Клетки сферические, спиральные, палочковидные или вибриоидные, диаметром 0,3–6,0 мкм, одиночные либо в агрегатах правильной или неправильной формы. Встречаются также одноклеточные или однорядные многоклеточные нитчатые формы. Грамотрицательные. Размножение в большинстве случаев происходит путем бинарного деления; для некоторых видов характерно размножение почкованием. Газовые вакуоли присутствуют либо отсутствуют. Подвижные или неподвижные; движение обеспечивается за счет жгутиков или происходит путем скольжения. Жгутикование монотрихиальное или мультитрихиальное. Фотосинтеэирующие пигменты локализованы в цитоплазматической мембране (Подгруппа 4), во внутрицитоплазматических мембранных системах различных типов (Подгруппы 1–3) или в хлоросомах (Подгруппы 5 и 6). Цвет клеточных суспензий от пурпурно-фиолетового до пурпурно-красного, розово-красный, желтовато-коричневый, коричневый и зеленый. Общее свойство для всех видов – присутствие бактернохлорофиллов (см. табл. 10.2). Фотоавтотрофы или фотоорганотрофы в анаэробных или микроаэробных условиях. В противоположность оксигенному фотосинтезу цианобактерий аноксигеннын фотосинтез зависит от внешних доноров электронов, таких как восстановленные соединения серы, молекулярный водород или органические соединения. При окислении сульфида временно откладываются либо внутри (Подгруппа 1), либо вне клеток (Подгруппы 2 и 5) сильно преломляющие свет глобулы серы. Запасными веществами служат полисахариды, поли-β-гидроксибутират и полифосфаты. Диоксид углерода фототрофные бактерии ассимилируют через восстановительный пентозо-фосфатный цикл или восстановительный цикл лимонной кислоты (Подгруппа 5). В качестве источника азота обычно используют соли аммония. Фиксация молекулярного азота продемонстрирована у большинства представителей всех подгрупп, кроме Подгруппы 6. В аэробных или микроаэробных условиях многие виды, за исключением относящихся к Подгруппе 5, способны расти как хемоавтотрофы или хемоорганотрофы. Донорами электронов и источниками углерода служат жирные кислоты, органические кислоты или спирты. Места обитания – бескислородные зоны увлажненных почв и водных сред, включая пресноводные, солоноватые, морские и сильно соленые водоемы.
Таблица 10.1. Характеристики максимумов поглощения бактериохлорофиллов в живых клетках
Таблица 10.2. Группы каротиноидов аноксигенных фототрофных бактерий
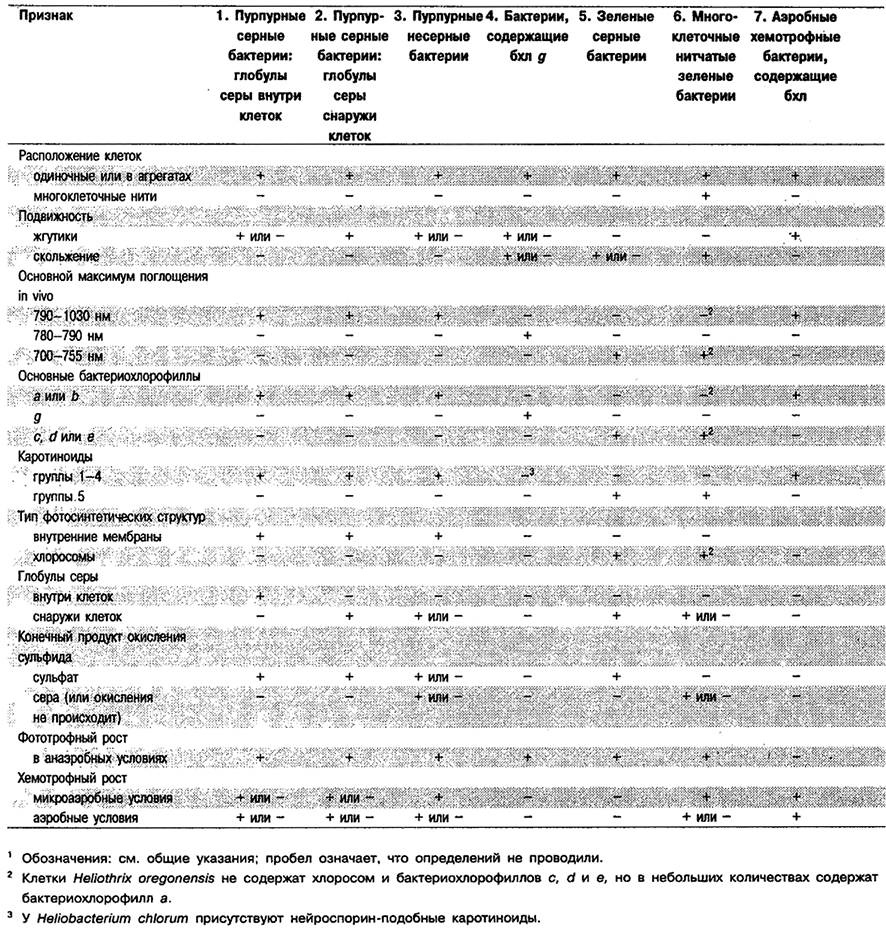
Аноксигенные фототрофные бактерии разделены на семь подгрупп, которые можно дифференцировать по признакам, приведенным в табл. 10.3.
Таблица 10.3. Дифференциация Подгрупп 1–7 аноксигенкых фототрофных бактерий1
Подгруппа 1
Клетки способны расти с использованием сульфида и серы в качестве единственных доноров электронов для ассимиляции СО2; хорошо растут в фототрофных условиях. В присутствии одновременно сульфида и света внутри клеток появляются глобулы серы, которая впоследствии может окисляться до сульфата. Содержат бактериохлорофиллы α и Ь и каротиноиды групп 1–4 (табл. 10.2). Могут нуждаться для роста в витамине В12.
Дифференциация родов в Подгруппе 1: См. табл. 10.4.
Таблица 10.4. Дифференцирующие признаки родов Подгруппы 11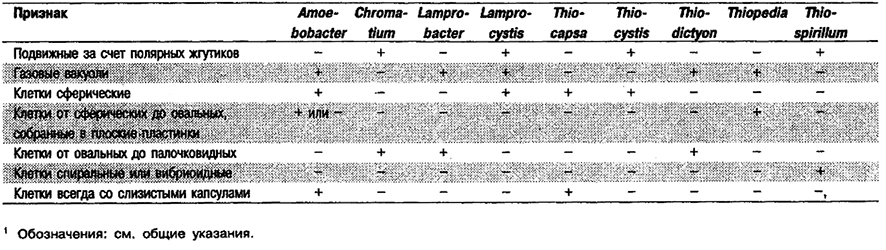
Род Amoebobacter
Клетки сферические, диаметром 1,5–3,0 мкм, одиночные или в агрегатах неправильной либо пластинчатой формы. Клетки окружены слизью. Неподвижные. Содержат газовые вакуоли, расположенные в центре или на периферии клеток. Культуры окрашены в розово-красный цвет, у одного вида – в пурпурно-красный. Внутрицитоплазматические мембранные системы везикулярного типа. Фотосинтетические пигменты – бактериохлорофилл α и каротиноиды спириллоксантиновой или океноновой серии. В анаэробных условиях фототрофы; в микроаэробных или аэробных условиях способны к хемотрофии. При фотоавтотрофном росте используют сульфид, серу, тиосульфат или молекулярный водород. В случае окисления сульфида временно накапливают внутри клеток глобулы серы. Конечный продукт окисления – Сульфат. Осуществляют . фотоассимиляцию ацетата, прбпионата, пирувата или лактата. Для роста могут нуждаться .в витамине B12 Диапазон рН д|я роста 6,8–7,5; диапазон, температуры – 20–35°С. Вместе с Thiocapsa могут вызывать цветение воды в загрязненных сточными водами лагунах с высоким содержанием; органических веществ. Amoebobacter purpureas встречается в основном в содержащей сульфид воде стратифицированных пресноводных озер.
Типовой вид: Amoebobacter roseus.
Дифференциация видов рода Amoebobacter: См. табл. 10.5.
Таблица 10.5. Дифференцирующие признаки видов рода Amoebobacter1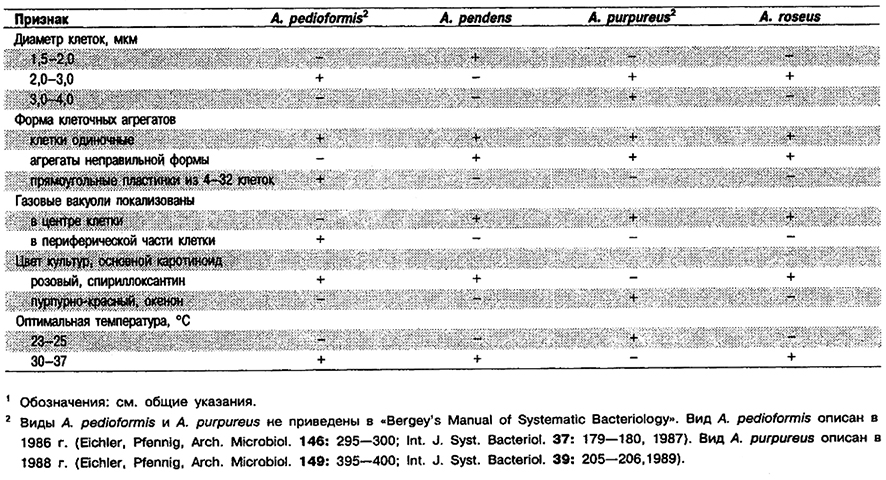
Род Chromatium
Клетки палочковидные, от прямых до слегка изогнутых, или овальные, с закругленными концами, диаметром 1,0–6,0 мкм и длиной 1,5–16,0 мкм (рис. 10.1). Подвижные за счет одного или многочисленных полярных жгугиков. В присутствии сульфида и света внутри клеток, которые не содержат газовых вакуолей, появляются равномерно распределенные глобулы серы. Внутрицитоплазматические мембранные системы везикулярного типа. Фотосинтезирующие пигменты – бактериохлорофилл α и каротиноиды групп 1, 3 и 4. В анаэробных условиях растут фотоавтогрофно с использованием сульфида или серы в качестве доноров электронов для ассимиляции СО2. Конечный продукт окисления – сульфат. Некоторые виды могут также использовать молекулярный водород или тиосульфат. Все виды фотоассимилируют ряд простых органических соединений в присутствии сульфида и бикарбоната; наиболее широко используемые субстраты – ацетат-и пируват. Могут нуждатьсядля роста в витамине В12. Оптимальная температура 20–35°С; только, один вид, Chromatium tepidum, растет при 50° С. Диапазон рН для роста 6,8–7,5. Виды Chromatium встречаются в содержащих сульфид участках пресноводных водоемов, эстуариев, морей и сильно соленых сред.
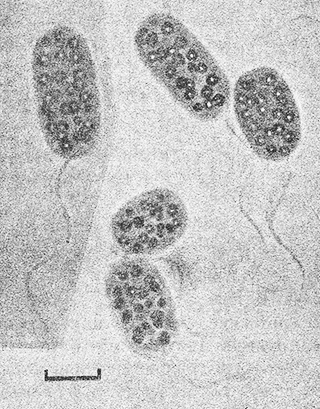
Рис. 10.1. Клетки Chromatium okenii DSM, выращенные фотоавтотрофно в среде с сульфидом. Видны равномерно распределённые внутриклеточные глобулы элементной серы и спиральные пучки жгутиков. Светлое поле. Шкала – 5 мкм.
Типовой вид:с Chromatium okenii.
Дифференциация видов рода Chromatium: См. табл. 10.6.
Таблица 10.6. Дифференцирующие признаки видов рода Chromatium1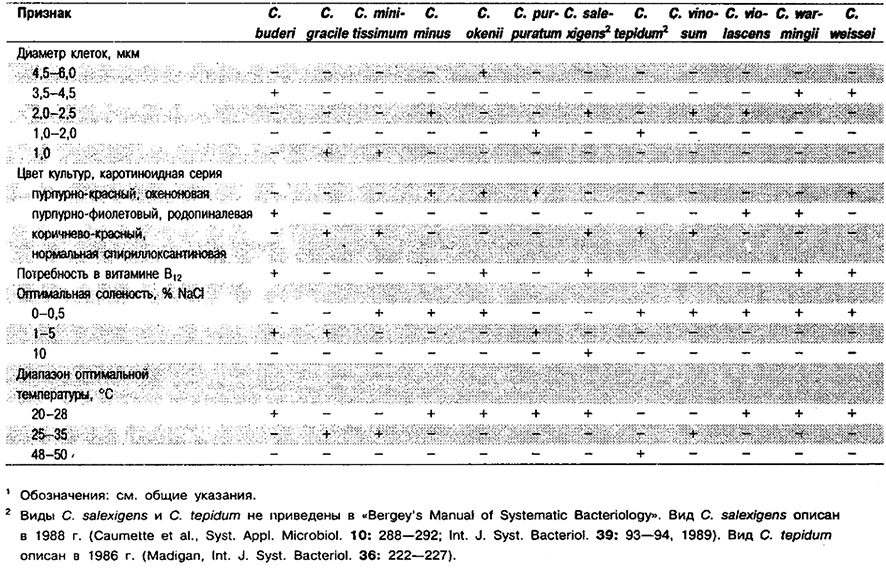
Род Lamprocystis
Клетки от сферических доовальных, диаметром 2–3,5 мкм, одиночные, в небольших агрегатах неправильной формы или в длинных и ветвящихся агрегатах, окруженных слизью (рис. 10.2). В центральной части клеток присутствуют газовые вакуоли. В благоприятных для роста условиях агрегаты могут распадаться на более мелкие скопления и сферические клеточные колонии, которые становятся подвижными; высвобождаются также отдельные подвижные клетки. Клетки подвижные за счет единственного жгутика. Культуры окрашены в пурпурно-фиолетовый цвет. Содержат внутрицитоплазматическую мембранную систему везикулярного типа. Содержат бактериохлорофилл α и каротиноиды родопиналевой серии. В анаэробных условиях осуществляют фотоавтотрофный рост с использованием сульфида или серы в качестве доноров электронов для ассимиляции СО2; конечный продукт окисления – сульфат. При окислении сульфида в периферической части клеток временно Накапливаются глобулы серы. В присутствии сульфида и бикарбоната фотоассимилируют ацетат и пируват. Могут нуждаться в витамине В12. Температура для ,роста 20–30°С; диапазон рН 6,8–7,5. Встречаются в иле и стоячей воде прудов и озер, содержащих сероводород; распространены как – планктонные бактерии в верхнем слое сульфидсодержащего гиполимниона стратифицированных пресноводных озер.
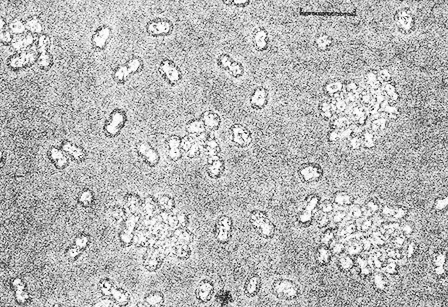
Рис. 10.2. Клетки Lamprocystis roseopersicina DSM 229, выращенные фотоавтотрофно в среде с сульфидом. Отдельные клетки и клеточные колонии подвижны в жидкой среде. Светлые области неправильной формы внутри клеток — газовые вакуоли. Мелкие сферические тела в клетках — глобулы серы. Фазовый контраст. Шкала — 10 мкм.
Типовой (и единственный) вид: Lamprocystis roseopersicina.
Признаки вида: Как для рода.
Род Thiocapsa
Клетки от сферических до почти овальных, диаметром 1,0–З,0 мкм, одиночные или в агрегатах из 2, 4 и более с клеток; клетки обычно окружены слизью (рис. 10.3). Неподвижные; газовых вакуолей не содержат. Культуры розово-красные или желто-коричневые. Внутрицитоплазматические мембранные системы везикулярного или тубулярного типа. Содержат бактериохлорофилл α либо b и каротиноидьг сгшриллоксантиновой серии. В анаэробных условиях осуществляют фотоавтотрофный рост с использованием сульфида или серы в качестве донора электронов для ассимиляции СО2; конечный продукт окисления – сульфат. При окислении сульфида внутри клеток временно образуются глобулы серы. Фотоассимилируют ацетат, пируват и другие органические соединения. В аэробных условиях в темноте растут как облигатные фототрофы или факультативные хемоавтотрофы. Температура для роста 20–35°С; диапазон рН 6,8–7,5. Обнаружены в стоячей воде и иле прудов, луж и загрязненных лагун, содержащих органические вещества и сероводород; обычно присутствуют также в эстуариях, в песке морских побережий и в микробных матах соляных маршей.
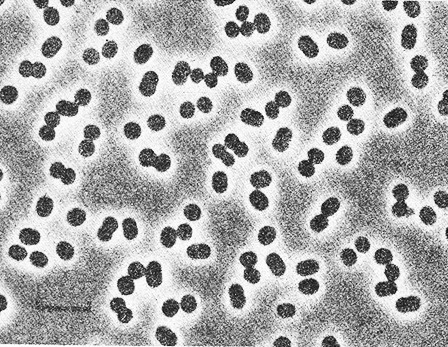
Рис. 10.3. Клетки Thiocapsa roseopersicina DSM 219, выращенные фотоавтотрофно в среде с сульфидом. Фазовый контраст. Шкала — 5 мкм.
Типовой вид: Thiocapsa roseopersicina.
Дифференциация видов рода Hiiocapsa: См. табл. 10.7.
Таблица 10.7. Дифференцирующие признаки видов рода Thiocapsa1
Род Thiocystis
Клетки сферические или слегка овальные, диаметром 2,5–3 мкм. Могут встречаться отдельные клетки бблыиих размеров. Перед делением могут иметь форму диплококков (рис, 10.4). Подвижные за счет единственного жгутика. В присутствии сульфида в условиях высокой освещенности образуют агрегаты неправильной формы из неподвижных клеток, окруженных слизью. Газовых вакуолей не содержат. Культуры окрашены в пурпурно-фиолетовый или пурпурно-красный цвет. Фотосинтетические мем- бранные системы везикулярного типа. Содержат бактериохлорофилл а и каротиноиды групп 3 и 4. В анаэробных условиях растут как фотоавтотрофы с Использованием сульфида или серы в качестве донора электронов для ассимиляции СО2. При окислешш сульфида внутри клеток временно образуются глобулы серы. Конечный продукт окисления – сульфат. В присутствии сульфида и бикарбоната фотоассимилируют ацетат и гтируват. В микроаэробных или аэробных условиях в темноте растут как факультативные хемоавтотрофы. Диапазон рН 6,8–7,6. Температура для роста 25–35°С. Морские штаммы нуждаются в 1–2% NaCl. Встречаются в воде, содержащей сульфид, и в иле пресноводных и солоноватых водоемов, а также в морских местообитаниях: лагунах, загрязненных сточными водами, эстуариях, соляных маршах и серных источниках.
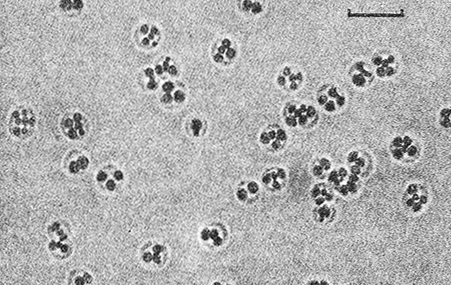
Рис. 10.4. Клетки Thiocystis violaceae DSM 207, выращенные фотоавтотрофно в среде с сульфидом. Клетки содержат глобулы серы. Светлое поле. Шкала – 5 мкм.
Типовой вид; Thiocystis violacea.
Дифференциация видов рода Thiocystis:
1. Культуры пурпурно-фиолетовые; каротиноиды родопиналевой ceрии; глобулы серы в центре клетки.
Т. violacea
2. Культуры пурпурно-красные; каротиноиды океноновой серии; глобулы серы только в цитоплазме по периферии клетки.
Т. gelatinosa
Род Thiodictyon
Клетки палочковидные, с закругленными концами, 2,0 х 4–8 мкм, неподвижные во всех условиях.
Могут образовывать агрегаты в виде сетевидных структур неправильной формы, в которых клетки расположены конец в конец (рис. 10.5). Могут также образовывать более компактные скопления; агрегаты могут распадаться на отдельные клетки. Содержат крупные газовые вакуоли в центральной части клеток. Культуры окрашены в пурпурно-фиолетовый цвет. Фотосинтетические мембранные системы везикулярного типа. Содержат бактериохлорофилл а и каротиноиды родопиналевой серии. Облигатные анаэробы и фототрофы, Растут фотоавтотрофно с использованием сульфида или серы в качестве донора электронов для ассимиляции СО2. При окислении сульфида временно образуются глобулы серы в периферической, свободной от газовых вакуолей части клеток. Конечный продукт окисления – сульфат. В присутствии сульфида и бикарбоната фотоассимилируют ацетат и пируват. Оптимум рН 7,3; температура для роста 20–30°С. Встречаются в воде, содержащей сульфид, и в иле прудов и озер; обычно присутствуют в верхнем слое планктона сульфид- содержащего гиполимниона стратифицированных пресноводных озер.
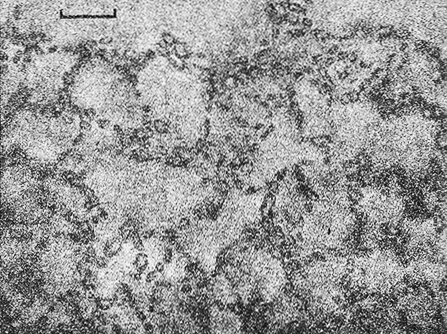
Рис. 10.5. Клетки Thiodictyon elegans DSM 232, типично расположенные в виде сети. Светлое поле. Шкала – 12 мкм.
Типовой вид: Thiodictyon elegans.
Дифференциация видов рода Thiodictyon:
В присутствии сульфида в условиях сильной освещенности клетки способны расти в форме типичных сетевидных агрегатов.
Т. elegans
Клетки одиночные, не образующие сетевидных агрегатов.
Т. bacillosum
Род Thiopedia
Клетки от сферических до овальных или удлиненных овальных, 1,4–1,8 х 1,5–2,5 мкм; образуют прямоугольные пластинки из 4, 8, 16, 32 или более клеток. Свойство образования пластинок может быть утрачено в результате длительного культивирования в лаборатории. Неподвижные. В центральной части клеток присутствуют газовые вакуоли неправильной формы. Культуры окращены в ярко-пурпурно-красный цвет. Фотосинтетическая мембранная система везикулярного типа. Содержат бактериохлорофилл α и каротиноиды окено- новой серии. Облигатные анаэробы и фототрофы. Фотоавтотрофный рост возможен в присутствии сульфида в концентрации до 0,6 мМ; в более высоких концентрациях сульфид полностью ингибирует рост. При окислении сульфида в периферической части клеток временно накапливаются глобулы серы. Конечный продукт окисления – сульфат. В присутствии небольшого количества сульфида (0,1–0,5 мМ) и бикарбоната способны осуществлять фотоассимиляцию ацетата, бутирата или валерата (в концентрации не выше 3 мМ). Использование двух последних жирных кислот характерно для Thiopedia в сравнении с Amoebobacter purpureus. Рост стимулируется при добавлении 100 мкМ дитионита. Нуждаются для роста в витамине В12, Оптимальный рН 7,3; оптимальная температура 20°С. Свет интенсивностью выше 200–300 лк (лампа накаливания) ингибирует рост. Местообитание – ил и стоячая вода прудов и озер, содержащие сероводород; распространены как планктонные бактерии в верхнем слое сульфиде одержащего гиполимниона стратифицированных пресноводных озер.
Типовой (и единственный) вид: Thiopedia rosea.
Признаки вида: Как для рода.
Род Thiospirillum
Клетки в виде изогнутых палочек или вибриоидные, сигмоидные либо спиральные с закругленными концами, диаметром 2,5–4,0 мкм. Длина сигмоидных клеток обычно 30–40 мкм; спиральные клетки достигают в длину 100 мкм. Глубина колец 3–7 мкм; могут встречаться почти прямые палочковидные клетки. Подвижные за счет пучка многочисленных полярных жгутиков длиной 10–12 мкм. Редко клетки имеют пучки жгутиков на обоих концах. В присутствии сульфида на свету клетки содержат равномерно распределенные в цитоплазме глобулы серы. Газовых вакуолей не содержат. Внутрицитоплазматическая мембранная система везикулярного типа. Клеточные суспензии окрашены в желтовато-коричневый цвет. Содержат бактериохлорофилл α и каротиноиды ликопин и родопин. Облигатные анаэробы и фототрофы. Фотоавтотрофный рост происходит с использованием сульфида или серы в качестве донора электронов для ассимиляции СО2. При окислении сульфида в клетках временно накапливаются глобулы серы; конечный продукт окисления – сульфат. В присутствии сульфида и бикарбоната осуществляют фотоассимиляцию ацетата. Нуждаются для роста в витамине B12. Диапазон рН для роста 6,8–7,5. Оптимальный рН 7,3; температура для роста 20–28°С. Обнаружены в иле и стоячей воде канав и пресноводных водоемов, содержащих сероводород.
Типовой (и единственный) вид: Thiospirillum jenense.
Признаки вида: Как для рода.
Подгруппа 2
Клетки способны расти с использованием сульфида и серы в качестве единственных фотосинтетических доноров электронов для ассимиляции СО2. В присутствии сульфида и света снаружи клеток появляются глобулы серы, которая может впоследствии окисляться до сульфата. Как источник азота используют аммиак и молекулярный азот. Рост зависит от солености и щелочности среды. Содержат бактериохлорофиллы α и b и каротиноиды группы 1 (табл. 10.2).
Подгруппа 2 включает только один род.
Род Ectothiorhodospira
Клетки вибриондные или палочковидные, диаметром 0,5–1,5 мкм, подвижные за счет полярных жгутиков. Размножение происходит путем бинарного деления. Внутренние фотосинтетические мембраны в виде стопок ламелл, связанных с цитоплазматической мембраной. Фотосинтетические пигменты – бактериохлорофиллы α и b и каротиноиды. Растут фотоавтотрофно в анаэробных условиях с использованием восстановленных соединений серы или молекулярного водорода в качестве доноров электронов или фотогетеротрофно с использованием ограниченного числа простых органических соединений. Сульфид окисляют до серы, которая откладывается снаружи клеток и может быть в дальнейшем окислена до сульфата. Некоторые виды способны расти в темноте в микроаэробных или аэробных условиях. Рост зависит от солености и щелочности среды. В факторах роста не нуждаются, но витамин В12 стимулирует рост некоторых штаммов. Оптимальная температура для роста 25–44°С, оптимальный рН 7,6–9,5. Встречаются в морской и экстремально соленых средах, содержащих сульфид и имеющих рН от нейтрального до экстремально щелочного, например в эстуариях, соляных равнинах, соленых озерах, щелочных озерах и других; иногда могут быть обнаружены в почве.
Типовой вид: Ectothiorhodospira mobilis.
Дифференциация видов рода Ectothiorhodospira: См. табл. 10.8.
Таблица 10.8. Дифференцирующие признаки рода Ectothiorhodospira 1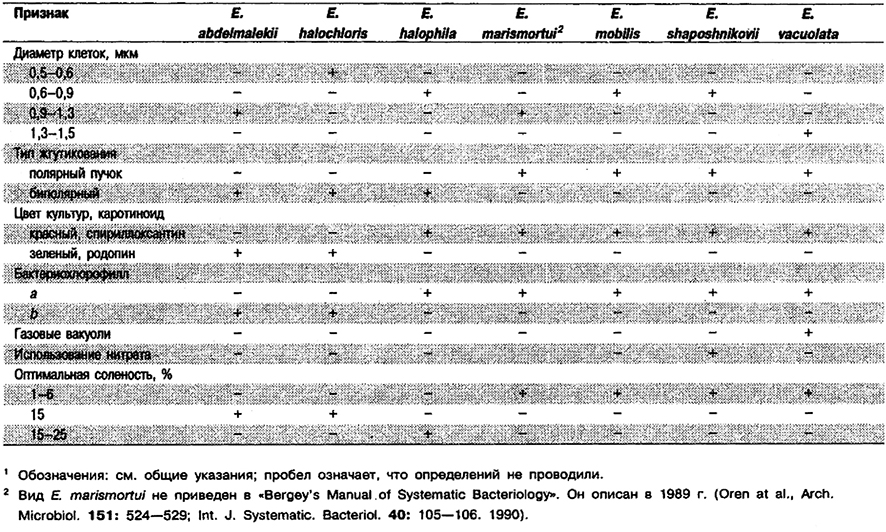
Подгруппа 3
Клетки растут предпочтительно за счет фотоассимиляции простых органических веществ; для некоторых видов характерна способность использовать сульфид или тиосульфат в качестве донора электронов для ассимиляции СО2. В присутствии сульфида и света возможно появление глобул серы только снаружи клеток, никогда не внутри. Серу редко окисляют дальше до сульфата. Представители большинства родов способны расти как хемогетеротрофы в микроаэробных или аэробных условиях. В качестве источника азота использут аммиак или молекулярный азот. Рост бактерий большинства родов зависит от одного или большего числа факторов роста; наиболее часто нуждаются в биотине, тиамине, ниацине и н-аминобензойной кислоте.
Дифференциация родов в Подгруппе 3: См, табл. 10.9.
Таблица 10.9. Дифференцирующие признаки родов Подгруппы 31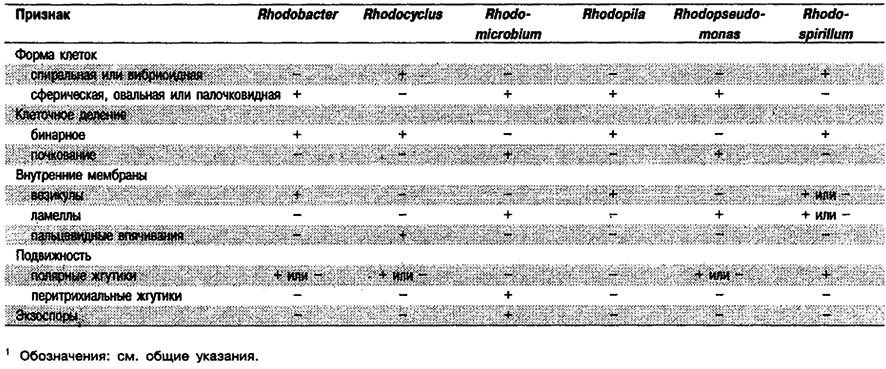
Род Rhodobacter
Клетки овальные или палочковидные, диаметром 0,5–1,2 мкм, подвижные или неподвижные; подвижные формы имеют полярные жгутики. Размножаются путем бинарного деления; могут образовывать капсулы и слизь; способны образовывать цепочки клеток. Внутриклеточные фотосинтетические мембраны везикулярного типа. Фотосинтетические пигменты – бактериохлорофилл а и каротиноиды группы 2 (табл. 10.2). Культуры окрашены в желтовато-коричневый цвет в анаэробных условиях и красновато-коричневые в присутствии воздуха. Фотоавтотрофный рост возможен в присутствии сульфида как донора электронов, а у некоторых видов в присутствии тиосульфата и молекулярного водорода. Растут фотогетеротрофно в анаэробных условиях на свету с использованием разнообразных органических соединений в качестве источников углерода и электронов, Большинство видов растут хемогетеротрофно в аэробных условиях. Нуждаются для роста в витаминах. Оптимальная температура для роста 25–35°С, оптимальный рН 6,5–7,5.
Типовой вид: Rhodobacter capsulatus.
Дифференциация видов рода Rhodobacter: См. табл. 10.10.
Таблица 10.10. Дифференцирующие признаки видов рода Rhodobacter 1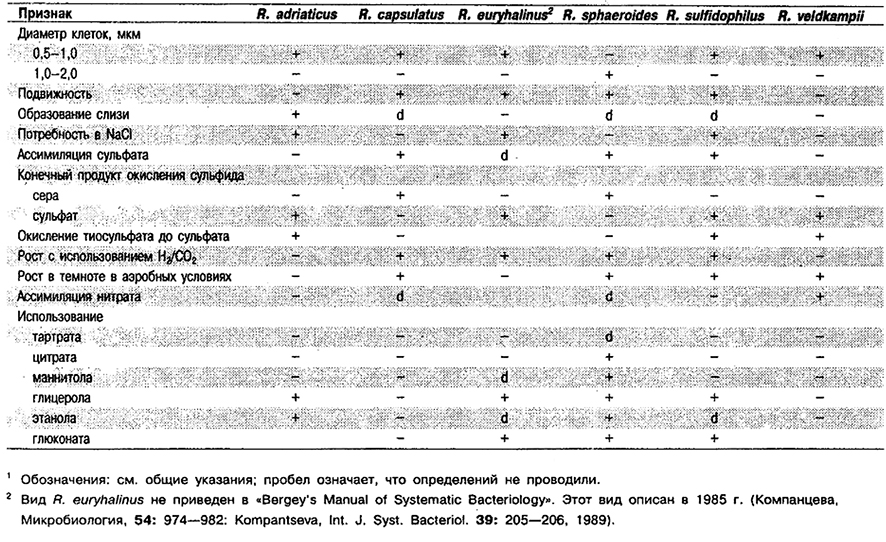
Род Rhodocyclus
Изогнутые или прямые, тонкие палочки диаметром 0,3–1,0 мкм, неподвижные или подвижные за счет полярных жгутиков. Размножаются бинарным делением. Внутриклеточные фотосинтетические мембраны в виде небольших одиночных пальцевидных впячиваний цитоплазматической мембраны. Фотосинтетические пигменты – бактериохлорофилл α и каротиноиды: Возможен фотоавтотрофный рост с использованием молекулярного водорода. Предпочтительно растут фотогетеротрофно в анаэробных условиях на свету с использованием, различных органических субстратов как источников углерода и доноров электронов. Для роста, нуждаются в витаминах. Оптимальная температура 30°С, оптимальный рН 6,5–7,5.
Типовой вид: Rhodocyclus purpureus.
Дифференциация видов рода Rhodocyclus: См. табл. 10.11.
Таблица 10.11. Дифференцирующие признаки видов рода Rhodocyclus 1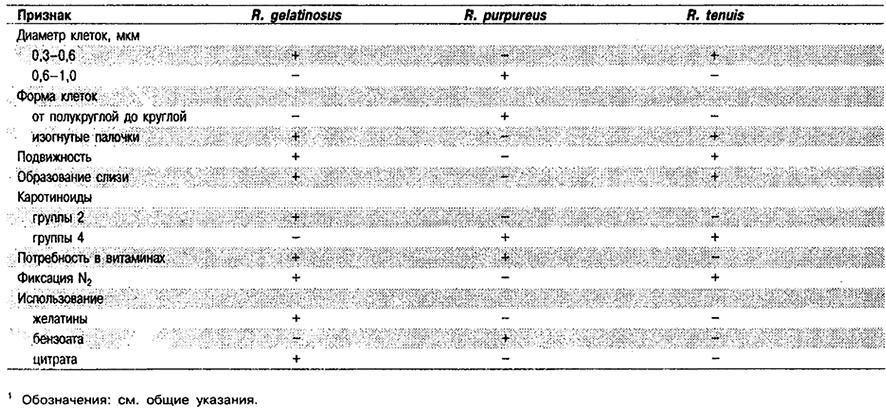
Род Rhodomicrobium
Клетки от овальных до удлиненно-овальных, диаметром 1,0–1,2 мкм, с полярным типом роста и характерным циклом вегетативного роста (рис. 10.6). Этот цикл включает образование подвижных клеток-швермеров с перитрихиальным жгутикованием и неподвижных материнских клеток, из которых вырастают нити длиной от равных материнской клетке до превышающих ее в несколько раз. На концах нитей возникают в виде круглых почек дочерние клетки, которые могут подвергаться дифференцировке того или иного типа. Способны образовывать устойчивые к высушиванию, полиэдральные экзоспоры. Внутрицитолазматические мембраны ламеллярного типа. Содержат бактериохлорофилл и каротиноиды. Растут предпочтительно фотогетеротрофно в анаэробных условиях на свету с использованием в качестве источников углерода и доноров электронов разнообразных органических субстратов. Могут использовать как доноры электронов для фотосинтеза молекулярный водород и сульфид, если они присутствуют в низких концентрациях. В факторах роста не нуждаются. Способны расти в темноте в микроаэробных или аэробных условиях. Оптимальная температура для роста 25–35°С; оптимальный рН 6,0.
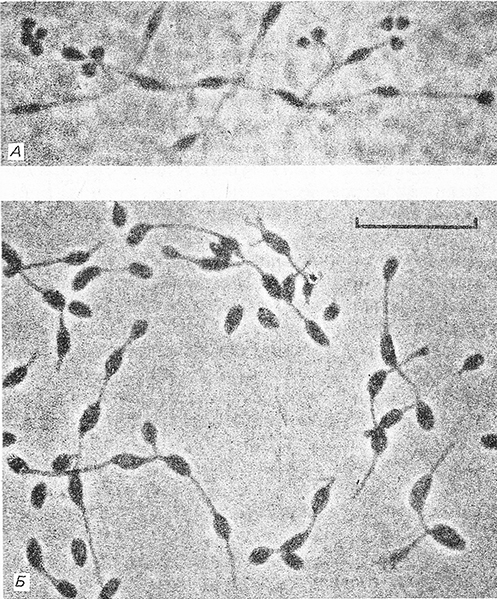
Рис. 10.6. Клетки Rhodomicrobium vannieli штамм DSM 163. А. Полиэдральные экзоспоры в виде почек на концах коротких нитей с обычно ветвящимся окончанием. Б. КЛетки с нитями и почками разных размеров. Фазовый контраст. Шкала – 10 мкм.
Типовой (и единственный) вид: Rhodomicrobium vannielii.
Признаки вида: Как для рода.
Род Rhodopila
Клетки от сферических до овальных, диаметром 1,6–1,8 мкм при определенных условиях роста, подвижные за счет, полярных жгутиков. Размножаются бинарным делением. Культуры пурпурно-красные. Внутриклеточные мембранные системы везикулярного типа; фотосинтетические пигменты – бактериохлорофилл α и каротиноиды. Растут предпочтительно, фотогетеротрофно в анаэробных условиях на свету. Чувствительны к кислороду, но растут в микроаэробных условиях в темноте. Оптимальная температура для роста 30–35°С; оптимальный рН, 4,8–5,0. Единственные, известные штаммы выделены из теплых кислых серных источников в Йеллоустонском национальном парке (шт. Вайоминг, США).
Признаки вида: Как для рода.
Род Rhodopseudomonas
Клетки палочковидные, 0,6–2,5 х 0,6–5,0 мкм, подвижные за счет жгутиков или неподвижные. Характерны полярный рост, почкование и ассиметричное клеточное деление. Внутриклеточные фотосинтетические мембраны в виде ламелл, подстилающих цитоплазматическую мембрану и расположенных параллельно ей. Фотосинтетические пигменты – бактериохлорофилл α или b и каротиноиды различных типов. Растут фотогетеротрофно с использованием ряда органических соединений в качестве источников углерода и доноров электронов. Фотоавтотрофный рост возможен в анаэробных условиях с использованием водорода, тиосульфата или сульфида в качестве донора электронов. Некоторые виды растут хемотрофно в микроаэробных или аэробных условиях. Нуждаются для роста в витаминах. Оптимальная температура 25–35°С.
Типовой вид: Rhodopseudomonas palustris.
Дифференциация видов рода Род Rhodopseudomonas: См. табл. 10.12.
Таблица 10.12. Дифференцирующие признаки видов рода Rhodopseudomonas 1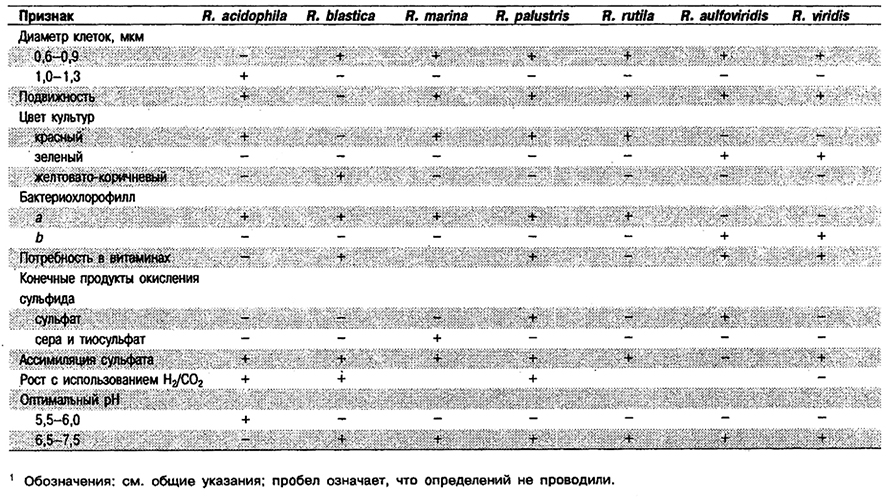
Род Rhodospirillum
Клетки спиральные, шириной 0,7–1,5 мкм, подвижные за счет полярных жгутиков. Размножаются бинарным делением. Внутриклеточные фотосинтетические мембраны в виде везикул или ламелл, расположенных параллельно цитоплазматической мембране или под острым углом к ней. Фотосинтезирующие пигменты – бактериохлорофилл α и каротиноиды спириллоксантиновой серии. Растут предпочтительно фотогетеротрофно в анаэробных условиях на свету, но способны также расти в микроаэробных или аэробных условиях в темноте. Нуждаются в факторах роста. Некоторые виды очень чувствительны к кислороду. В качестве донора электронов при фотосинтезе могут использовать молекулярный водород. Температура для роста 25–35°С; оптимальный рН 6,8–7,2.
Типовой вид: Rhodospirillum rubrum.
Типовой (и единственный) вид: Rhodopila globijormis.
Дифференциация видов рода Rhodospirillum: См. табл. 10.13.
Таблица 10.13. Дифференцирующие признаки видов рода Rhodospirillum 1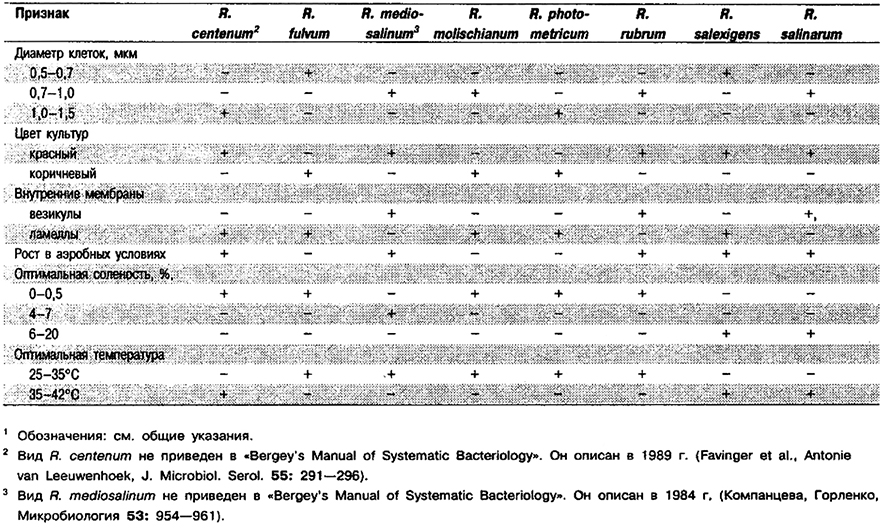
Подгруппа 4
Клетки растут за счет фотоассимиляции простых органических соединений; строгие анаэробы и фотогетеротрофы. Восстановленные соединения серы не используют. В качестве источников азота используют аммиак и молекулярный азот. Нуждаются для роста в витаминах. Внутриклеточные мембранные системы или хлоросомы отсутствуют. Содержат бактериохлорофилл g и каротиноиды. Клеточная стенка не содержит липополисахарид.
Дифференциация родов Подгруппы 4: Подгруппа 4 содержит два рода.
1. Подвижные за счет перитрихиальных жгутиков.
Heliobacillus
2. Безжгутиковые; способны к скользящему движению.
Heliobacterium
Род Heliobacillus
От редакции: Род Heliobacillus не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1987 г. (Beer-Romero, Gest, FEMS Microbiol. Lett. 41: 109–114) и включает только один вид Н. mobilis.
Палочки, 1,0 х 7–10 мкм, часто более длинные. Подвижные за счет перитрихиальных жгутиков. Культуры окрашены в зеленый цвет. Содержат бактериохлорофилл g и каротиноиды. Обингатные анаэробы и фототрофы; растут фотогетеротрофно с использованием ацетата, Пирувата, лактата и бутирата. Нуждаются для роста в витаминах. Оптимальная температура для роста 40–42°С; рН 7,0–7,2. Встречаются на рйсовых полях и во временно затопляемых почвах.
Типовой (и единственный вид): Heliobacillus mobilis.
Признаки вида: Как для рода.
Род Heliobacterium
Палочки, часто изогнутые, 1,0 х 4–10 мкм или длиннее. Способны к скользящему движению. Культуры окрашены в зеленый цвет. Содержат бактериохлорофилл g и каротиноиды. Облигатные анаэробы и фотогетеротрофы; растут предпочтительно с использованием органических кислот или сложных органических соединений в качестве источников углерода. Нуждаются для роста в витаминах. Оптимальная температура 35–42°С; рН 7,0–7,2. Выделены из поверхностного слоя почвы.
Типовой вид: Heliobacterium chlorum.
Признаки вида: Как для рода.
Подгруппа 5
Клетки способны расти с использованием сульфида или серы в качестве единственных доноров электронов для фотосинтетической ассимиляции СО2. На свету в присутствии сульфида происходит образование глобул серы снаружи клеток, никогда не внутри. Облигатные анаэробы и фототрофы; хорошо растут в фотоавтотрофных условиях. Простые органические соединения фотоассимилируют только в присутствии сульфида и бикарбоната. Могут нуждаться для роста в витамине В12. Культуры окрашены в зеленый цвет (бактериохлорофилл с и d) либо коричневый цвет (бактериохлорофилл е); см. табл. 10.1. Антенные бактериохлорофиллы локализованы в хлоросомах, которые подстилают цитоплазматическую мембрану и прикреплены к ней.
Дифференциация родов в Подгруппе 5: См. табл. 10.14.
Таблица 10.14. Дифференцирующие признаки родов Подгруппы 51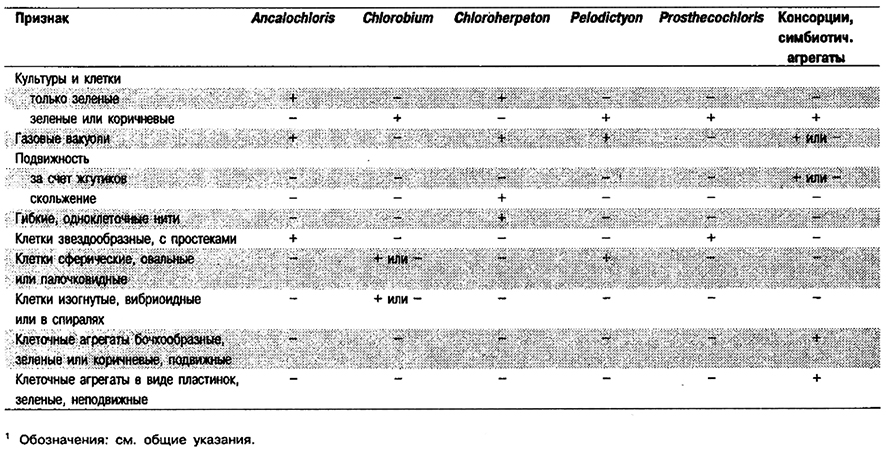
Род Ancalochloris
Клетки неправильной, звездчатой формы, диаметром 0,5–1,0 мкм без простек. Одна клетка может иметь до шести простек (рис. 10.7). Простеки различной длины, до 2 мкм, шириной 0,5–0,7 мкм у основания и 0,1 мкм на заостренных концах. Клетки размножаются путем неравного деления и могут оставаться соединенными, образуя цепочки неправильной формы, дающие начало микроколониям, в которых может быть до 30 клеток. Содержат газовые вакуоли. Неподвижные. Бактериохлорофиллы и каротиноиды локализованы в цитоплазматической мембране и хлоросомах. Облигатные анаэробы и фототрофы. Растут фотоавтотрофно с использованием сульфида и поли сульфидной серы в качестве доноров электронов для ассимиляции СО2; при этом снаружи клеток появляются глобулы серы. Оптимальный рН 6,8–7,0; оптимальная температура 20°С. Нуждаются для роста в витамине В12. Местом обитания служит бескислородный гиполимнион стратифицированных пресноводных озер или меромихтовых озер с низким содержанием сульфида, (0,1–1,0 мМ).

Рис. 10.7. Клетки Ancalochloris регfilievii из пробы воды гиполимниона микромиктового оз. Большой Кичиер, Россия. Электронная фотомикрография отдельной клетки, контрастированной фосфовольфрамовой кислотой (х З0 000).
Типовой (и единственный вид): Род Ancalochloris регfilievii.
Признаки вида: Как для рода.
Род Chlorobium
Клетки палочковидные, от прямых до изогнутых, овальные или спиральные, 0,3–1,1 х 0,4–3,0 мкм, хотя могут встречаться намного более длинные. Часто клетки остаются прикрепленными одна к другой, образуя нити или стрептококкоподобные цепочки; изогнутые палочковидные клетки могут образовывать плотно и свободно закрученные спирали. Неподвижные. Чистьте культуры окрашены либо в зеленый (травяной) либо в коричневый (шоколадно-коричневый) цвет; эти цвета можно также выявить при микроскопии в светлом поле. Фотосинтетические пигменты ; локализованы в цито плазматической мембране и хлоросомах, которые прикреплены к цитоплазматической мембране. Основные пигменты – бактериохлорофиллы с, d и е, кроме того в небольшом количестве присутствует бактериохлорофилл α. Основные каротиноиды – хлоробактин (у зеленых штаммов) или изорениератин (у коричневых штаммов). Облигатные анаэробы и фототрофы. При фотоавтотрофном росте используют сульфид или полисульфидную серу в качестве доноров электронов для ассимиляции СО2. При окислении сульфида снаружи клеток появляются глобулы серы; конечный продукт, окисления – сульфат. В восстановленных сульфидом средах могут использовать в качестве доноров электронов тиосульфат или молекулярный водород. В присутствии сульфида и бикарбоната фотоассимилируют ряд простых органических субстратов; наиболее часто используют ацетат и пропионат. Могут нуждаться для роста в витамине B12. Источником азота служит аммиак; многие штаммы фиксируют молекулярный азот. Температура для роста 20–35°С, Оптимальный рН 6,8–7,0. Местами обитания служат сульфиде одержащие ил и вода пресноводных, солоно- ватоводных и морских сред.
Типовой вид: Chlorobium limicola.
Дифференциация видов рода Chlorobium: См. табл. 10.15.
Таблица 10.15. Дифференцирующие признаки видов рода Chlorobium 1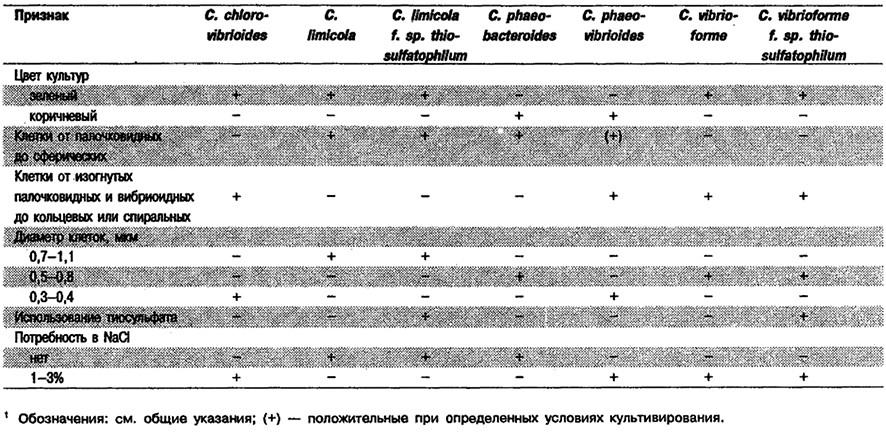
Род Chloroherpeton
Длинные палочки, 0,6–1,0 х 8–20 мкм, с закругленными концами. Клетки гибкие, прямые или изогнутые различным образом; угол изгиба до 180°. После деления клетки немедленно расходятся, причем никаких септ в каждой единице не наблюдается (одноклеточные). Могут содержать газовые вакуоли. Способны к скользящему движению со скоростью примерно 10 мкм • мин-1 при 20°С. Растут предпочтительно на поверхности плотных субстратов или в полужидких средах. При росте в жидкой среде образуют скопления и выделяют внеклеточную слизь. Культуры окрашены в зеленый цвет. Фотосинтетические пигменты локализованы в цитоплазматической мембране и хлоросомах. Основные пигменты – бактериохлорофилл с и γ-каротин. Облигатные анаэробы и фототрофы; способны к фотоавтотрофному росту с использованием сульфида и серы в качестве доноров электронов для ассимиляции СО2. При окислении сульфида снаружи клеток появляются глобулы серы, которая лишь медленно окисляется далее до сульфата. В присутствии сульфида и бикарбоната осуществляют фотоассимиляцию ацетата, пропионата, малата, сукцината или глугамата. Нуждаются для роста в витамине В12. Источником азота служит аммиак. Оптимальная температура 25°С; оптимальный рН 6,8–7,2. Обнаружены в сульфидсодержащих морских осадках на литорали, соляных маршах и приливных протоках с большим количеством гниющего растительного материала.
Типовой (и единственный вид); Chloroherpeton thalassium.
Признаки вида: Как для рода.
Род Pelodictyon
Клетки от палочковидных до овальных, 0,6–1,2 х 1,2–2,5 мкм. У двух видов клетки одиночные или в виде сетевидных агрегатов неправильной формы». Ветвление происходит в результате тройного деления. У двух других видов клетки одиночные или собраны в агрегаты сферической или неправильной формы. Содержат газовые вакуоли. Неподвижные. Культуры окрашены в зеленый или коричневый цвет. Фотосинтетические пигменты локализованы в цитоплазматической мембране и хлоросомах. Содержат бактериохлорофиллы c и d или е и, кроме того, небольшое количество бактериохлорофилла α. Основные каротиноиды – хлоробактин или изорениератин. Облнгатные анаэробы и фототрофы. При фотоавтотрофном росте используют сульфид и серу в качестве доноров электронов для ассимиляции СО2. При окислении сульфида снаружи клеток появляются глобулы серы. В присутствии сульфида и бикарбоната осуществляют фотоассимиляцию немногих простых органических веществ, в большинстве случаев ацетата или пропионата. Источником азота служит аммиак; могут ассимилировать молекулярный азот. Нуждаются для роста в витамине В12. Оптимальная температура 15–25°С; оптимальный рН 6,7– 7,1. Местами обитания служат содержащие H2S вода и ил пресноводных, солоноватоводных и морских сред. Эти организмы, для которых характерны газовые вакуоли, часто встречаются в планктоне и могут вызывать цветение в сульфидсодержащей стоячей воде озер.
Типовой вид: Pelodictyon clathratiforme.
Дифференциация видов рода Pelodictyon: См. табл. 10.16.
Таблица 10.16. Дифференцирующие признаки видов рода Pelodictyon 1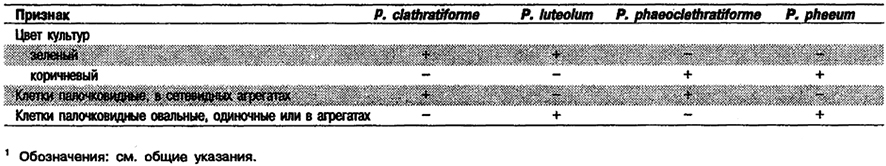
Род Prosthecochloris
Клетки сферические или в виде коротких палочек (перед делением), 0,5–0,7 х 0,5–1,2 мкм, каждая с 10–20 простеками шириной 0,1–0,17 и длиной 0,07–0,3 мкм, с закругленными концами. Клетки одиночные или в группах и цепочках. Газовых вакуолей не содержат. Неподвижные. Культуры окрашены в зеленый или коричневый цвет. Основные пигменты – бактериохлорофилл с или е и каротиноиды хлоробактин или изорениератин. Пигменты локализованы в цитоплазматической мембране и хлоросомах. Облигатные анаэробы и фототрофы. При фотоавтотрофном росте используют сульфид и серу в качестве доноров электронов для ассимиляции СО2. При окислении сульфида снаружи клеток появляются глобулы серы, которая может не полностью окисляться далее до сульфата. В присутствии сульфида и бикарбоната фотоассимилируют ацетат, пируват и некоторые другие субстраты. В качестве источника азота используют аммиак и могут фиксировать молекулярный азот. Нуждаются для роста в витамине В12. Диапазон температуры 20–30° С, оптимальный рН 6,7–7,0. Диапазон солености 0,2–7,0% NaCl, оптимальная концентрация NaCl 2%. Наиболее распространенные морские зеленые серобактерии. Местообитания – содержащие H2S ил и вода морских водоемов и озер с соленостью до 18% NaCl.
Типовой вид: Prosthecochloris aestuarii.
Дифференциация видов рода Prosthecochloris:
1. Культуры и клеточный материал окрашены в зеленый цвет; основные фотосинтетические пигменты – бактериохлорофилл с и хлоробактин.
P. aestuarii
2. Культуры и клеточный материал окрашены в коричневый цвет; основные фото синтетические пигменты – бактериохлорофилл d и изорениератин.
P. phaeoasteroidea
Консорции, симбиотические агрегаты
Консорции бактерий Подгруппы 5 (зеленые серобактерии) с бесцветными бактериями встречаются вместе с другими фототрофными бактериями Подгрупп 1 и 5 в иле и содержащей сульфид воде стоячих пресноводных прудов и стратифицированных пресноводных озер. Ни одна из консорций пока не получена в чистой культуре. Родовые и видовые обозначения, существующие в литературе, незаконны и приведены в табл. 10.17 в кавычках как лабораторные названия.
Таблица 10.17. Дифференцирующие признаки консорции,
симбиотических ассоциаций зеленых серных бактерий с бесцветными бактериями
(лабораторные названия заключены в кавычки)1
Подгруппа 6
Многоклеточные нити с однорядным расположением клеток, способные к скользящему движению. Факультативные аэробы; предпочтительно используют в фототрофном или хемотрофном метаболизме органические вещества. Содержат бактериохлорофиллы и различные каротиноиды.
Дифференциация родов в Подгруппе 6: См. табл. 10.18.
Таблица 10.18. Дифференцирующие признаки родов Подгруппы 61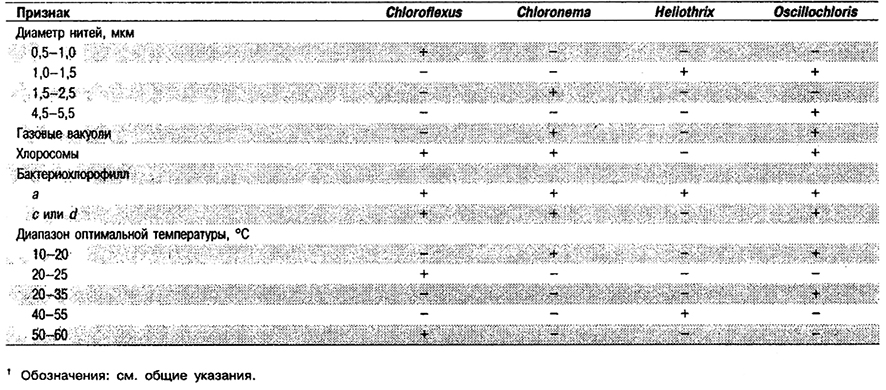
Род Chloroflexus
Многоклеточные нити неопределенной длины, разделенные септами, плохо различимыми в световой микроскоп; клетки 0,5–1,0 х 2–6 мкм. Иногда присутствует тонкий чехол. Способны к скользящему движению (0,01–0,04 мкм • с–1). Культуры, выращенные в анаэробных условиях на свету, окрашены в желтовато-зеленый цвет, в аэробных условиях – в оранжево-красный. Клетки, выращенные в анаэробных условиях на свету, содержат хлоросомы и бактериохлорофиллы с и α, а также каротиноиды – β- и γ-каротин. В анаэробных условиях растут преимущественно как фотогетеротрофы, а в аэробных условиях как хемогетеротрофы. В тех и других условиях используют следующие органические субстраты: дрожжевой экстракт, казаминовые кислоты, ацетат, глицерол, глюкозу, пируват или глутамат. Могут нуждаться для роста в фолиевой кислоте и тиамине. Источником азота служит аммиак. Оптимальный рН 7,6–8,4, Диапазон температуры для термофильных штаммов 40–66°С, оптимальная температура 52–60°С. Для мезофильных штаммов оптимальная температура 20–25°С, оптимальный рН 7,0–7,2. Термофильные штаммы встречаются по всему земному шару в горячих (40–68°С) источниках с нейтральной или щелочной реакцией воды; они образуют гелеобразные оранжево-красные маты, находящиеся обычно под тонким слоем цианобактерий. Мезофильные штаммы образуют морские микробные маты на мелководьях в соляных маршах или на освещенных участках донных осадков в пресноводных озерах.
Типовой (и единственный вид): Chloroflexus aurantiacus.
Признаки вида: Как для рода.
Род Chloronema
Многоклеточные, четко септированные нити различной длины, состоящие из клеток размерами 2,0–2,5 х 3,5–4,5 мкм и окруженные чехлом. Нити прямые или спиральные, способные к скользящему движению (~10 мкм • с–1). Клетки содержат крупные, расположенные центрально газовые вакуоли. Нити в суспензии и при наблюдении в светлопольный микроскоп желтовато-зелены е. Клетки содержат хлоросомы, бактериохлорофилл с или d и каротиноиды. Способны осуществлять аноксигенный фотосинтез и аэробный хемотрофный метаболизм. В чистой культуре не получены. Встречаются в области ниже хемоклина в стратифицированных пресноводных озерах вместе с другими видами фототрофных бактерий Подгрупп 1 и 5.
Типовой вид: Chloronema giganteum. Кроме типового вида описан (неполно) еще только один вид: С. spiroideum.
Признаки обоих видов: Как для рода.
Род Heliothrix
Многоклеточные, четко септированные нити различной длины, состоящие из клеток шириной примерно 1,5 мкм и длиной, значительно превышающей ширину. Возможно наличие тонкого чехла или чехол отсутствует. Подвижные за счет скольжения. Фототрофные культуры окрашены в ярко-оранжевый цвет. Клетки содержат только бактериохлорофилл α и насыщены каротиноидами; хлоросомы и дополнительные бактериохлорофиллы отсутствуют. Аэротолерантные; метаболизм главным образом фотогетеротрофный, с использованием ацетата. В чистой культуре не получены; возможно получить совместную культуру с аэробной хемогетеротрофной бактерией Isosphaera pallida. Диапазон температуры 35–56°С с оптимумом в пределах 40–55°С. Встречаются в некоторых горячих источниках в шт. Орегон, США, где образуют оранжевые маты толщиной несколько миллиметров; такой мат обычно покрыт зеленоватым слоем цианобактерий.
Типовой (и единственный) вид: Heliothrix oregonensis.
Признаки вида: Как для рода.
Род Oscillochloris
Многоклеточные нити неопределенной длины с поперечными септами, видимыми в световой микроскоп; клетки шириной 1,0–1,4 мкм или 4,5–5,5 мкм. Клетки не имеют чехла, но содержат газовые вакуоли. По Граму окрашиваются положительно или отрицательно. Нити гибкие и подвижные за счет скольжения, желтовато-зеленые или зеленые. В анаэробных условиях клетки содержат хлоросомы, бактериохлорофилл с и каротиноиды. Растут фототрофно в анаэробных условиях и могут обладать способностью расти хемогетеротрофно в аэробных условиях. Температура для роста 10– 20°С; оптимальный рН 7,5. В чистой культуре не получены. Встречаются в матах на поверхности сульфидсодержащего ила пресноводных прудов и озер. Совместно с другими видами фототрофных бактерий Подгрупп 1 и 5 вызывают цветение.
Типовой вид: Oscillochloris chrysea.
Дифференциация видов рода Oscillochloris:
1. Нити шириной 4,4–5,5 мкм и длиной 3,5– 7,0 мкм; желтовато-зеленые в анаэробных и аэробных условиях.
О. chrysea
2. Нити шириной 1,0–1,4 и длиной 2,3–3,8 мкм; клеточные суспензии темно-зеленые в анаэробных условиях на свету и оранжевые в аэробных условиях.
О. trichoides
Подгруппа 7
Клетки растут хемогетеротрофно в аэробных условиях; в анаэробных условиях на свету роста нет. Метаболизм преимущественно дыхательного типа. Содержат бактериохлорофилл а и каротиноиды.
Подгруппа 7 содержит только один род.
Род Erythrobacter
Клетки от овальных до палочковидных, шириной 0,5 мкм н длиной 1,0–5,0 мкм. Грамотрицательные. Размножаются бинарным делением. Подвижные за счет субполярных жгутиков. Культуры и колонии оранжевые или розовые. Содержат бактериохлорофилл а и каротиноиды. Растут в аэробных условиях как хемогетеротрофы. В анаэробных условиях на свету не растут. Источниками углерода служат ацетат, пируват, бутират, глутамат и глюкоза. Нуждаются для роста в витаминах. Оптимальная температура дня роста 25–30°С; диапазон рН 7,0–8,0; оптимальная соленость 1,7–3,5% NaCl. Встречаются в кислородсодержащих морских местообитаниях, преимущественно на водорослях.
Типовой (и единственный) вид: Erythrobacter longus.
Признаки вида: Как для рода.
