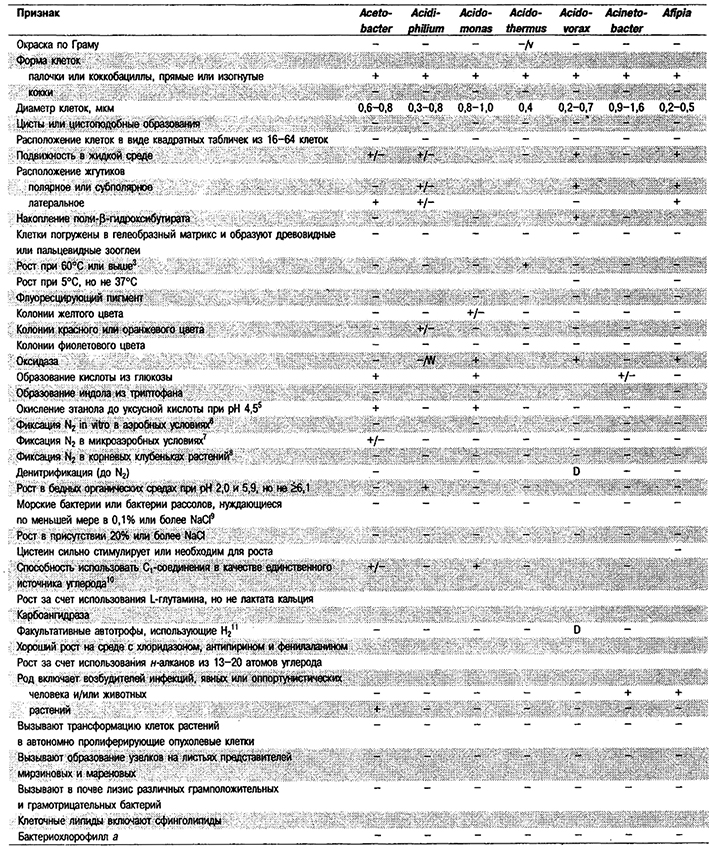

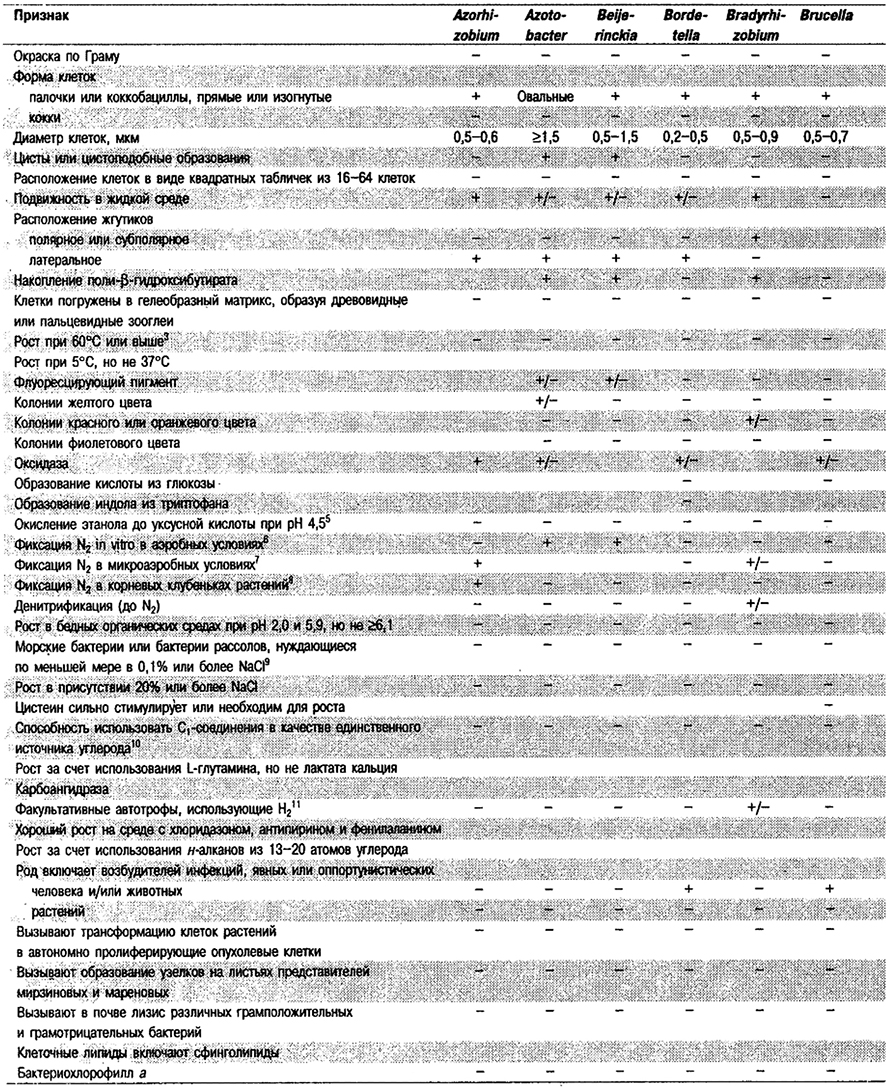
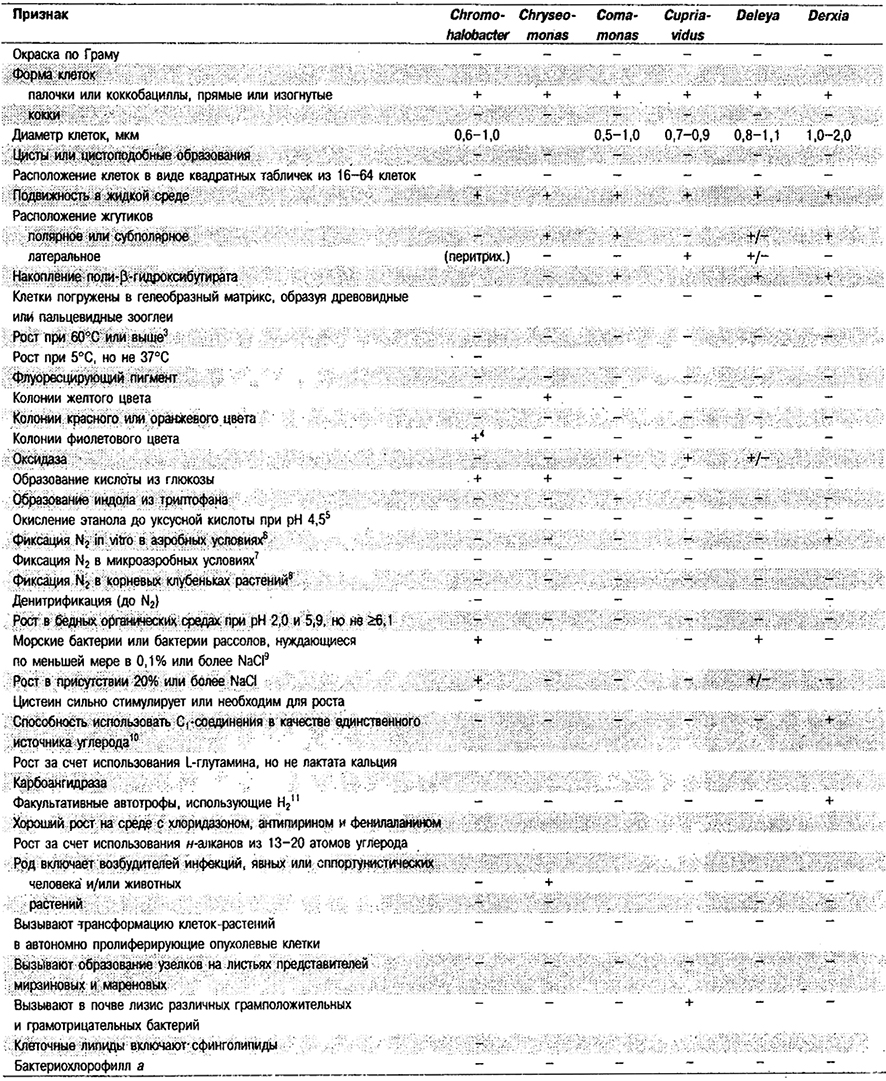

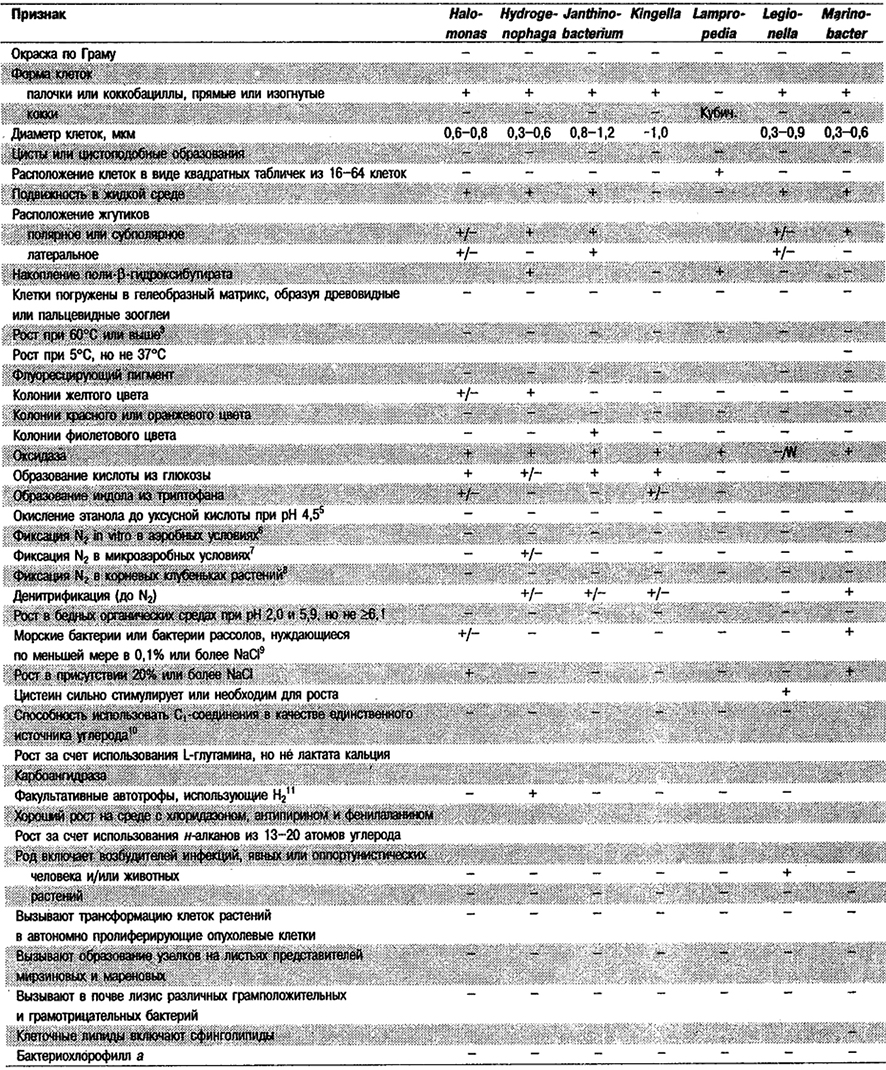
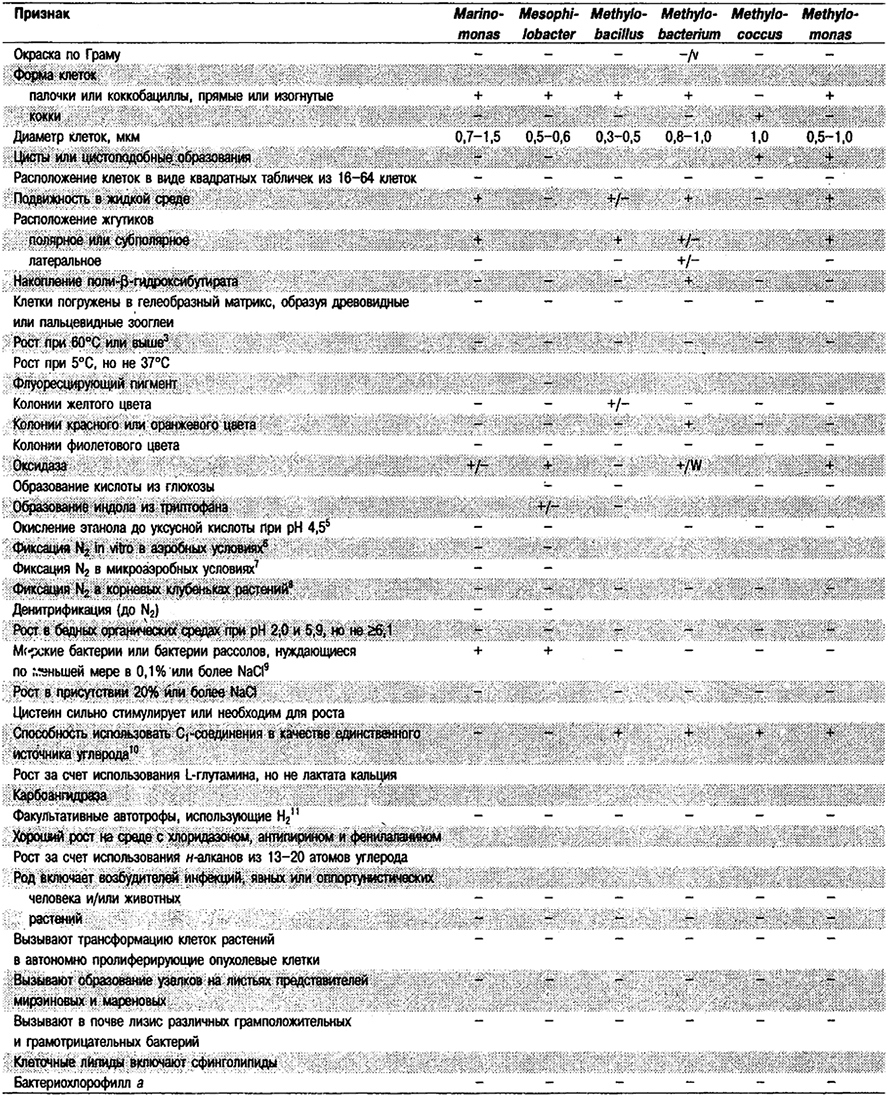

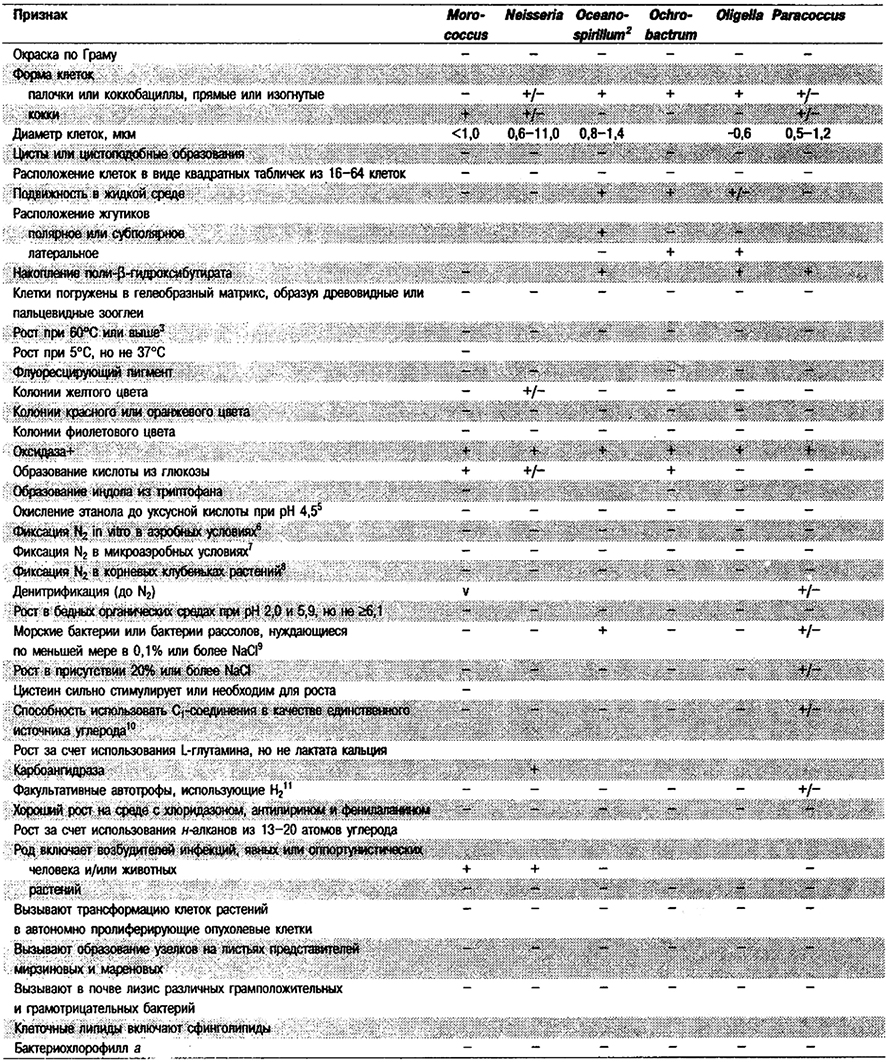
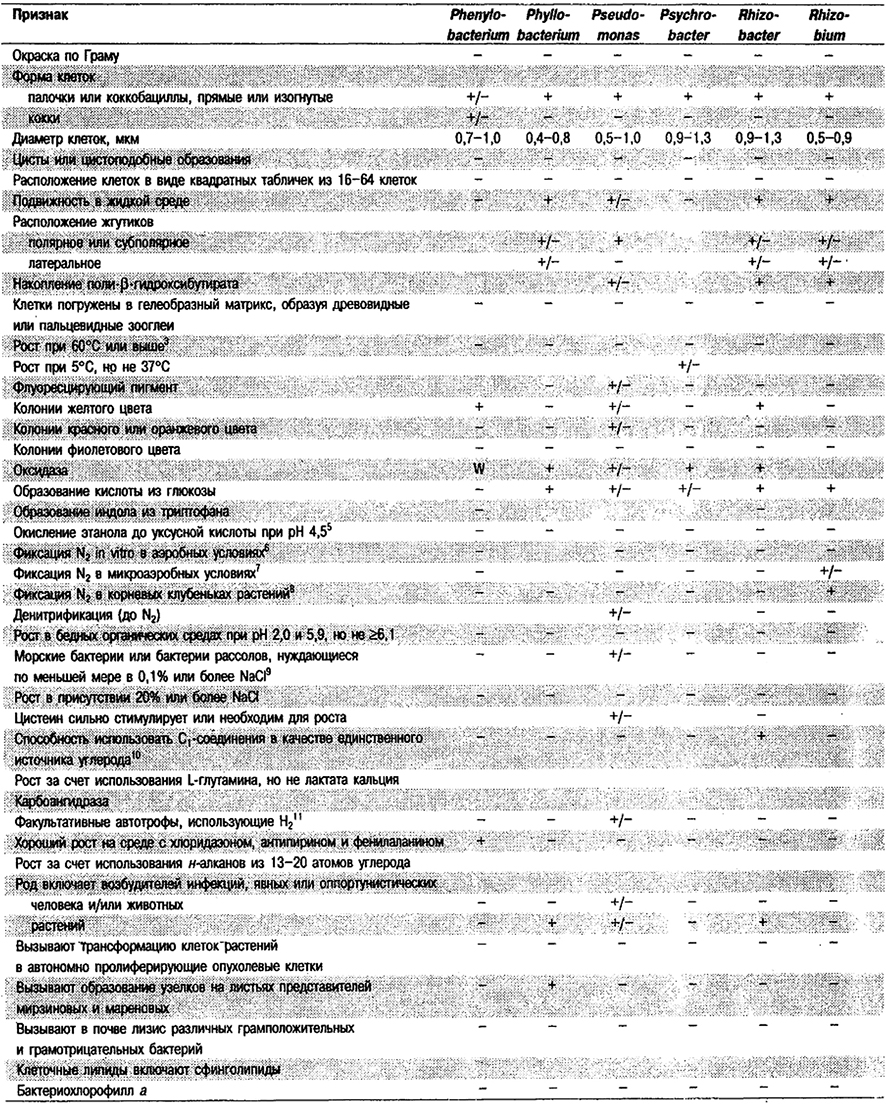
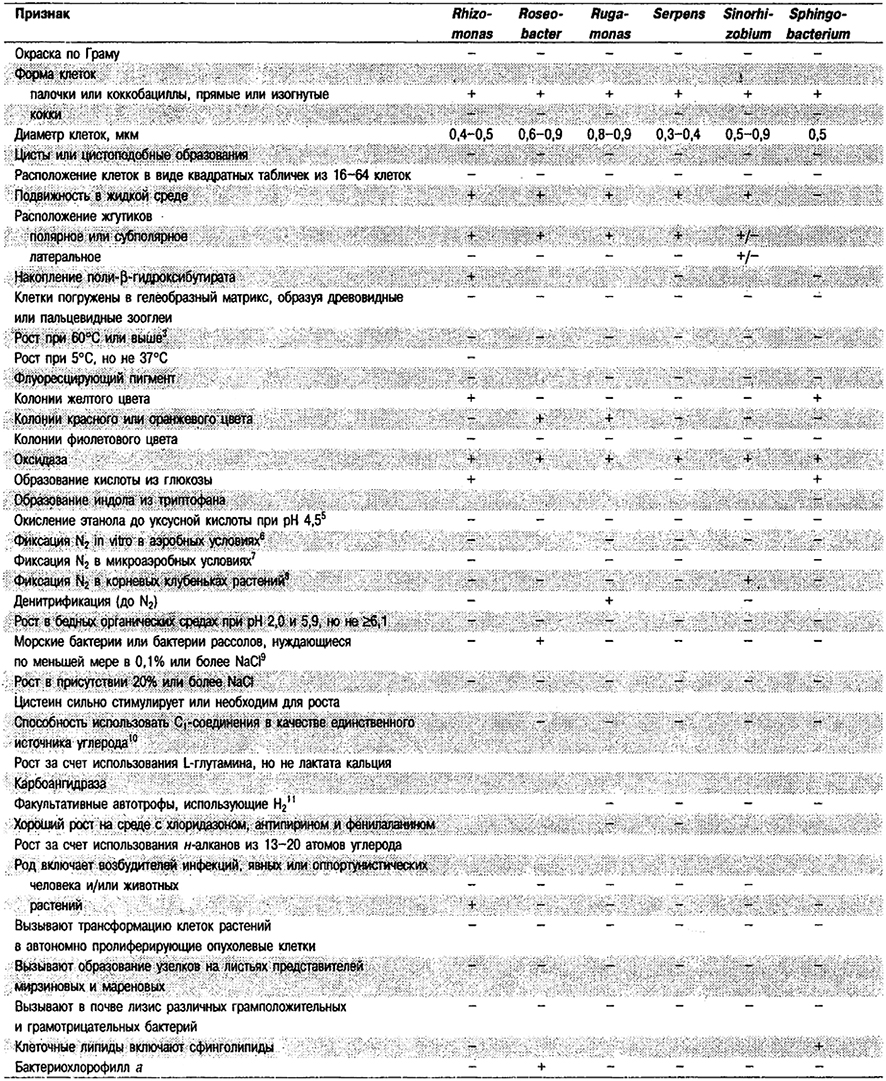
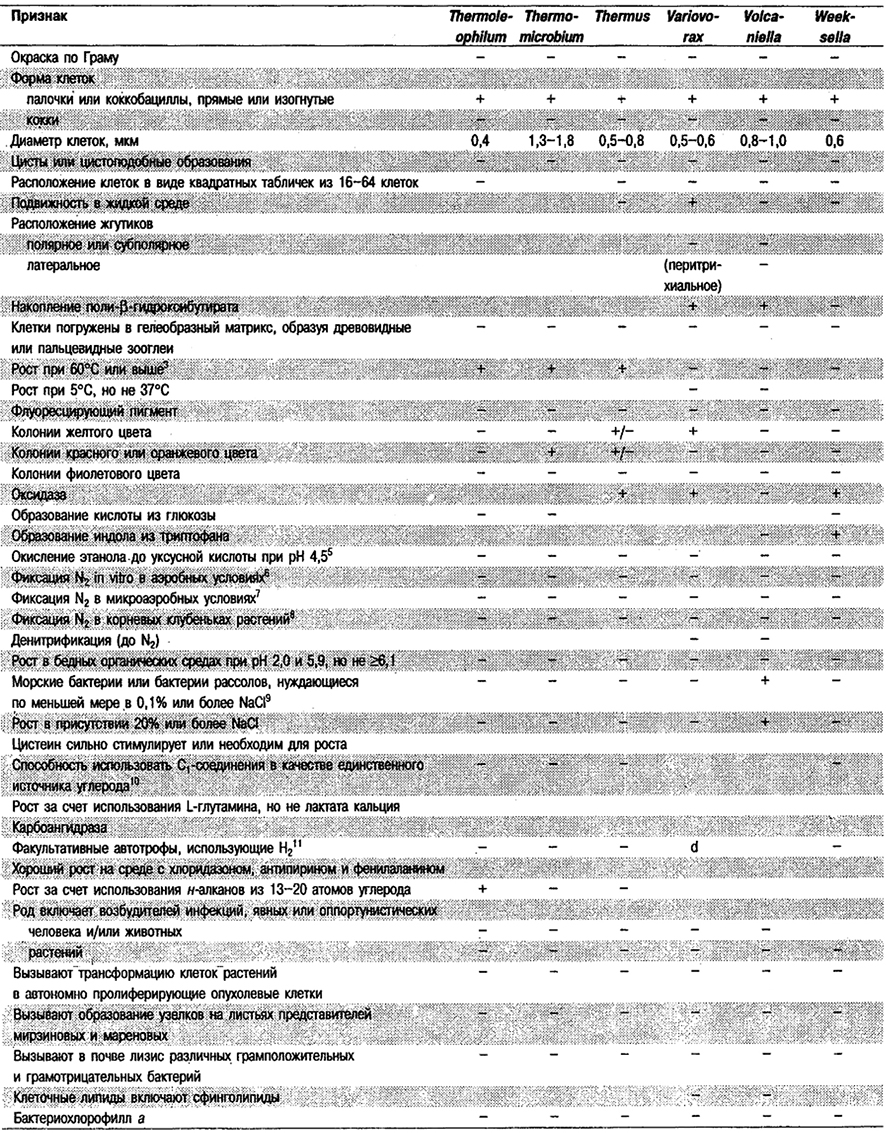
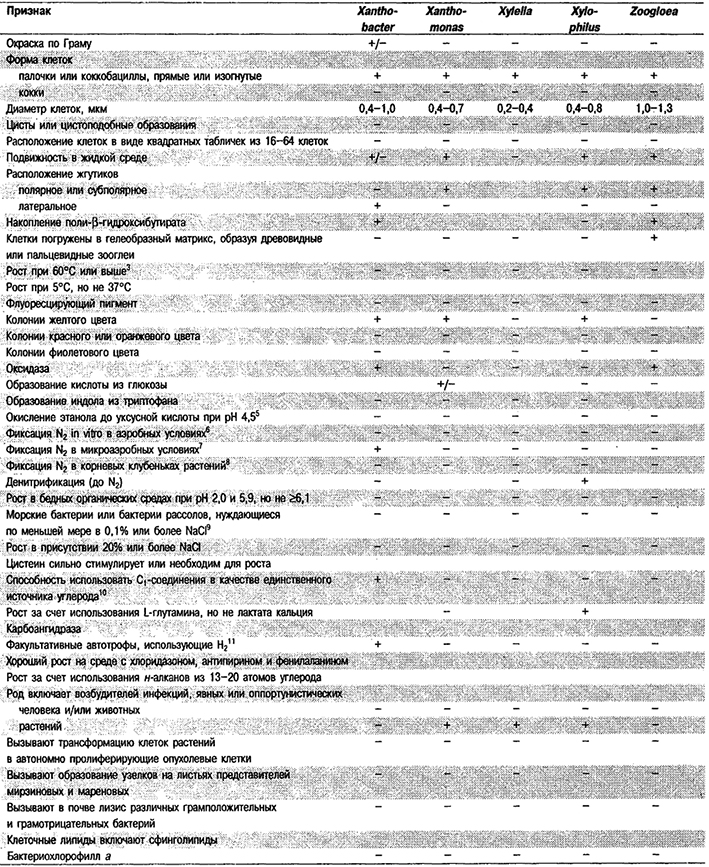

Таблица 4.2. Дифференциация грамотрицательных, аэробных,
гетеротрофных, палочковидных эубактерий, растущих при 60°С
или более высокой температуре1
Таблица 4.3. Дифференциация аэробных грамотрицательных бактерий,
окисляющих этанол до уксусной кислоты в нейтральных или кислых (рН 4,5) средах1 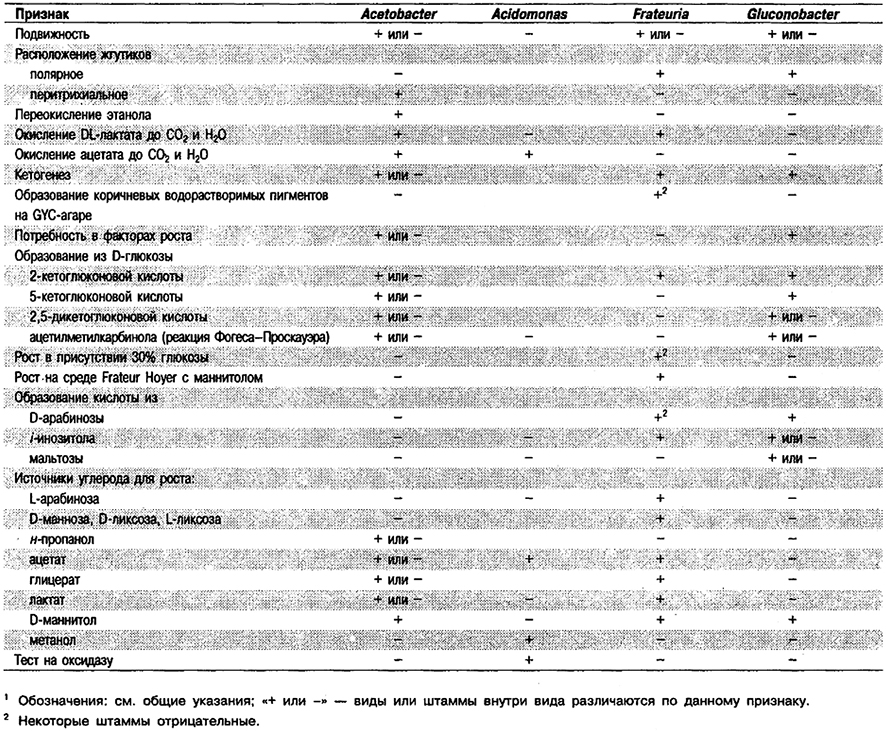
Таблица 4.4. Дифференциация грамотрицательных эубактерий,
фиксирующих N2 в аэробных условиях1 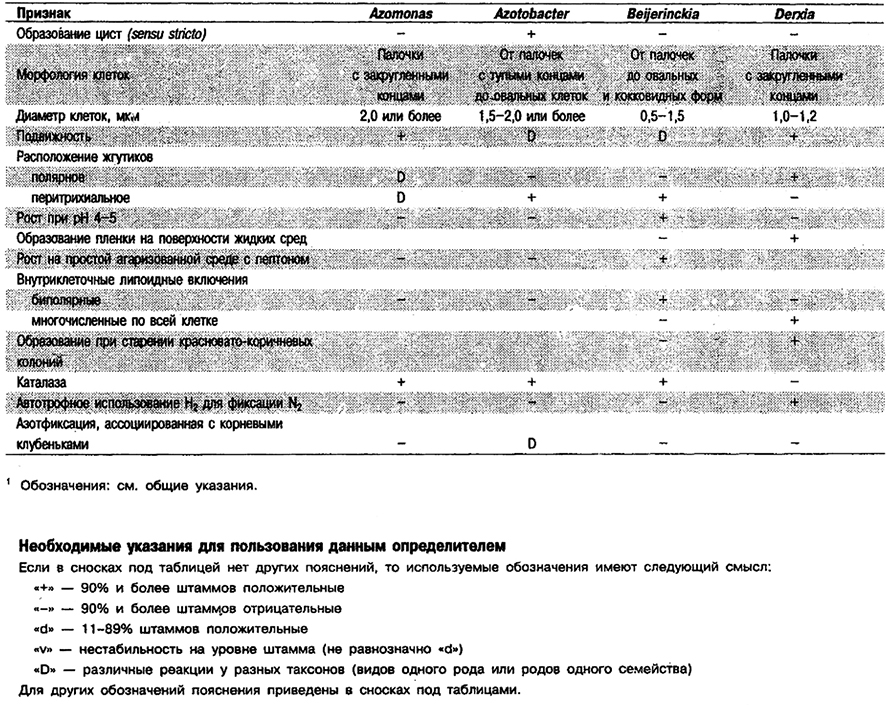
Таблица 4.5. Аэробные грамотрицательные палочковидные эубактврии,
фиксирующие N2 in vitro в микроаэробных условиях (например, в полужидких средах)
и не индуцирующие образование клубеньков на корнях или стеблях растений1 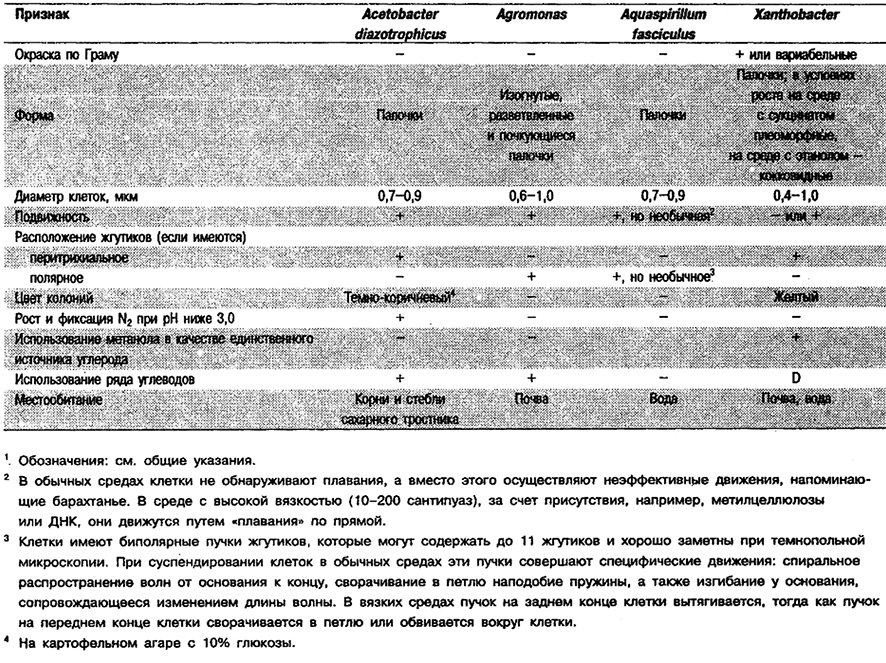
Таблица 4.6. Дифференциация родов азотфиксирующих,
грамотрицательных эубактернй, вызывающих образование клубеньков
на корнях и/или стеблях растений1 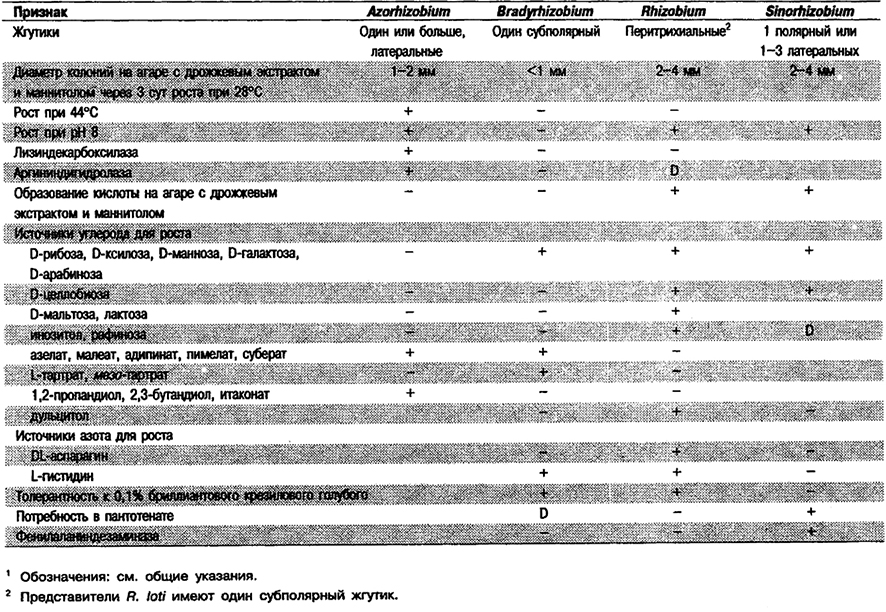
Таблица 4.7. Дифференцирующие признаки некоторых аэробных,
палочковидных морских бактерий, нуждающихся для роста в NaCI1 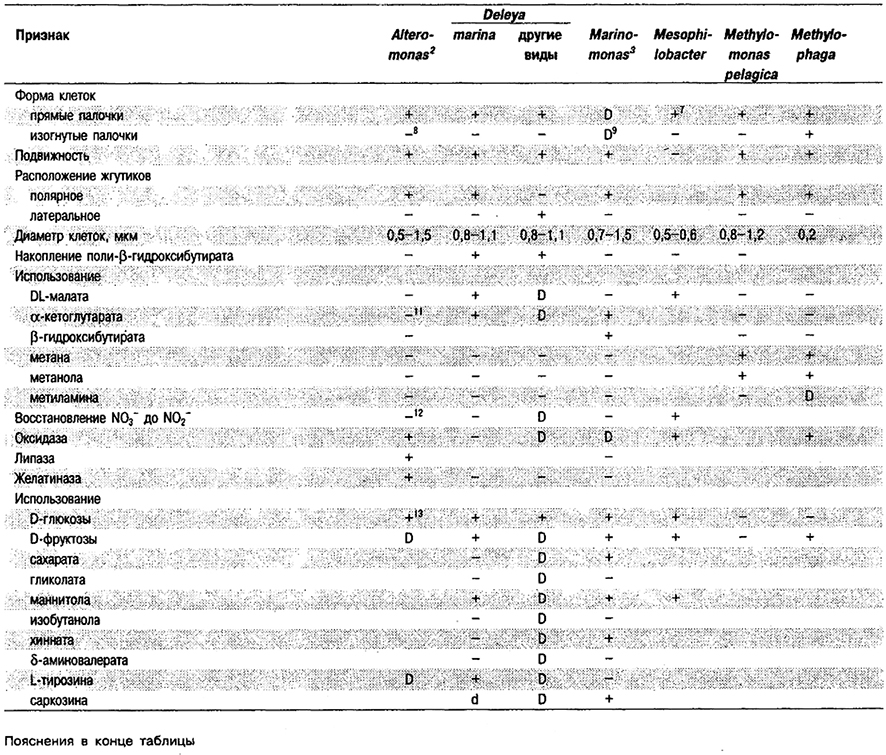
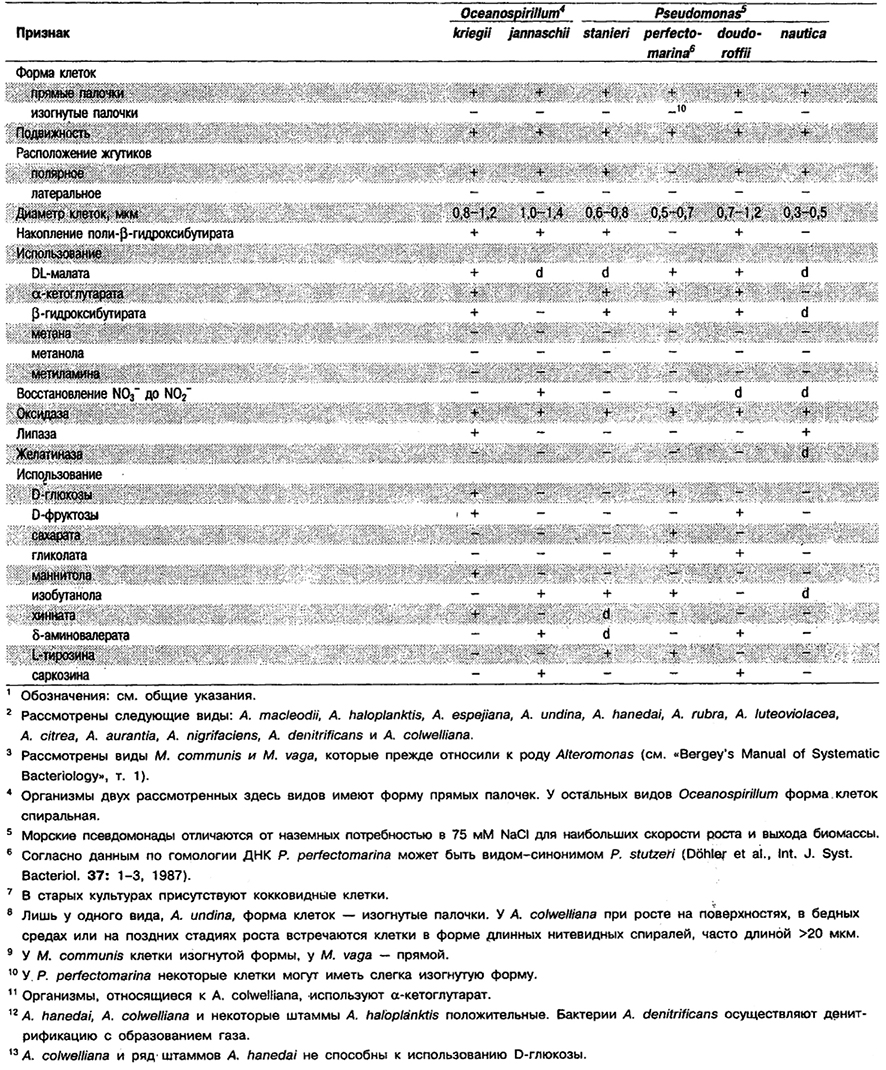
Таблица 4.8. Дифференцирующие признаки некоторых аэробных
грамотрицательных эубакгерий, способных использовать в качестве единственного источника
углерода одноуглеродные (С1–) соединения1 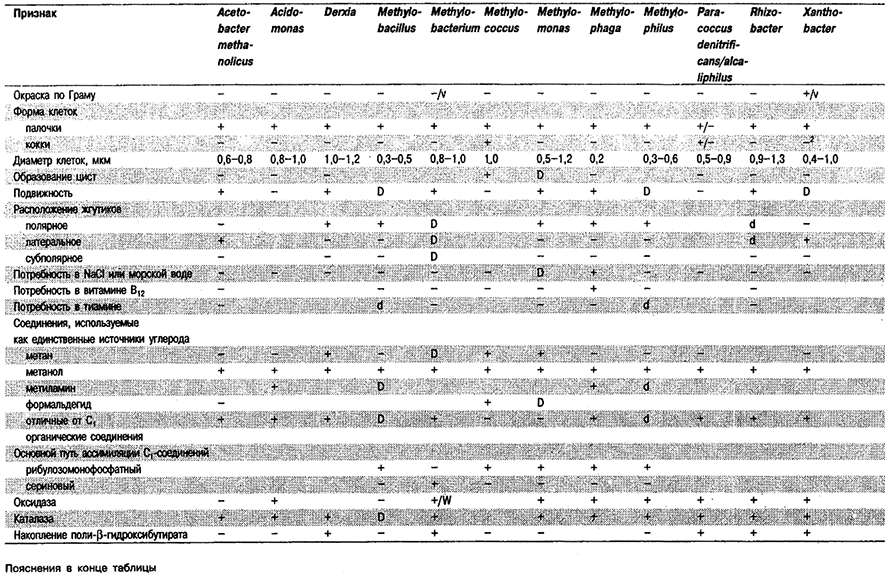
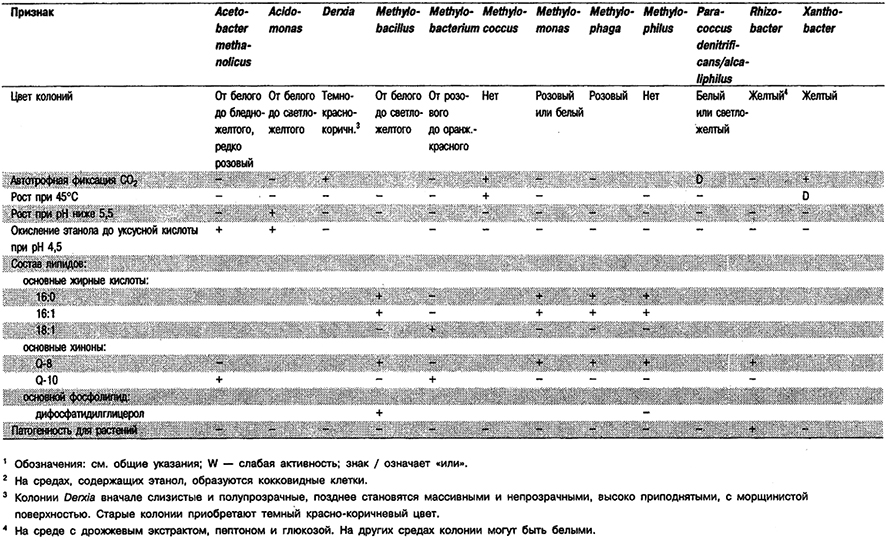
Таблица 4.9. Некоторые аэробные фамотрицательные эубакгерии –
факультативные водородокисляннцие автотрофы1 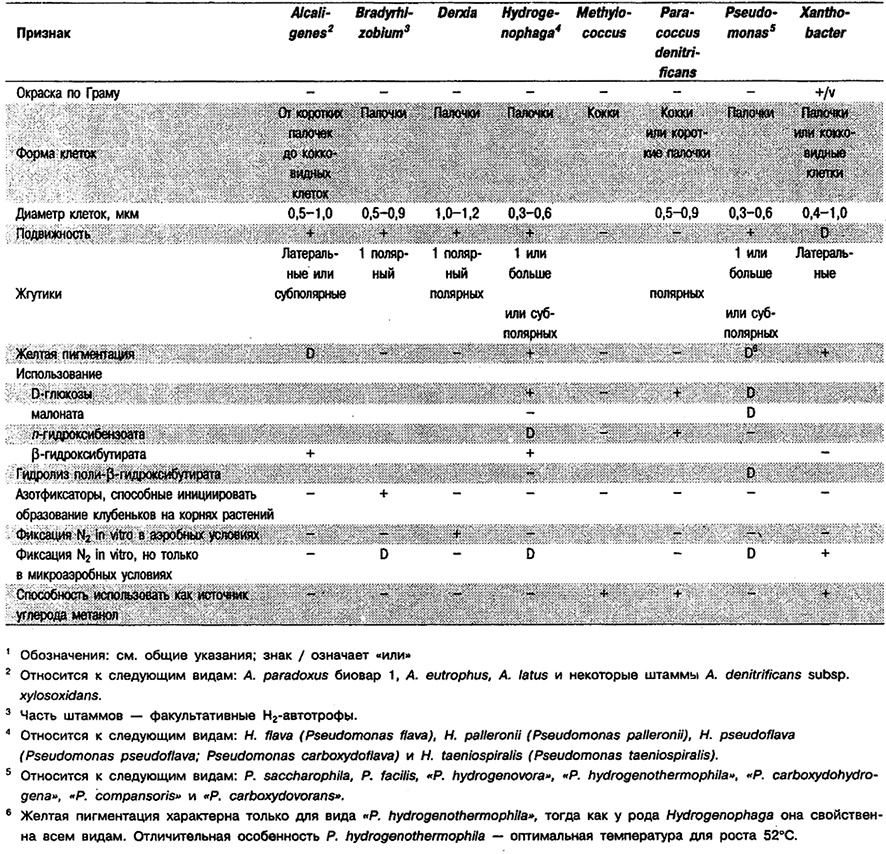
Род Acetobacter
Клетки от эллипсоидных до палочковидных, прямые или слегка изогнутые, 0,6–0,8 х 1,0–4,0 мкм, одиночные, в парах или цепочках. У некоторых штаммов часто присутствуют инволюционные формы, которые могут быть сферическими, удлиненными, раздутыми, булавовидными, изогнутыми или нитевидными. Клетки подвижные или неподвижные; у подвижных жгутики перитрихиальные или латеральные. Эндоспор не образуют. По Граму окрашиваются отрицательно (в редких случаях вариабельно). Облигатные аэробы; метаболизм дыхательного типа, никогда не бродильного. Колонии бледные, большинство штаммов пигменты не продуцирует. Для небольшого числа штаммов характерно образование водорастворимых пигментов или розовая окраска колоний за счет порфиринов. Каталазоположительные и оксидазо отрицательные. Желатину не разжижают, индол и H2S не образуют. Окисляют этанол до уксусной кислоты, ацетат и лактат – до СО2 и Н2О. Наилучшими источниками углерода для роста служат этанол, глицерол и лактат. Образуют кислоту из н-пропанола, н-бутанола и D-глюкозы. Лактозу и крахмал не гидролизуют. Хемоорганотрофы. Оптимальная температура 25–30°С. Оптимальный диапазон рН 5,4–6,3, Виды Acetobacter встречаются на цветах, фруктах, медоносных пчелах, в сакэ, спиртном напитке из сока агавы (производимом в Мексике), пальмовом и виноградном винах, сидре, пиве, южно-африканском пиве банту, кефире, пивных дрожжах, уксусе, буковых стружках из чанов-окислителен и чанах-окислителях, используемых в производстве уксуса, соке сахарного тростника, «чайном грибе», растительных дубильных растворах, «нате», садовой почве и воде каналов.
Некоторые представители рода Acetobacter могут вызывать розовую болезнь плодов ананасов и гниль яблок и груш. Бактерии, относящиеся к одному из видов, – микроаэрофилы и азотфиксаторы; они присутствуют на корнях и стеблях сахарного тростника.
Типовой вид: Acetobacter aceti.
Дифференциация видов рода Acetobacter: См. табл. 4.10.
Таблица 4.10. Дифференцирующие признаки видов рода Acetobacter1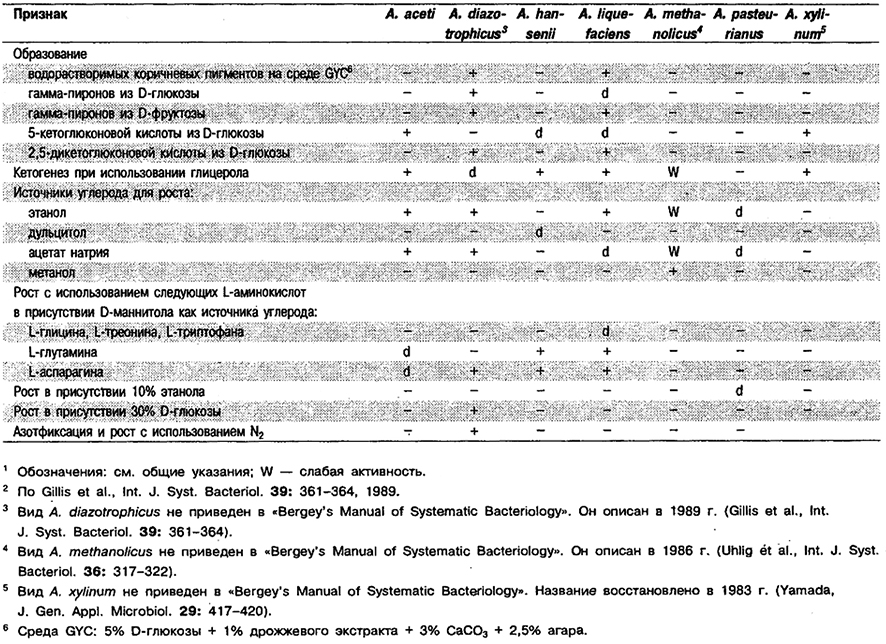
Род Acidiphilium
От редакции: Род Acidiphilium не приведен в «Веrgey`s Manual of Systematic Bacteriology». Этот род был описан в 1981 г. (Harrison, Int. J. Syst. Bacteriol. 31: 327-332) и первоначально включал единственный вид А, cryptum. Позднее были добавлены А. organovorum (Lobos et ah, Int i. Syst. Bacteriol. 36: 139-144, 1986), A. angustum, A. facilis и A. rubrum (Wtchlacz et art., Int, J. Syst. Bacteriol. 36: 197-201, 1986).
Прямые палочки с закругленными концами. Размеры варьируют у разных штаммов: диаметр от 0,3 до 1,2 мкм и длина от 0,6 до 4,2 мкм. Грамотри- цательные. Подвижные за счет единственного полярного или двух латеральных жгутиков; у некоторых штаммов клетки неподвижные. Эндоспор не образуют. Аэробы и ацидофилы; растут при рН от 2,5 до 5,9, но не при рН 6,1, Отдельные штаммы растут при рН 2,0. Мезофилы; хемоорганотрофы. В процессе выделения некоторые штаммы могут не расти при обычно используемых концентрациях пептона и экстрактов в среде; например, сухой триптиказо-соевый концентрат, добавленный в концентрации 0,05%, может ингибировать рост, а в концентрации 0,01% стимулировать его. Ацетат в качестве источника углерода не используют; это соединение может ингибировать рост. Легкоразлагаемым субстратом служит цитрат. Все штаммы растут за счет использования глюкозы, однако иногда требуется внесение в основную среду следовых количеств дрожжевого экстракта. Обнаружены в кислых минеральных природных средах: шахтных водах пиритовых рудников, отвалах угля с примесью пирита и отходах медных и урановых рудников. Виды Acidiphilium обнаружены также как спутники в накопительных культурах Thiobacillus ferrooxidans.
Типовой вид: Acidiphilium cryptum.
Дифференциация видов рода Acidiphilium: См. табл. 4.11.
Таблица 4.11. Дифференцирующие признаки видов рода Acidiphilium 1,2 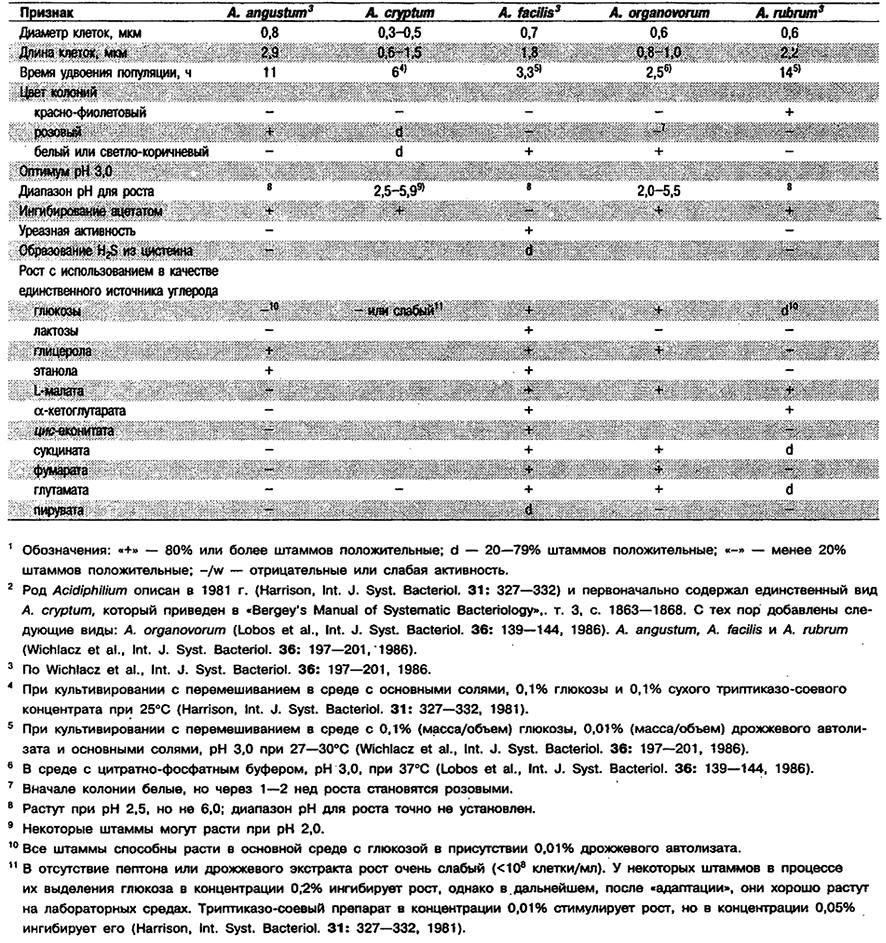
Род Acidomonas
От редакции: Род Acidomonas не приведен в «Sergey's Manual of Systematic Bacteriology». Этот род описан в 1989 г. (Urakami et al., fnt. J. Syst. Bacteriol. 39: 50-55) и включает единственный вид А. methanolica (прежнее название Acetobacter methanolica).
Грамотрицательные палочки с закругленными концами, 0,8-1,0 x 1,5-3,0 мкм, одиночные, редко в парах. Неподвижные. Колонии на агаризованной среде с пептоном и дрожжевым и солодовым экстрактами блестящие, гладкие, приподнятые, с цельным краем, от белых до светло-желтых, 2-3 мм в диаметре через 3 сут роста при 30°С. Водорастворимых флуоресцирующих пигментов не образуют. Аэробы; метаболизм чисто дыхательного, не бродильного, типа. Нитрат до нитрита не восстанавливают. Реакция Фогеса-Проскауэра отрицательная. Индол и H2S не образуют, Желатину и крахмал не гидролизуют. Образуют аммиак. Цвет лакмусового молочка не изменяют. Дигидроацетон из глицерола не образуют. Образуют уксусную кислоту из этанола. Образуют кислоту из D-глюкозы при ее метаболизировании по окислительному пути, но не из L-apaбинозы, D-ксилозы, D-маннозы, D-фруктозы, D-галактозы, мальтозы, сахарозы, лактозы, трегалозы, D-сорбитола, D-маннитола, инозитола, глицерола или растворимого крахмала. Используют в качестве единственного углеродного субстрата для получения энергии и роста метанол, этанол, уксусную кислоту, D-глюкозу, глицерол и пектин; не используют L-арабинозу, D-ксилозу, D-фруктозу, D-галактозу, мальтозу, сахарозу, лактозу, трегалозу, D-сорбитол, D-маннитол, ийозитол, растворимый крахмал, лимонную кислоту, молочную кислоту, метиламин, метан или молекулярный водород. Некоторые штаммы слабо используют D-маннозу. Для роста необходим пантотенат кальция. Источниками азота служат аммиак, нитрат и мочевина. Образуют уреазу, оксидазу и каталазу. Растут при рН 2,0–5,5, хорошо – при рН 3,0-5,0; при рН выше 6,0 или ниже 1,5 роста нет. Хорошо растут при 30 и 37°С, но не при 42°С. В присутствии 3% NaCl не растут. В составе клеточных жирных кислот преобладает не разветвленная ненасыщенная кислота С18:1, а также присутствуют в небольших количествах неразветвленные насыщенные кислоты C15:0, С16:0. С17:0, C18:0 и С19:0, неразветвленная ненасыщенная кислота С16:1 и циклопропановая кислота С19:0. В составе гидроксилированных кислот преобладают 3-ОН-кислоты C14:0 и C16:0, а также 2-ОН кислоты С14:0 и C16:0. Убихиноны представлены в основном Q-10, а также Q-9 и минорным Q-11.
Типовой и единственный вид: Acidomonas methanolica.
Признаки вида: Как для рода.
Род Acidothermus
От редакции: Род Acidothermus не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1986 г. (Mohagheghi et al., Int. J. Syst. Bacteriol. 36: 435-443) и включает единственный вид А. cellulolyticus.
Тонкие палочки и длинные тонкие нити, 0,4 x 5-20 мкм, с закругленными концами. Эндоспор, жгутиков и округлых тел не образуют. Неподвижные. Грамвариабельные, однако обычно окрашиваются по Граму отрицательно. На тонких срезах наружная мембрана в составе клеточной стенки не обнаружена; этим клеточные стенки Acidothermus отличаются от типичных грамотрицательных клеточных стенок бактерий видов Thermus и Thermomicrobium. Основные компоненты очищенных препаратов клеточных стенок – диаминопимелиновая кислота, глюкозамин, мурамовая кислота, серин и аланин. Колонии (выращенные на агаре LPBM с минеральными солями плюс 0,5 г дрожжевого экстракта и 5,0 г D-глюкозы или D-целлобиоэы на 1 л) сметанообраз- ные, белые, гладкие, округлые, с цельным краем, 1-3 мм в диаметре. Облигатные аэробы. Растут за счет использования нескольких углеводов, в том числе D-глюкозы и D-целлобиозы. Термофилы; растут в диапазоне температуры 37-70°С, Ацидофилы; растут при рН 3,5-7,0. Выделены из кислых горячих источников (рН 4-5,5, температура 45-65°С) на территории Йеллоустонского Национального Парка, США.
Типовой (и единственный) вид: Acidothermus cellulolyticus.
Признаки вида: Оптимальная температура для роста 50-60°С. Оптимальное значение рН-5,0. Источники углерода и энергии включают D-галактозу, D-целлобиозу, целлюлозу, ксилан, D-галактозу, мальтозу, сахарозу, рафинозу, D-маннозу, D-маннитол и D-сорбитол; источниками азота служат ионы аммония или аминокислоты. Растут на среде с казаминовыми кислотами (0,1%) и триптоном (0,1%); в МП Б, а также за счет использования ацетата, лактата, цитрата или пектина не растут. Цитрат и ацетат в концентрации 0,01 М ингибируют рост. Каталазоположительные. Активно разлагают целлюлозу.
Род Acidovorax
От редакции: Род Acidovorax не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1990 г. (Willems et al., Int. J. Syst. Bacteriol. 40: 384–398) и включает три вида: А. facilis (прежнее название – Pseudomonas facilis), А. delafieldii (прежнее название – Pseudomonas dela- fieldii) и A, temperans.
Палочки, от прямых до слегка изогнутых, 0,2–0,7 х 1,0–5,0 мкм, одиночные или в коротких цепочках. Подвижные за счет единственного полярного жгутика. Грамотрицательные. Оксидазоположительные. По активности уреазы штаммы различаются. Некоторые штаммы растут на агаре Кристенсена с мочевиной, однако по результатам тестов API 20NE уреазы не содержат. Пигмент при росте на питательном агаре не образуют. Аэробы и хемоорганотрофы. Acidovorax facilis и несколько штаммов Acidovorax delafieldii способны к литоавтотрофному росту с использованием Н2 в качестве источника энергии. Метаболизируют углеводы путем окисления с использованием О2 как конечного акцептора электронов; в то же время некоторые штаммы Acidovorax delafieldii и Acidovorax temperans способны восстанавливать нитрат, осуществляя гетеротрофную денитрификацию. Хорошо растут на средах с органическими кислотами, аминокислотами или пептоном, тогда как из сахаров рост обеспечивают лишь немногие. В состав жирных кислот всегда входят две гидроксилированные жирные кислоты – 3-гидроксиоктановая кислота (3-ОН-С8:0) и 3-гидроксидекановая кислота (3-ОН-С10:0); жирные кислоты, гидрокси- лированные по второму положению, отсутствуют; у большинства штаммов присутствует цикл о пропан овая жирная кислота (С17:цик). Средние величины содержания G+C в ДНК составляют 62-66 мол.%.
Типовой вид: Acidovorax facilis.
Дифференциация видов рода Acidovorax: См. табл. 4.11а.
Таблица 4,11а. Дифференцирующие признаки видов рода Acidovorax1 
Род Acinetobacter
Палочки диаметром 0,9–1,6 мкм и длиной 1,5–2,5 мкм в стационарной фазе роста становятся сферическими. Обычно в парах и цепочках различной длины. Спор не образуют. Грамотрицательные, однако иногда плохо обесцвечиваются. Движение типа плавания не осуществляют, но обнаруживают движение рывками («twitching motility»), предположительно обусловленное наличием полярных фимбрий. Аэробы; метаболизм чисто дыхательного типа с использованием кислорода в качестве конечного акцептора электронов. Все штаммы растут при температуре 20–З0°С; оптимальная температура для большинства штаммов 33–35°С. Хорошо растут на всех обычно используемых сложных средах. Оксидазоотрицательные и каталазоположительные. Большинство штаммов растет в средах определенного состава, содержащих один источник углерода и энергии; в качестве источника азота используют соли аммония или нитраты; в факторах роста не нуждаются. Некоторые штаммы из всех гексоз используют только D-глюкозу. Ряд штаммов использует также пентозы: D-рибозу, D-ксилозу и L-арабинозу. Встречаются в почве, воде и сточных водах.
Типовой вид: Acinetobacter calcoaceticus.
Дифференциация видов рода Acinetobacter: См. табл. 4.12.
Таблица 4.12. Дифференциация видов рода Acinetobacter 1,2 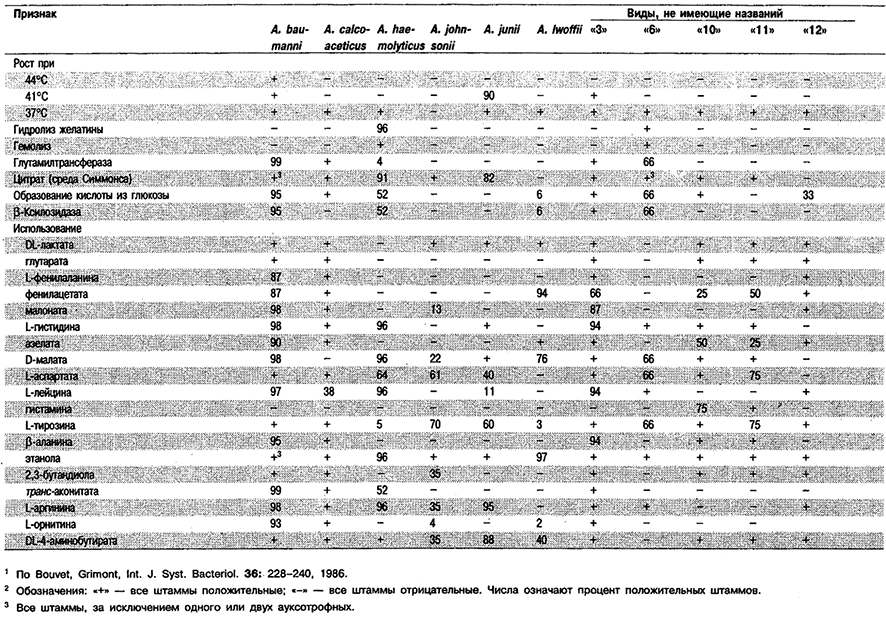
Род Afipia
От редакции: Род Afipia не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1991 г. (Brenneretal., J. Clin. Microbiol. 29: 2450-2460; Int. J. Syst. Bacteriol. 42: 327, 1992) и включает три вида; A. broomeae, A. clevelandensis и A. felts.
Клетки палочковидные, подвижные за счет единственного жгутика, расположенного полярно, субполярно или латерально. Грамотрицательные. Оксидазоположительные. Растут на забуференной агаризованной среде с обработанным активированным углем дрожжевым экстрактом (BCYE) и в МПБ, но не в МПБ с 6% NaCl и редко на агаре Макконки при 25 и 30°С. При 35°С обнаруживают по крайней мере слабый рост; при 42°С не растут. Колонии серо-белого цвета, блестящие, выпуклые, непрозрачные, с цельным краем, диаметром 1,5 мм; они появляются через 72 ч инкубации при 32°С на кровяном агаре или на агаре BCYE (на последнем размеры 0,5-1,5 мм). Уреазоположительные: лакмусовое молочко подщелачивают. Гемолиз, образование газа из нитрата, образование индола, образование H2S (метод посева на среду с тремя сахарами и железом, см. приложение), гидролиз желатины и эскулина и брожение не осуществляют. Кислоту при окислении D-глюкозы, лактозы, мальтозы или сахарозы не образуют. В составе клеточных жирных кислот преобладают разветвленная ω7-11-метил октадеценовая (С19:1), цис-ω7-октадеценовая (С18:1), а также обычно кислоты из 17 и 19 атомов углерода, содержащие циклопропановое кольцо; гидроксилированные кислоты присутствуют лишь в следовых количествах.
Ранее A. felis предположительно считали возбудителем болезни ог кошачьих царапин, однако, согласно последним данным, этот вид, возможно, не служит причиной заболевания. Другие виды считаются предположительно патогенными для человека.
Типовой вид: Afipia felis.
Дифференциация видов рода Afipia: См. дифференцирующие признаки трех указанных видов, а также трех не имеющих названий геновидов в оригинальной статье.
Род Agrobacterium
Палочки 0,6–1,0 x 1,5–3,0 мкм, одиночные или в парах. Спор не образуют. Грамотрицательные. Подвижные за счет перитрихиальных жгутиков, число которых варьирует от 1 до 6. Аэробы; метаболизм дыхательного типа с использованием кислорода в качестве конечного акцептора электронов. Некоторые штаммы могут осуществлять анаэробное дыхание в присутствии нитрата. Большинство штаммов способны расти в тканях растений при пониженном парциальном давлении кислорода. Оптимальная температура 25-28°С. Колонии обычно выпуклые, округлые, гладкие, от непигментированных до светло-бежевых. Рост на средах с углеводами, как правило, сопровождается обильным образованием внеклеточной полисахаридной слизи. Каталаэоположительные и обычно оксидазо- и уреазоположительные. Большинство штаммов, относящихся к A. tumefaciens биовар 1 и к A. radiobacter биовар I, образует 3-кетогликоэиды. Хемоорганотрофы; используют в качестве источников углерода разнообразные углеводы, соли органических кислот и аминокислоты, но не крахмал, целлюлозу и агар, содержащий глюкозу, D-галактозу и другие углеводы. Для штаммов некоторых видов и биоваров источниками азота могут служить соли аммония и нитраты, другим необходимы аминокислоты и дополнительные факторы роста. За исключением A. radiobacter представители этого рода внедряются через поврежденные покровы в ткани верхушек, корней и стеблей многочисленных двудольных, а также некоторых голосеменных растений и вызывают трансформацию растительных клеток в автономно пролиферирующие опухолевые клетки. Индуцируемые болезни известны как галл корончатый (бактериальный), корень волосяной и рак стеблей бактериальный. Для некоторых штаммов характерен широкий круг хозяев, тогда как для других (например, выделенных с виноградных лоз) очень ограниченный. Опухоли саморазмножаются и способны трансплантироваться. Индукция опухолей представителями Agrobacterium коррелирует с присутствием в клетках бактерий большой опухоленндуцирующей плазмиды (Ti-плазмнда). Местообитанием агробактерий служит почва. Онкогенные штаммы встречаются главным образом в почвах, куда ранее попадал пораженный растительный материал. Ряд неонкогенных штаммов выделен у человека из клинического материала.
Типовой вид: Agrobacterium tumefaciens.
Дифференциация видов рода Agrobacterium: См. табл. 4.13.
Таблица 4.13. Дифференцирующие признаки видов рода Agrobacterium 1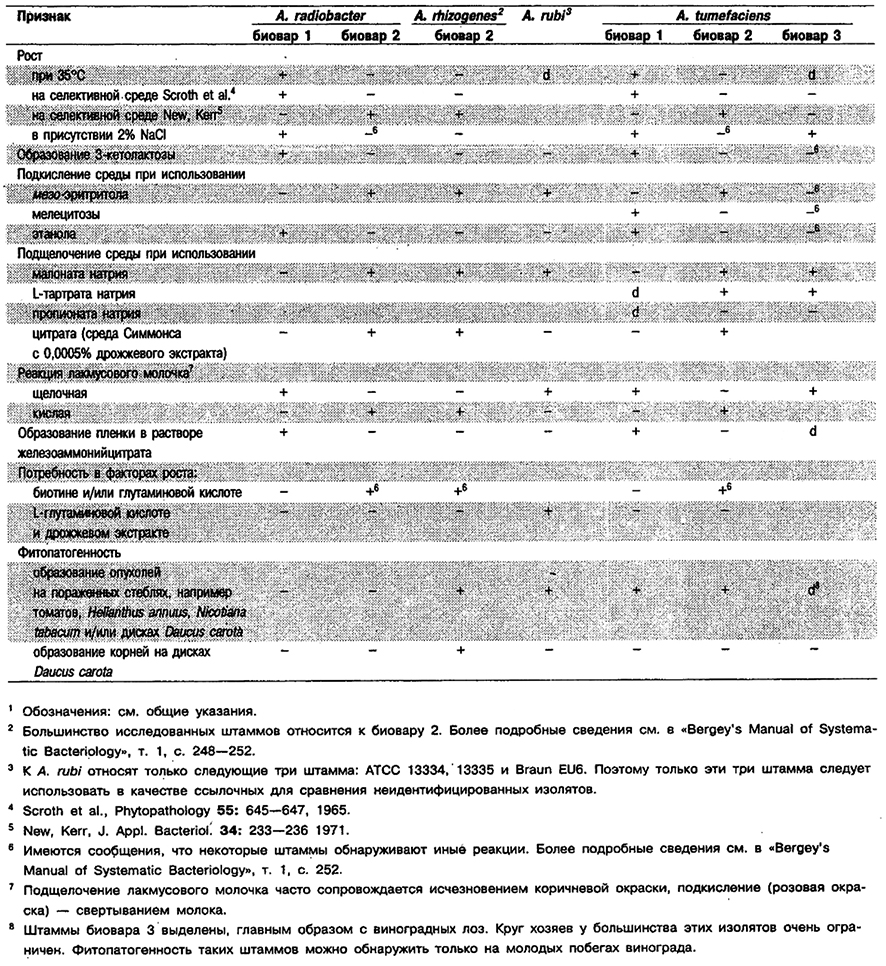
Род Agromonas
От редакции: Род Agromonas не приведен в «Вегgey's Manual of Systematic Bacteriology». Этот род описан в 1983 г. (Ohta, Hattori, Antonie van Leeuwenhoek, J. Microbiol. Serol. 49: 429-446; Int. J. Syst. Bactenol. 35: 223, 1985) и включает единственный вид А. oligotrophia.
Клетки изогнутые, разветвленные и почкующиеся, 0,6–1,0 x 2–7 мкм, подвижные за счет полярных жгутиков. Покоящиеся стадии неизвестны. Грамотрицательные. Колонии бесцветные. Каталазо- и оксидазоположительные. Казеин, желатину, крахмал и целлюлозу не гидролизуют. При низком парциальном давлении О2 фиксируют атмосферный азот. В составе жирных кислот преобладает неразветвленная ненасыщенная кислота C18:1. Содержат убихинон Q-10.
Типовой (и единственный) вид: Agromonas oligotrophia.
Признаки вида: В разбавленной жидкой питательной среде клетки изогнутые, разветвленные и почкующиеся, соединенные по нескольку вместе в виде розеток, подвижные за счет одного полярного жгутика. Эндоспор не образуют. Колонии на разбавленной агаризованной питательной среде точечные, подушечкообразные, с цельным краем, бесцветные. Олигогрофы, т. е. могут расти в среде, содержащей менее 1 мг органического углеродного субстрата в 1 л. NaCl, КСl, казамнновые кислоты, пептон и мясной экстракт в концентрации 0,5–1,0% ингибируют рост. Используют несколько сахаров и многие органические кислоты. Используют ароматические кислоты, такие как феруловая, n-кумаровая и n-анисовая, но не бензойную. Из глюкозы не образуют ни кислоты, ни газа. Протеолитичес кой активностью (тесты с желатиной и казеином) не обладают. Целлюлозу и крахмал не гидролизуют, Аэробы. В составе клеточных жирных кислот преобладает С18:1, в качестве минорных компонентов обнаружены С16:0, а также ненасыщенная кислота С19 с одной двойной связью и, возможно, одной боковой цепью. Содержат убихинон.
Род Aicaligenes
Палочки, кокко видные палочки или кокки, 0,5–1,0 x 0,5–2,6 мкм, как правило одиночные. Покоящиеся стадии неизвестны. По Граму окрашиваются отрицательно. Подвижные за счет 1-8 (в отдельных случаях до 12) перитрихи альных жгутиков. Облигатные аэробы; метаболизм чисто дыхательного типа с использованием кислорода в качестве конечного акцептора электронов. Некоторые штаммы способны к анаэробному дыханию в присутствии нитрата или нитрита. Оптимальная температура 20–37°С. Колонии на МПА неокрашенные. Оксидазо- и каталазоположнтельные. Индол не образуют. Целлюлозу, эскулин, желатину и ДНК, как правило, не гидролизуют. Хемоорганотрофы; используют в качестве источников углерода разнообразные органические кислоты и аминокислоты. При использовании ряда оргаькических солей и амидов образуют щелочь. Углеводы обычно не используют. Некоторые штаммы образуют кислоту из D-глюкозы и D-ксилозы и используют оба углевода как источник углерода. Встречаются в воде и почве. Некоторые представители относятся к обычным, по-видимому сапрофитным, обитателям кишечного тракта позвоночных. Многочисленные штаммы вьщелены из клинического материала, такого как кровь, моча, фекалии, гнойные ушные выделения, спинномозговая жидкость, отделяемое из ран и т. д. Иногда вызывают оппортунистические инфекции у человека.
Типовой вид: Alcaligenes faecal is.
Дифференциация видов рода Alcaligenes: См. табл. 4.14.
Таблица 4.14. Дифференцирующие признаки видов рода Alcaligenes 1,2 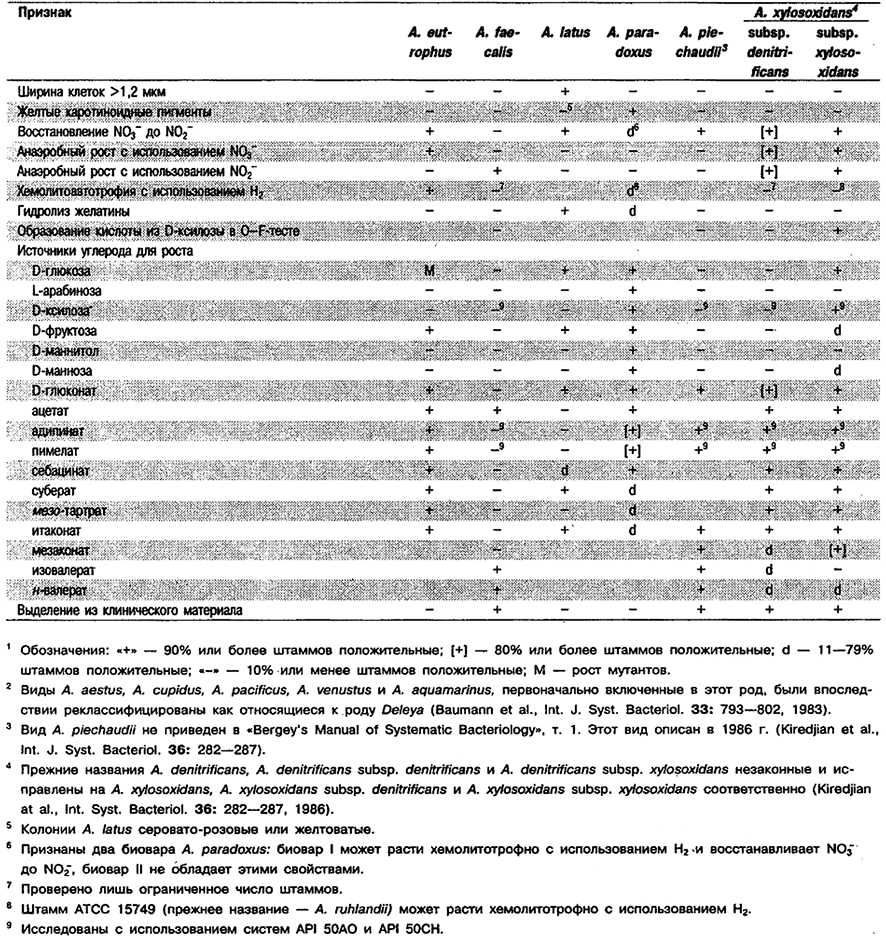
Род Alteromonas
Прямые или изогнутые палочки, 0,7-1,5 x 1,8-3,0 мкм. Поли-β-гидрокеибутират в качестве запасного продукта не накапливают. Микроцисты и эндоспоры не образуют. Грамотрицательные. Подвижные за счет единственного полярного, не окруженного чехлом жгутика; у двух видов (A. luteoviolacea и A. denitrificans) жгутики окружены чехлом. Хемоорганотрофы; метаболизм дыхательного, не бродильного, типа. Универсальным акцептором электронов служит молекулярный кислород. Один вид способен к денитрификацин. Ни один из штаммов не обладает конститутивной аргининдигидролазной системой. Все штаммы нуждаются для роста в морской воде как основе среды, многие также – в органических факторах роста. Все растут при 20°С. Как правило, присутствуют в морской воде в прибрежной зоне и открытом океане.
Типовой вид: Alteromonas macleodii.
Дифференциация видов рода Alteromonas: См, табл. 4.15.
Таблица 4.15. Дифференцирующие признаки видов рода Alteromonas 1,2 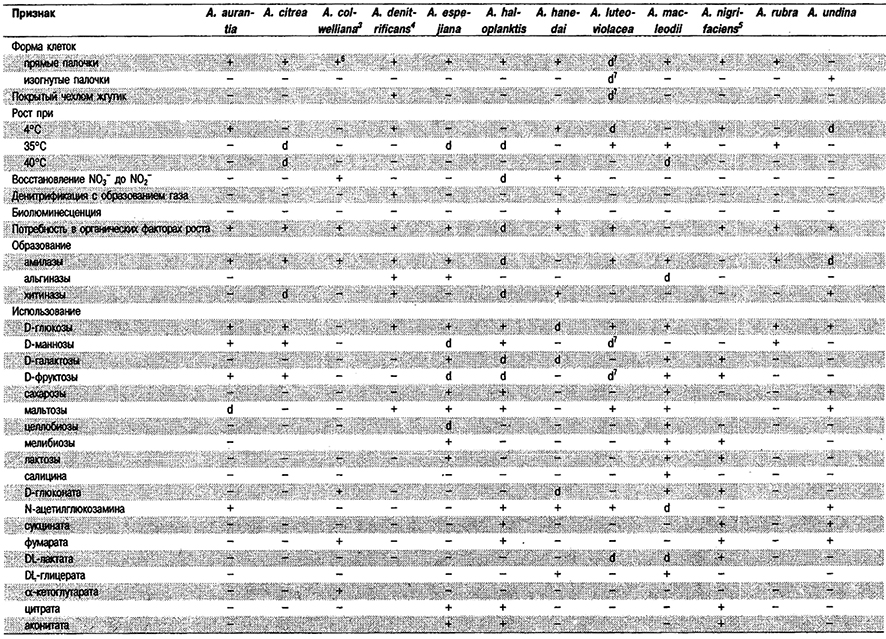
Род Aminobacter
От редакции: Род Aminobacter не приведен в «Вегgey's Manual of Systematic Bacteriology». Этот род описан в 1992 г. (Urakami et al., Int. J. Syst. Bactenol. 42: 84–92) и включает три вида: A. aminovorans (прежнее название – Pseudomonas aminovorans), A. aganoensis и A, niigataensis.
Палочки с закругленными концами, 0,5–0,9 х 1,0–3,0 мкм. Грамотрицательные. Подвижные за счет субполярных жгутиков. Размножаются почкованием. Накапливают в клетках гранулы поли-β-гидроксибутирата. Колонии от белых до светло-желтых. Хорошо растут в МПБ и жидкой среде с пептоном, дрожжевым гидролизатом и глюкозой. Водорастворимых флуоресцирующих пигментов не образуют. Проба с метиловым красным и реакция Фогеса-Проскауэра отрицательные. Индол и H2S не образуют. Желатину и крахмал не гидролизуют. Образуют аммиак. Денитрификацию не осуществляют. Образуют кислоты, из сахаров в результате их окисления, но не сбраживания. Используют монометиламии, триметиламин, триметиламин-N-оксид и сахара, но не метанол, метан и молекулярный водород. В факторах роста как существенных компонентах среды не нуждаются. В качестве источников азота используют аммиак, нитрат, мочевину, пептон и метиламин. Оксидазо- и каталазоположительные, уреазоотрицательные. Аэробы. Метаболизм чисто дыхательного, не бродильного, типа. Хорошо растут в диапазоне рН 6,0–8,0, но не при рН выше 9,0 и ниже 5,0. Хороший рост наблюдается при 30 и 37°С, но не при 42°С. В присутствии 3% NaCl не растут.
Клеточные жирные кислоты включают в большом количестве неразветвленную ненасыщенную кислоту С18:1, а гнлроксилированные кислоты – большое количество 3-ОН-кислотьг С12:0. Основной убихинон – Q-10.
Типовой вид: Aminobacter aminovorans.
Дифференциация видов рода Aminobacter: См. табл. 4.15а.
Таблица 4.15а. Дифференцирующие признаки видов рода Aminobacter 1 
Род Aquaspirillum: A. fasciculus
Прямые палочки, 0,7–0,9 x 3,6–43,0 мкм. У одного штамма после многократных пересевов отмечены варианты S-образной или изогнутой формы. Присутствуют внутриклеточные гранулы поли-β-гндроксибутирата. В старых культурах происходит массовое превращение клеток из палочковидных в округлые «кокковидные тела». Клетки из культур в жидкой среде имеют биполярные пучки жгутиков, числом до 11 в каждом пучке. Эти пучки, четко видимые при микроскопировании в темном поле, обнаруживают необычное и характерное поведение, когда клетки суспендированы в обычной невязкой среде: распространение спиральных волн от основания к верхушке, способность сворачиваться в кольца наподобие пружины и изгибаться у основания с изменением длины волны. В обычной среде клетки не плавают, а производят неэффективные движения, напоминающие «барахтанье». Если клетки, выращенные в жидкой среде, суспендировать в среде с высокой вязкостью (10–200 сантипуаз), получаемой с помощью таких соединений, как метшшеллюлоза «400 сантипуаз», клетки плавают, двигаясь по прямой. На начальных стадиях выделения клетки образуют очень вязкие хлопья, при последующих пересевах способность к образованию хлопьев постепенно утрачивается. Если хлопья разрушить и гомогенизировать в небольшом количестве воды, то в них видны свободно плавающие клетки, двигающиеся по прямой; в этом случае хвостовой пучок жгутиков тянется позади каждой клетки, а пучок на переднем конце клетки либо закручивается в петлю, либо обвивается вокруг клетки. Оптимальная температура 30°С; при 20 или 40°С не растут, Каталазо- и оксидазоположительные. Углеводы не катаболизируют. В качестве единственного источника углерода наиболее эффективны пируват и пролин. В микроаэробных условиях проявляют ннтрогеназную активность. Местообитание – прудовая вода.
Род Azomonas
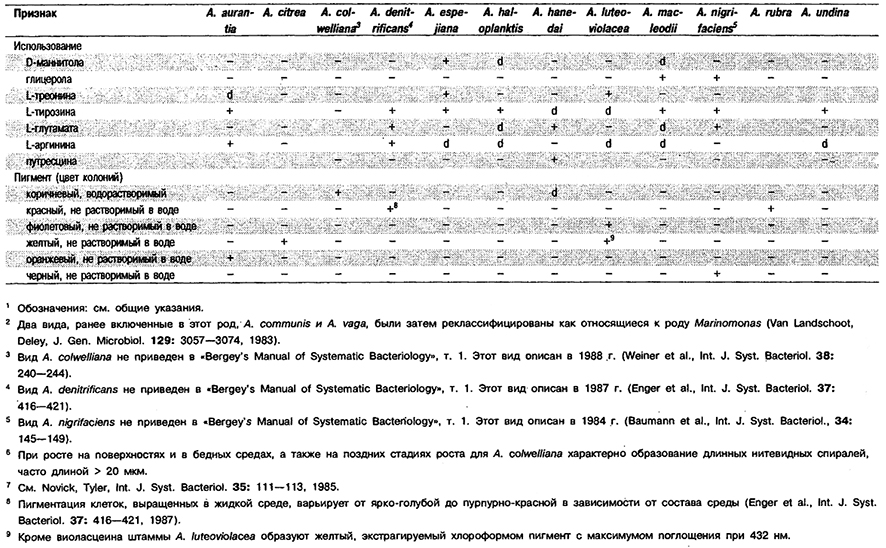
Клетки от палочковидных до овальных и кокковидных, диаметром 2 мкм и различной длины, одиночные, в парах или группах. Обычно наблюдается плеоморфизм. Как правило, грамотрицательные, иногда грамвариабельные. Эндоспор или цист не образуют. Наблюдается подвижность за счет перитрихиальных или полярных жгутиков. Аэробы, но могут также расти и при пониженном парциальном давлении кислорода. Почти все штаммы образуют водорастворимые пигменты и флуоресцирующие пигменты. Хемоорганотрофы; используют в качестве субстратов для роста сахара, спирты и соли органических кислот. Азотфиксаторы; обычно фиксируют в несимбиотическом состоянии не менее 10 мг N2 в расчете на 1 г потребленного углевода (как правило, глюкозы). Для азотфиксацин нуждаются в молибдене. Белки не гидролизуют; в качестве источника азота могут использовать соли аммония и некоторые аминокислоты. Катал азоположительные. Оптимальное значение рН для азотфиксации близко к нейтральному, однако некоторые штаммы могут также фиксировать азот и при рН 4,6–4,8. Встречаются в почве и воде.
Типовой вид: Azomonas agilis.
Дифференциация видов рода Azomonas: См. табл. 4.16.
Таблица 4.16. Дифференцирующие признаки видов рода Azomonas1 
Род Azorhizobium
От редакции; Род Azorhizobium не приведен в «Berge`s Manual of Systematic Bacteriology». Этот род описан в 1988 г. (Dreyfus et al., Int. J. Syst. Bacteriol. 38: 89-98).
Палочки, 0,5-0,6 x 1,5-2,5 мкм. Подвижные; при росте на плотной среде имеют перитркхиальные жгутики, а в жидкой среде – один латеральный. Колонии на агаризованной среде округлые, кремового цвета. Облигатные аэробы. В микроаэробных условиях фиксируют N2 и хорошо растут за счет использования N2 в среде без азота в присутствии витаминов. Оксидазо- и каталазоположнтельные. Уреазоотрицательные. Из Сахаров окисляют только глюкозу. Как при NH4+-, так и при N2-зависимом росте в качестве углеродных субстратов предпочтительны органические кислоты, такие как молочная или янтарная. Используют также малонат. Крахмал не гидролизуют. Могут расти за счет использования DL-пролина. Ни один из штаммов не восстанавливает нитрат. Штаммы эффективно образуют клубеньки на корнях и стеблях Sesbania rostrata.
Типовой (и единственный) вид: Azorhizobium caulinodans.
Признаки вида: Обладают аргининдигидролазой и лизиндекарбоксилазой. Растут в присутствии 8% KNО3. Растут за счет использования азелата, малеината, адипината, пимелата, суберата, глкжоната, муката, кротоната, никотината, 2-кетоглюконата, пропионата, бутирата, изобутирата, валерата, изовалерата, капроната, лаурата, 2-кетоглугарата, фумарата, глутарата, себацината, DL-малата, цитрата, пирувата, аконитата, цитраконитата, D-глюкуроната, α-D-галактуроната, м-гидроксибензоата, L-аспартата, хинната, L-аланина, L-лизина, L-аспарагина, бетаина и саркозина. Маннитол не окисляют. Ассимилируют 1,2-пропандиол и 2,3-бутандиол. Диапазон температуры для роста 12-43°С, Из витаминов нуждаются в никотиновой кислоте для фиксации N2 в микроаэробных условиях. Одинаково хорошо растут при рН 5,5 и 7,8.
Род Azotobacter
Крупные клетка овальной формы, диаметром 1,5–2,0 мкм, плеоморфные, от палочковидных до кокковидкых. Располагаются одиночно, парами или группами неправильной формы, иногда в виде цепочек разной длины. Эндоспор не образуют, но образуют цисты. Грамотрицательные. Подвижные за счет перитрихиальных жгутиков или неподвижные. Аэробы, но могут также расти при пониженном парциальном давлении кислорода, У всех видов некоторые штаммы продуцируют водорастворимые и водонерастворимые пигменты. Хемоорганотрофы; используют для роста сахара, спирты и соли органических кислот. Азотфиксаторы; обычно в несимбиотическом состоянии фиксируют не менее 10 мг N2 в расчете на 1 г потребленного углевода (как правило, глюкозы). Для азотфиксацин нуждаются в молибдене, который; однако, может быть частично заменен ванадием. Белки не гидролизуют. В качестве источников азота используют нитрат и (за исключением одного вида) соли аммония, а также некоторые аминокислоты. Каталазоположительные. Диапазон рН для роста в присутствии источников связанного азота 4,8-8,5; оптимальный диапазон рН для роста и азотфиксации 7,0-7,5. Встречаются в почве и воде, один вид – в ассоциации с корнями растений.
Типовой вид: Azotobacter chroococcum.
Дифференциация видов рода Azotobacter: См. Табл. 4.17.
Таблица 4.17. Дифференцирующие признаки видов рода Azotobacter 1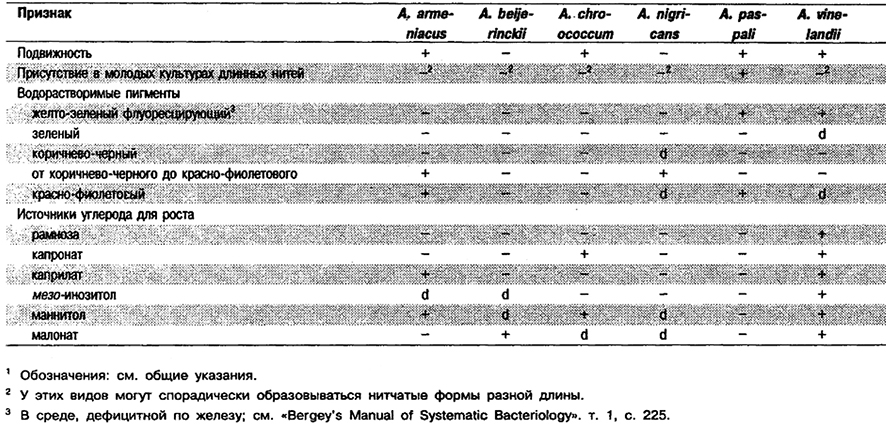
Род Beijerinckia
Прямые или слегка изогнутые палочки, 0,5–1,5 х 1,7–4,5 мкм, с закругленными концами. Располагаются одиночно. Иногда встречаются крупные, 3,0 x 5,0–6,0 мкм, бесформенные клетки, в отдельных случаях разветвленные или вильчатые. Могут содержать крупные, сильно преломляющие свет внутриклеточные гранулы поли-β-гидроксибутирата, обычно по одной у каждого полюса. Некоторые виды способны образовывать цисты (окружающие одну клетку) и капсулы (окружающие несколько клеток). Грамотрицательные. Подвижные за счет перитрихнальных жгутиков или неподвижные. Аэробы; метаболизм чисто дыхательного типа с использованием кислорода в качестве конечного акцептора электронов. Фиксируют молекулярный азот в аэробных условиях, а также при пониженном содержании кислорода (в микроаэробных условиях). Оптимальная температура 20–30°С, при 37°С не растут». Рост происходит в диапазоне рН от 3,0 до 9,5–10,0. При росте в жидкой среде поверхностной пленки не образуют, но всю среду превращают в гомогенную, очень вязкую, полупрозрачную массу; у некоторых видов в процессе роста среда становится опалесцирующей и мутной, а клейкая слизь не образуется. При росте на агаризованной среде, особенно в условиях азотфиксации, выделяют обильную, прочную, эластичную слизь и образуют гигантские колонии с гладкой, складчатой или «плиссированной» поверхностью; для некоторых штаммов характерна более зернистая консистенция слизи, подобная таковой у Azotobacter. Каталазоположительные. Все штаммы нспользуют глюкозу, фруктозу и сахарозу с образованием СО2 и небольшого количества уксусной кислоты. На пептонном агаре или в жидкой среде с пептоном не растут. Глутамат используют слабо или не используют. Встречаются в почвах, особенно часто в тропическом поясе.
Типовой вид: Beijerinckia indica.
Дифференциация видов рода Beijerinckia: См. табл. 4.18.
Таблица 4.18. Дифференцирующие признаки видов рода Beijerinckia 1 
Род Bordetella
Мелкие коккобациллы, 0,2–0,5 х 0,5–2,0 мкм, часто биполярно окрашенные, одиночные или в парах, реже в цепочках. Грамотрицательные, Могут быть подвижными или неподвижными; подвижность, если есть, обусловлена перитрихиальными жгутиками. Облигатные аэробы. Оптимальная температура 35–37°С, Колонии на среде Бордэ-Жангу гладкие, выпуклые, жемчужные, блестящие, почти прозрачные и окруженные зоной гемолиза без выраженной границы. Метаболизм дыхательного типа, никогда не бродильного. Хемоорганотрофы, нуждаются в никотииамиде, органических соединениях серы (например, цистеине) и органических азотистых соединениях (аминокислотах). Окисляют глутаминовую кислоту, пролин, аланин, аспарагиновую кислоту и серин с образованием аммиака и СО2. Лакмусовое молочко подщелачивают. Паразиты млекопитающих; патогенные. Локализуются и размножаются среди ресничек эпителия дыхательного тракта.
Типовой вид: Bordetella pertussis.
Дифференциация видов рода Bordetella: См. табл. 4.19.
Таблица 4.19. Дифференцирующие признаки видов рода Bordetella 1 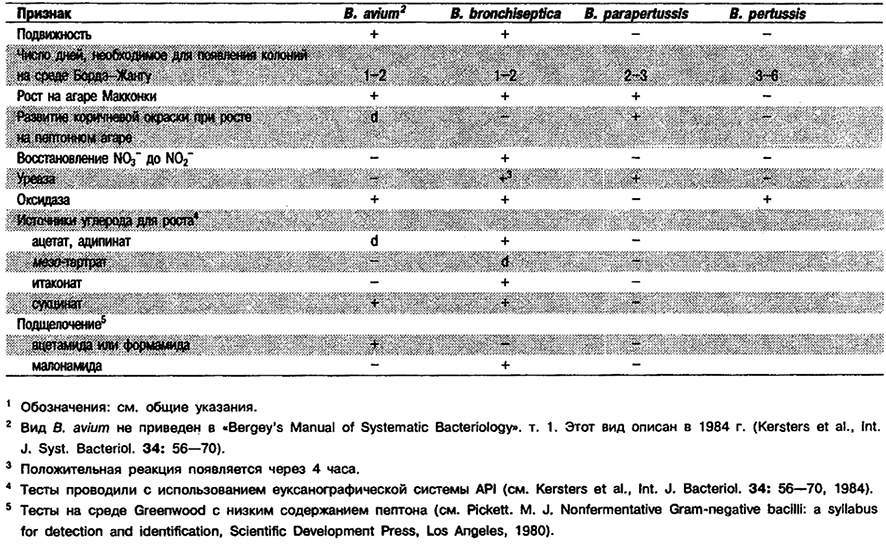
Род Bradyrhizobium
Палочки, 0,5-0,9 x 1,2-3,0 мкм. В неблагоприятных для роста условиях, как правило, плеоморфные; обычно содержат гранулы поли-β-гндрокснбутнрата, преломляющие свет при фазово-контрастной микроскопии. Спор не образуют. Грамотрицательные. Подвижные за счет единственного полярного или субполярного жгутика, Фимбрии не описаны. Аэробы; метаболизм дыхательного типа с использованием кислорода в качестве конечного акцептора электронов. Оптимальная температура 25-30°С. Оптимальный диапазон рН 6-7, однако для штаммов, выделенных из кислой почвы, могут быть оптимальны более низкие значения. Колонии округлые, непрозрачные, редко полупрозрачные, белые, выпуклые, более или менее гранулированной консистенции, диаметром не более 1 мм через 5-7 сут инкубации на агаризованнон среде с дрожжевым экстрактом, манннтолом и минеральными солями. У некоторых штаммов, выделенных из Lotononis bainesii, колонии красные за счет внутриклеточных пигментов. Через 3-5 или больше суток роста в жидкой среде при перемешивании (на качалке) культуры достигают лишь средней плотности. Штаммы с более высокой скоростью роста встречаются редко. Хемоорганотрофы; используют ряд углеводов, из которых как источники углерода наиболее предпочтительны пентозы. Целлюлозу и крахмал не используют. При росте на среде с минеральными солями и маннитолом или одним из многих других углеводов вызывают ее подщелочение. Рост на средах с углеводами обычно сопровождается образованием внеклеточной слизи полисахарндной природы. Некоторые штаммы могут расти хемолитотрофно в присутствии Н2, СО2 и при низком содержании О2. Источниками азота могут служить соли аммония, обычно нитраты и некоторые аминокислоты. Пептон используют слабо (за исключением штаммов, выделенных из Lotononis). Казеин и агар не гидролизуют. В витаминах обычно не нуждаются; как редкое исключение может быть потребность в биотине, который в то же время у ряда штаммов может ингибировать рост. 3-кетоглюкозиды не образуют. Характерное свойство – способность внедряться в корневые волоски бобовых растений (сем. Leguminosae) тропического и в ряде случаев умеренного поясов и вызывать образование корневых клубеньков с бактериями в качестве внутриклеточных симбионтов. Все штаммы обнаруживают сродство к определенному кругу хозяев (специфичность по хозяину). В корневых волосках клетки бактерий имеют раздутую форму, обычную для них в условиях фиксации атмосферного азота с его превращением в доступную для растения связанную форму. Некоторые штаммы фиксируют азот в свободноживущем состоянии при создании специальных условий.
Типовой (и единственный) вид: Bradyrhizobium japonicum.
Признаки вида: Как для рода. В корневых клубеньках бактероиды имеют форму слегка раздутых палочек, иногда разветвленных, или кокков (в Arachts spp). Присутствует один полярный или субполярный жгутик. Обычно вызывают образование клубеньков у видов Glycine (соевые) и Macroptilium artropupureujn (сиратро).
Другие организмы, принадлежащие к Bradyrhizobium: Другие брадиризобии еще не классифицированы как виды или биовары. Эти организмы вызывают образование клубеньков у ряда видов Lotus (L. uliginosus и L. pedunculatus), а также Vigna, Lupinus, Ornithopus, Cicer, Sesbania, Leucaena, Mimosa, Lablad и Acacia; у перечисленных растений клубеньки может индуцировать также быстрорастущий вид Rhizobium loti. Некоторые штаммы в особых условиях фиксируют азот в свободноживущем состоянии. Пока внутри рода Bradyrhizobium не будут описаны новые виды или биовары, предлагается называть эти организмы (отличные от В. japonicum) Bradyrhizobium sp. с указанием в скобках соответствующего растения-хозяина, например, Bradyrhizobium sp. (Vigna) или Bradyrhizpbium sp. (Lupinus).
Род Brucella
Кокки, коккобациллы или короткие палочки, 0,5-0,7 x 0,6-1,5 мкм, расположенные одиночно и, реже, парами, короткими цепочками или небольшими группами. Настоящих капсул не образуют. Настоящего биполярного окрашивания обычно не обнаруживают. Покоящиеся стадии неизвестны. Грамотрицательные. Неподвижные; жгутиков не образуют. Аэробы; метаболизм дыхательного типа. Электронтранспортная цепь содержит цитохромы; конечным акцептором электронов служит кислород или нитрат. Образуют нитратредуктазу. Многие штаммы нуждаются для роста в добавлении СО2, особенно при выделении. Колонии, образующиеся на сывороточно-декстрозном агаре или другой прозрачной среде, просвечивающие, приподнятые, выпуклые, с цельным краем и гладкой, блестящей поверхностью. В проходящем свете имеют бледно-медовый цвет. У видов с гладкими колониями встречаются складчатые варианты, но имеются также виды со стабильно складчатыми колониями и специфическим кругом хозяев. Оптимальная температура 37°С. Рост происходит в диапазоне температуры 20-40°С. Оптимум рН 6,6-7,4. Каталазоположительные и обычно океидазоположительные, но встречаются и отрицательные штаммы. Хемоорганотрофы; большинство штаммов нуждается для роста в сложных средах, содержащих несколько аминокислот, тиамин, никотинамид и ионы магния; у некоторых штаммов можно индуцировать рост на минимальной среде, содержащей в качестве единственного источника азота соль аммония. Сыворотка и кровь стимулируют рост, но гемин (Х-фактор) и никотинамидадениндинуклеотид (NAD; V-фактор) влияния не оказывают. За исключением В. neotomae кислоту из углеводов в обычной среде не образуют. Индол не образуют. Желатину и конденсированную сыворотку не разжижают. Не лизируют эритроциты и не образуют ацетилметилкарбинол (реакция Фогеса-Проскауэра). Проба с метиловым красным отрицательная. Обладают характерными внутриклеточными антигенами, специфичными для рода. Внутриклеточные паразиты, способные заражать широкий круг животных и человека.
Типовой вид: Brucella melitensis.
Дифференциация видов рода Brucella: См. табл., 4,20 и 4.21.
Таблица 4.20. Дифференцирующие признаки видов рода Brucella 1,2 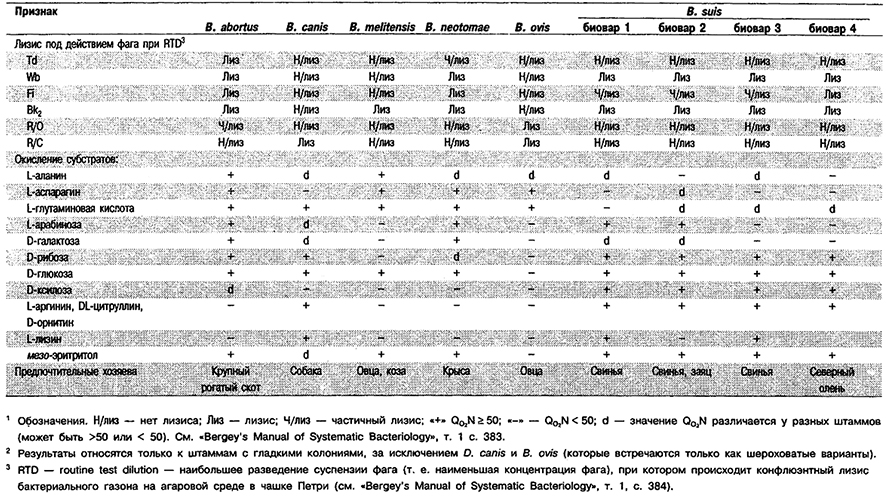
Таблица 4.21. Дифференциация видов и биоваров рода Brucella 1 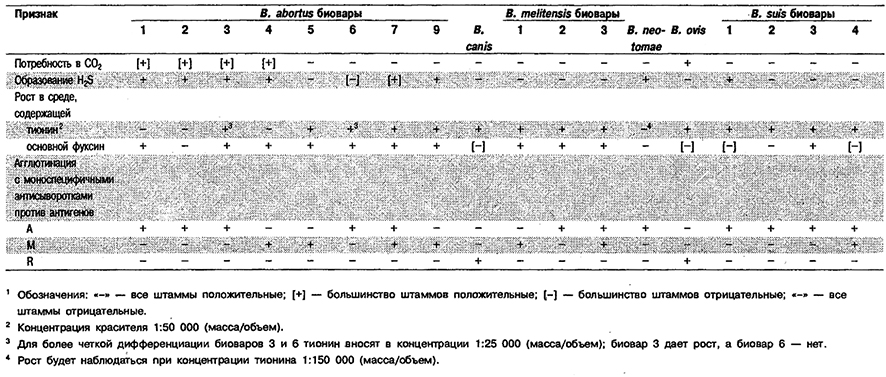
Род Chromohalobacter
От редакции: Род Chromohalobacter не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1989 г. (Ventosa et al., Int. J. Syst. BacterioL 39: 382-386) и включает единственный вид С. marismortui.
Палочки, 0,6-1,0 x 1,5-4,0 мкм при росте в присутствии 10% (масса/объем) NaCl; при более высокой или низкой концентрации соли клетки имеют большую длину. Грамотрицательные. Подвижные за счет перитрнхиальных жгутиков. Аэробы; метаболизм чисто дыхательного типа. Оптимальная температура 37°С, оптимальное значение рН – 7,5. Цвет колоний от фиолетово-синего до коричневого. Каталазоположительные и оксвдазоотрицательные. Хемоорганотрофы. Наилучший рост наблюдается в средах с 10% NaCl, Образуют кислоту из D-глюкозы и других сахаров, Желатину, казеин, Твин-80, крахмал, эскулин и тирозин не гидролизуют. В качестве источников углерода могут использовать углеводы, аминокислоты и некоторые полиолы. Местообитание – Мертвое море и морские природные солеварни.
Типовой (и единственный) вид: Chromohalobacter marismortui.
Признаки вида: Рост происходит на средах, содержащих 1–30% NaCl. Колонии на плотных сложных средах с 10% NaCl округлые, выпуклые, гладкие, с цельным краем и концентрическими кругами – темно-коричневым центром и окружающими его последовательно голубовато-коричневым, серовато-коричневым и желтым кольцами. Образуют желтый и фиолетово-синий пигменты, причем последний не виоласцеин. Пигментообразованию способствуют субоптимальные для роста температуры, присутствие в среде глицерола и 10% NaCl, Диапазон рН 5–10; диапазон температуры 5–45°С. В среде на основе морской воды с общей концентрацией солей 10% для теста на окисление-брожение образуют кислоту, но не газ из D-глюкозы, D-галактозы, мальтозы, лактозы, D-арабинозы, D-ксилозы, сахарозы, трегалозы, глицерола и D-маннитола. Обычно восстанавливают NО3– до NО2–; NО2– не восстанавливают. Растут на агаре Симмонса с цитратом (см. приложение). H2S из цистеина не образуют; фосфатазу, индол, уреазу, ацетоин, ДНКазу, β-гaлактозидазу, фенилаланиндезаминазу, аргининдигидролазу, лизиндекарбоксилазу и орнитиндекарбоксилаэу не образуют. В качестве единственного источника углерода и энергии используют следующие соединения: дульцитол, D-фукозу, D-галактозу, D-глюконат, D-глюкозу, глутамат, минозитол, мальтозу, D-маннитол, D-манноэу, пируват, D-рибозу, сахарозу, D-сорбитол и D-ксилозу, но не N-ацетилглюкозамин, амигдалин, DL-α-аминобутират, бутират, целлобиозу, цитрат, эскулин, n-гидроксибензоат, гиппурат, инулин, малонат, мелибиозу, оксалат, рафинозу, салицилат, салицин и D-тартрат. В качестве единственного источника углерода, азота и энергии используют следующие соединения: L-аланин, DL- аргинин, L-глугамин, L-орнитин, L-пролин, путресцин и L-серин, но не L-аллантоин, бетаин, креатин, этионин, L-изолейцин, L-лейцин, фенилаланин, саркозин, L-треонин и L-валин.
Род Chryseomonas
От редакции: Род Chryseomonas не приведен в «Beigey's Manual of Systematic Bacteriology». Он описан в 1986 г. (Holmes et al., Int. J. Syst. Bacterid. 36: 161-165) и первоначально включал единственный вид С. polytricha. В 1987 г. (Holmes et al., Int. J. Syst. Bacterid. 37: 245-250) название Pseudomonas luteola было признано старшим субъективным синонимом С. polytricha; в связи с этим название вида было изменено на С. luteola.
Палочки с параллельными сторонами и закругленными концами. Внутриклеточных гранул поли-β-гидроксибутирата не содержат. Про стеки или чехлы не образуют. Покоящиеся стадии неизвестны. Грамотрицательные. Подвижные за счет многочисленных (10-12) полярных жгутиков. Аэробы; метаболизм чисто дыхательного типа. Диапазон температуры 18-42°С. При росте на плотных средах цвет биомассы от бледно- до темно-желтого. Колонии обычно округлые (диаметром 1 мм), слегка выпуклые, гладкие (некоторые штаммы образуют складчатые колонии), блестящие, с цельным краем. Каталазоположительные; оксидазоотрицательные. Агар не разлагают. Хемоорганотрофы; катаболизируют сахара по окислительному пути. В природной среде не обнаружены; по-видимому, сапрофиты или комменсалы человека и гомойотермных животных, причем в некоторых случаях могут быть патогенными.
Типовой (и единственный) вид: Chryseomonas luteola.
Признаки вида: Способны расти при 42°С, но не при 5°С, Гемолиз не осуществляют. В тесте на окисление-брожение осуществляют окисление. Гидролизуют трибутирин, Твин-20, тирозин и желатину. На агаризованной среде с лецитовителлином не опалесцируют. Нитрат не восстанавливают. Индол отрицательные; H2S не образуют [тесты с бумагой, пропитанной ацетатом свинца, и на среде с тремя сахарами и железом]. Растут на β-гидроксибутирате (без образования внутриклеточных включений) и на агаре Макконки. Гидролизуют зскулин, но не крахмал. Вызывают подщелочение среды Кристенсена с цитратом. Используют цитрат (агар Симмонса) и малонат. На среде В Кинга флуоресцирующий пигмент не образуют. Глюконат не окисляют. Образуют аргинингидролазу, но не лизиндекарбоксилазу или орнитиндекарбоксилаэу. 3-кетолактозу не продуцируют. Селенит не восстанавливают, Фенилаланиндезаминазу не образуют. Синтезируют β-D-гaлактозидазу, но не фосфатазу. При росте в аэробных условиях в среде с аммонием образуют кислоту из глюкозы, арабинозы, этанола, фруктозы, глицерола, инозитола, мальтозы, маннитола, трегалозы и ксилозы, но не из целлобиозы, дульцитола, лактозы и рафинозы. В среде на основе пептонной воды газ из глюкозы не образуют.
Род Comamonas
От редакции: Род Comamonas не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род был описан в 1985 г. (De Vos et al, Int. J. Syst. Bacteriol. 35: 443-453) и первоначально включал единственный вид С. terrigena. В 1987 г. (Tamoaka, Komagata, Int. J. Syst. Вас ten 1., 37: 52-59) Pseudomon as acidovorans и Pseudomonas testosteroni были pe- классифицированы как виды Comamonas.
Прямые или слегка изогнутые палочки, 0,5-1,0 x 1-4 мкм, одиночные или в парах. Подвижные за счет пучка полярных жгутиков. Эндоспор не образуют. Накапливают внутри клеток поли-β-гидрокс и бутират. Грамотрицательные. Оксидазо- и каталазоположительные. Облигатные аэробы, брожение не осуществляют; хемоорганотрофы. Хорошо растут на средах с органическими кислотами, аминокислотами или пептоном. Флуоресцирующие пигменты не образуют. Использование углеводов встречается редко. Преобладающие жирные кислоты в составе клеток – гексадекановая (16:0), гексадеценовая (16:1) и октадеценовая (18:1). Всегда содержат 3-гидрокеидека- новую кислоту (3-ОН; 10:0). Основной хинон – Q-8. Менахинон не образуют.
Типовой вид: Comamonas terrigena.
Дифференциация видов рода Comamonas: См. табл. 4.22.
Таблица 4.22. Дифференциация видов рода Comamonas 1,2 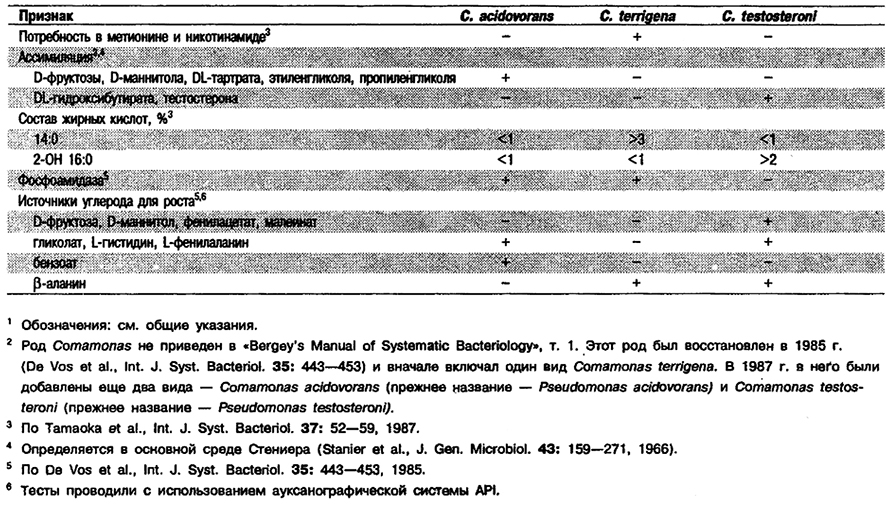
Род Cupriavidus
От редакции: Род Cupriavidus не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1987 г. (Makkar, Casida, Int. J. Syst. Bacteriol. 37: 323-326) и включает один вид С. necator.
Кокковидные палочки, 0,7-0,9 x 0,9-1,3 мкм. Грамотрицательные. Подвижные за счет 2-10 пернтрихиальных жгутиков. Хемогетеротрофы. В органических источниках азота не нуждаются. Глюкозу не используют; фруктозу катаболизируют путем окисления. Каталазо- и окисидазоположительные. Восстанавливают нитрат. Желатину, крахмал и мочевину не гидролизуют. Индол и H2S не образуют. В качестве единственного источника углерода и азота для роста могут использовать многие аминокислоты, но не L-лизин и L-метионин. Оптимальная температура 27°С. Оптимальный диапазон рН 7,0-8,0. NaCl в концентрации 3% ингибирует рост. Устойчивы к меди. Начало роста стимулируется медью. Колонии на МПА через 2 сут роста при 27°С беловатые, блестящие, слизистые, гладкие, выпуклые, с цельным краем, диаметром 2-4 мм. Выделены из почвы. Необлигатные хищники, вызывающие лизнс в почве разнообразных грамположительных и грамотрицательных бактерий. Могут лизировать клетки некоторых других необлигатных бактерий-хищников. Для роста в присутствии вида-жертвы не нуждаются.
Типовой (и единственный) вид: Cupriavidus hecator.
Признаки вида: Как для рода. При лабораторном культивировании С. necator может давать небольшое число клеток неслизистого варианта. Этот вариант похож на слизистый, но отличается более сухими, уплощенными и мелкими колониями.
Род Deleya
От редакции: Род Deleya не приведен в «Bergey's Manual оГ Systematic Bacteriology». Он описан в 1981 г, (Baumann et al Int. J. Syst. Bacteriol. 33: 793-802) в составе пяти видов: D. marina (прежнее название – Pseudomonas marina), D. aesta (прежнее название – Alcaligenes aestus), D. pacifica (прежнее название – Alcaligenes pacificus), D. cupida (прежнее название – Alcaligenes cupidus) и D. venusta (прежнее название – Alcaligenes venustus). Как типовой был определен вид D. aesta. В 1984 г, добавлен шестой вид D. halophila (Quesada et aL, Int. J. Syst. Bacteriol. 34: 287-292). В 1989 г. установлено (Akagawa, Yamasato, Int. J. Syst. Bacteriol, 39: 462-466), что D. aesta, Alcaligenes aquamarinus и Alcaligenes faecalis subsp. homari принадлежат к одному виду и, в связи с приоритетностью видового эпитета aqumarinus, вид D. aesta переименован в D. aquamarina.
Прямые палочки, 0,8-1,1 x 1,5-3,0 мкм. Накапливают в качестве внутриклеточного запасного вещества поли-β-гидроксибутират. В старых культурах и при неблагоприятных условиях культивирования обычно присутствуют инволюционные формы. Эндоспор и микроцист не образуют. У большинства видов клетки подвижные за счет 4–12 перитрихиальных жгутиков, у одного вида (D. marina) – за счет 2-5 полярных жгутиков. Облигатные аэробы; универсальным акцептором электронов служит молекулярный кислород. Все виды – хемоорганотрофы, способные расти в минеральной среде на основе морской воды с добавлением D-глюкозы и NH4Cl. Все виды облигатно нуждаются в Na+; минимальная необходимая для хорошего роста концентрация варьирует в пределах 75-200 мм. Все виды используют D-глюкозу, ацетат, сукцинат, DL-β-гидроксибутират, лактат, глицерол и D-аланин. Все виды растут при 35°С. Ни один из видов не образует внеклеточную липазу, желатиназу или хитиназу. Ни один из видов не способен использовать Н2 в качестве единственного источника энергии и СО2 в качестве единственного источника углерода. У всех видов отсутствует аргинидигидролазная система. Обнаружены в морских местообитаниях; один вид (D. halophila) обнаружен в сильно засоленных почвах.
Типовой вид: Deleya aquamarina.
Дифференциация видов рода Deleya: См. табл. 4.23.
Таблица 4.23. Дифференцирующие признаки видов рада Deleya1 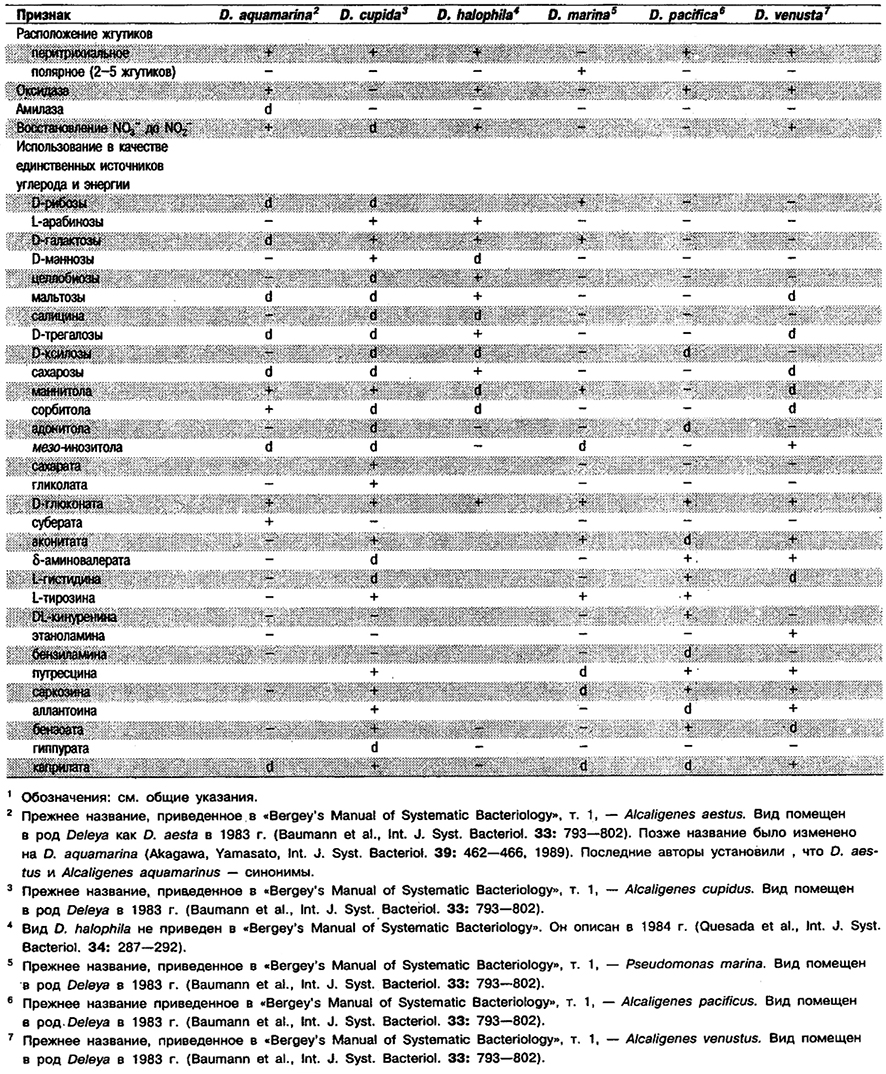
Род Derxia
Клетки палочковидные, с закругленными концами, 1,0-2,0 x 3,0-6,0 мкм, одиночные или в коротких цепочках. В большей или меньшей степени плеоморфные в зависимости от возраста и состава среды. В стареющих культурах клетки часто остаются вместе, образуя длинные нити, иногда с локально раздутыми или искривленными в отдельных местах клетками. Некоторые клетки могут достигать огромных размеров (до 30 мкм). В молодых клетках цитоплазма гомогенная, старые заполнены характерными крупными, преломляющими свет тельцами. Покоящиеся стадии неизвестны.
Грамотрицательные. Подвижные за счет единственного короткого полярного жгутика. Подвижные клетки образуются в большом количестве при росте в жидкой среде, содержащей глюкозу и источник связанного азота, но редко на плотной среде, дефицитной по азоту. Аэробы; метаболизм чисто дыхательного типа с использованием кислорода в качестве конечного акцептора электронов. Фиксируют молекулярный азот в аэробных условиях, а также при пониженном давлении кислорода (микроаэробные условия). Оптимальный диапазон температуры 25-35°С; при 15°С рост медленный, при 40°С – слабый, при 50°С роста нет. Диапазон рН для роста 5,5-9,0; при рН 4,4 роста нет. Культуры, растущие в жидкой среде» превращаются в гелеобразную массу, однако вблизи поверхности рост более интенсивный с образованием толстой, плотной пленки. Колонии на агаризованной среде вначале слизистые и полупрозрачные, позднее массивные и непрозрачные, высоко приподнятые, с морщинистой поверхностью. Старые колонии имеют темный красновато-коричневый цвет. Каталазоотрицательные. При росте в щелочной среде окисляют большое число сахаров, спиртов и органических кислот с образованием в основном СО2 и небольшого количества кислоты, вероятно уксусной. Могут расти как факультативные автотрофы с использованием водорода. Получен рост с использованием метана или метанола в качестве единственного источника углерода. Обнаружены в тропических почвах (Азия, Африка, Южная Америка).
Типовой (и единственный) вид: Derxia gummosa.
Признаки вида: Как для рода.
Род Ensifer
Клетки палочковидные, 0,7-1,1 х 1,0-1,9 мкм, одиночные или в парах. Подвижные за счет пучка из 3-5 субполярных жгутиков. Размножаются почкованием: почка возникает на одном из концов клетки и удлиняется, давая асимметричный полярный рост. Разделение клеток происходит путем бинарного деления. После деления рост возобновляется за счет новых почек на вновь образовавшихся полюсах как материнской, так и дочерней клеток. Грамотрицательные, но окраска может быть слабой. Аэробы; метаболизм чисто дыхательного типа. Хемогетеротрофы; не требовательны к составу среды. Колонии на агаре с 0,1% сердечной вытяжки 10-15 мм в диаметре, серовато-белые, выпуклые, слизистые, влажные, непрозрачные, округлые с волнистым краем, могут казаться почти прозрачными из-за обильного образования слизи. Слабо каталазоположительные. Оптимальная температура 27°С. Растут при 20 и 37°С. Необлигатные хищники, прикрепляющиеся концом клетки к различным живым грамотрицательным и грамлоложительным бактериям-хозяевам; специальных концевых структур для прикрепления не имеют. В зависимости от рН (оптимальный 6), концентрации питательных веществ в среде и вида клеток-хозяев могут убивать и часто также лизировать клетки-жертвы. Прикрепление к клеткам бактерий и их лизис наблюдаются как в природных условиях в почве, так и in vitro в чистых культурах. При росте на агаризованных средах выделяют диффундирующий фактор, который вызывает лизис клеток-хозяев. Используют разнообразные источники углерода, в том числе глюкозу, галактозу, маннозу, рамнозу, ксилозу, маннитол, глицерол, сорбитол, L-глутаминовую кислоту, L-аланин, L-аспарагин и L-глутамин. Восстанавливают нитрат и нитрит. Нитрогеназная активность отсутствует. Растут на чистой желатине, не осуществляя гидролиз. NaCl в концентрации 4%, но не 2%, ингибирует рост. Растут на агаре с дезоксихолатом; цвет колоний вначале беловато-красный, а затем меняется на желтовато-коричневый. Вьщелены из почвы.
Типовой (и единственный) вид: Ensifer adhaerens.
Признаки вида: Как для рода. Из культур выделены варианты, продуцирующие меньше слизи и образующие поэтому более мелкие, суховатые и слегка беловатые колонии.
Род Erythrobacter
Клетки палочковидные, подвижные за счет субполярных жгутиков. Грамотрицательные. Содержат бактериохлорофилл α, однако в анаэробных условиях на свету не растут. Колонии и суспензии клеток окрашены в оранжевый цвет из-за присутствия каротиноидных пигментов. Аэробные хемоорганотрофы с преимущественно дыхательным типом метаболизма. В микроаэробных условиях при использовании разнообразных углеводов образуют небольшое количество кислоты. Каталазо- и оксндазоположительные. Для роста нуждаются в биотине. Метанол не используют. Реакция Фогеса-Проскауэра и проба с метиловым красным отрицательные. Гидролизуют желатину. Встречаются в морской среде, содержащей О2, преимущественно на морских водорослях.
Типовой (и единственный) вид: Erythrobacter longus.
Признаки вида: Размеры клеток 0,3-0,4 х 2,0-5,0 мкм. См. также табл. 4.45.
