Род Flavimonas
От редакции: Род Flavimonas не приведен в «Вегgey's Manual of Systematic Bacteriology». Этот род описан в 1987 г, (Holmes et al., Int. J. Syst. Bac- terioi. 37: 247-250) и включает единственный вид F. oryzihabitans.
Палочки с параллельными сторонами и закругленными концами. Внутриклеточные гранулы поли-β-гмдроксибутирата обычно отсутствуют. Простек или чехлов не образуют. Покоящиеся стадии неизвестны. Грамотрицательные. Аэробы; метаболизм чисто дыхательного типа. Растут при температуре 18-42°С. На плотной среде окрашены в цвета от бледно- до темно-желтого. Колонии обычно округлые (диаметром 1 мкм), слабо выпуклые, гладкие (отдельные штаммы образуют морщинистые колонии), блестящие, с цельным краем. Каталазоположительные; оксидазоотрицательные. Агар не разлагают. Хемоорганотрофы. Используют сахара. Обнаружены в обычной природной среде; по-видимому, сапрофиты или комменсалы человека и гомойотермных животных, для которых иногда могут быть патогенными.
Типовой (и единственный) вид: Flavimonas oryzihabitans.
Признаки вида: Клетки подвижные за счет единственного полярного жгутика или неподвижные. На агаре с тирозином некоторые штаммы продуцируют темно-коричневый, диффундирующий меланиноподобный пигмент. Гемолиз не осуществляют. Флуоресцирующих пигментов на среде В Кинга не выделяют. Растут при комнатной температуре и 37°С, но не при 5 и 42°С. В тесте на окисление-брожение с глюкозой обнаруживают окислительную реакцию. Гидролизуют трибутирин и Твин-20, но не желатину. Твин-80 не гидролизуют. Казеин не разлагают. Опалесценции при росте на агаре с лецитовителлином не наблюдается. Нитрат или нитрит не восстанавливают. Индол и H2S не образуют (тесты с бумагой, пропитанной ацетатом свинца, и на среде с тремя сахарами и железом). Растут за счет использования β-гидроксибутирата, но не образуют при этом липидных включений, а также за счет цетримида. Растут на агаре Макконки. Крахмал и эскулин не гидролизуют. В среде Кристенсена с цитратом вызывают подщелочение. Используют цитрат (агар Симмонса). Глюконат не окисляют. Аргининдезимидазу, аргининдигидролазу, лизиндекарбоксилазу и орнитиндекарбоксилазу, а также 3-кетолакгозу не образуют. Селенит не восстанавливают. По фенилаланиндезаминазе отрицательные. β-Галактозидазу не образуют. В аэробных условиях на среде с солью аммония образуют кислоту из D-глюкозы, L-арабинозы, этанола, D-фруктозы, глицерола, маннитола, L-рамнозы, сорбитола, трегалозы и D-ксилозы. Кислоту из адонитола, дульцитола, лактозы, рафинозы и салицина на среде с солью аммония в аэробных условиях не образуют. В среде на основе пептонной воды ни кислоты, ни газа из D-глюкозы не образуют.
Род Flavobacterium
Палочки с параллельными сторонами и закругленными концами, как правило 0,5 х 1,0-3,0 мкм. Внутриклеточные гранулы поли-β-гидроксибутирата отсутствуют. Эндоспор не образуют. Грамотрицательные. Неподвижные. Скользящее движение не осуществляют и не распространяются. Аэробы; метаболизм чисто дыхательного типа. Изоляты из природной среды растут при 37°С. При росте на плотных средах обычно окрашены (в цвета от желтого до оранжевого), но есть и непигментированные штаммы. Колонии просвечивающие (в отдельных случаях непрозрачные), округлые (диаметром 1-2 мм), выпуклые или слабо выпуклые, гладкие и блестящие, с цельным краем. Каталазо-, оксидаэо- и фосфатазоположительные. Агар не разлагают. Хемоорганотрофы. В средах с низкой концентрацией пептона образуют из углеводов кислоту, но не газ. Широко распространены в почве и воде; обнаружены также в сыром мясе, молоке и других пищевых продуктах, а также в больницах и у человека (в клиническом материале).
Типовой вид: Flavobacterium aquatile.
Дифференциация видов рода Flavobacterium: См. табл. 4.24.
Таблица 4.24. Дифференцирующие признаки видов рода Flavobacterium 1,2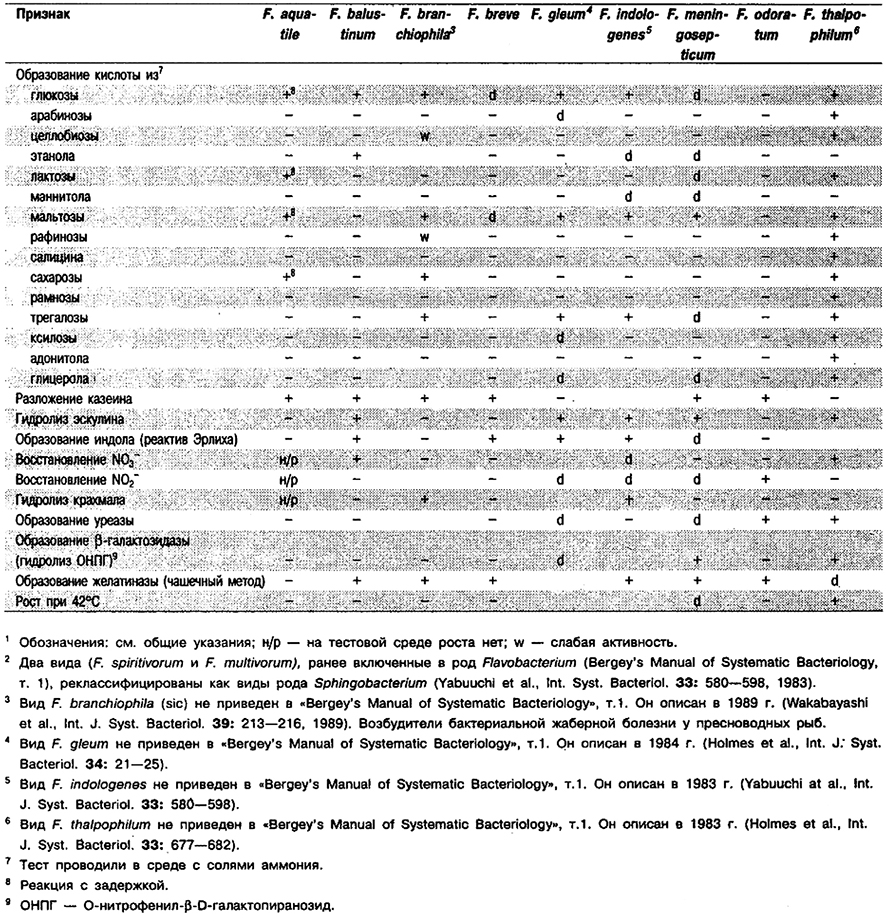
Род Francisella
Клетки палочковидные, 0,2 х 0,2-0,7 мкм (Francisella tularensis) или 0,7-1,7 мкм (Francisella novicicla) в период активного роста при культивировании на подходящей среде, впоследствии клетки плеоморфные. Грамотрицательные, окраска слабая. Неподвижные; облигатные аэробы. На кровяном агаре с глюкозой и цистеином образуют гладкие колонии серого цвета, достигающие максимальных размеров через 2-4 сут культивирования и окруженные характерной зоной просветления. Слабо каталазоположительные. Оксндазоотрицательные. Углеводы обычно разлагают медленно, с образованием кислоты, но не газа. Цистеин (или цистин) либо необходим для роста (F. tularensis), либо сильно стимулирует рост (F. novicida). Образуют H2S. Типовой вид в отличие от других бактерий содержит относительно большие количества длинноцепочечных насыщенных и мононенасыщенных жирных кислот С20-26, а также 3-гидроксигексадекановую, 2-гидроксидекановую и. 3-гидроксиоктадекановую кислоты. F. tularensis – возбудитель туляремии у человека и животных. F. novicida вызывает экспериментальные инфекции у лабораторных животных.
Типовой вид: Francisella tularensis.
Дифференциация видов рода Francisella: См. табл. 4.25.
Таблица 4.25. Дифференцирующие признаки видов рода Francisella 1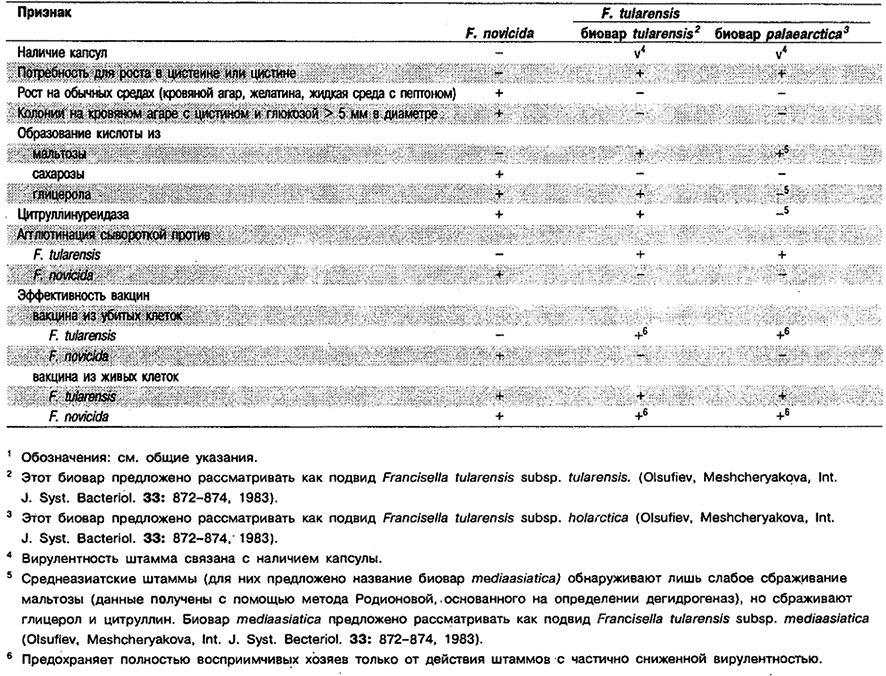
Род Frateuria
Правильные прямые палочки, 0,5-0,7 x 0,7-3,5 мкм, одиночные или в парах. Грамотрицательные. Подвижные за счет полярных жгутиков или неподвижные. Облигатные аэробы. Оптимальная температура для роста 25-30°С. На агаризованной среде с маннитолом, дрожжевым экстрактом и пептоном (МУР) колонии окрашены в цвета от желтого до оранжевого. На агаризованной среде с глюкозой, дрожжевым экстрактом и СаСО3 (GYC) большинство штаммов образует характерный коричневый водорастворимый пигмент. Оксидазоотрицательные. Растут прн рН 3,6. Нитрат не восстанавливают. Крахмал и желатину не гидролизуют. Образуют H2S. Хемоорганотрофы. Образуют кислоту при использовании этанола и многих других источников углерода, При росте в среде с D-глюкозой или D-ксилозой рН падает ниже 4,0. Из D-глюкозы образуют 2-кето- и 2,5-дикетоглюконовую кислоты, но не 5-кетоглюконовую кислоту. В факторах роста не нуждаются. Выделены из Lilium auratum, а также из плодов Rubus parvifolius (малина) в Японии.
Типовой (и единственный) вид: Frateuria aurantia.
Признаки вида: Как для рода.
Род Gluconobacter
Клетки от эллипсоидных до палочковидных, 0,5-1,0 х 2,6-4,2 мкм, одиночные и/или в парах, редко в цепочках. Могут присутствовать увеличенные клетки неправильной формы (инволюционные формы). Эндоспор не образуют. Грамотрицательные (в немногих случаях грамвариабельные). Подвижные или неподвижные; если подвижные, то за счет 3-8 полярных жгутиков, редко одного жгутика. Считались облигатными аэробами с метаболизмом чисто дыхательного типа, использующими кислород как конечный акцептор электронов, однако недавно установлена их способность восстанавливать тиосульфат до H2S, В среде, содержащей сорбитол и тиосульфат, образуют H2S (метод с бумажными полосками). Колонии бледные. Оптимальная температура для роста 25-З0°С, при 37°С не растут. Оптимальный диапазон рН 5,5-6,0; большинство штаммов растет при рН 3,6, Обладают высокой каталазной активностью; оксидазоотрипательные. Нитрат не восстанавливают; желатину не разжижают; индол не образуют. Хемоорганотрофы. Окисляют этанол до уксусной кислоты. Ацетат и лактат до СО2 и Н2О не окисляют. В присутствии многоатомных спиртов происходит интенсивный кетогенез. Образуют кислоту из D-глюкозы и D-ксилозы. Все штаммы образуют из D-глюкозы 2-кетоглюконовую кислоту, а большинство также 5-кетоглюконовую кислоту. В среде с крахмалом не растут и кислоту не образуют. Представители Gluconobacter встречаются на цветах, в садовой почве, пекарских дрожжах, на медоносных пчелах, фруктах, в сидре, пиве, внне, винном уксусе, южноафриканском пиве банту, пальмовом соке и безалкогольных напитках. О некоторых штаммах известно, что они вызывают «розовую болезнь» ананасов и гниль яблок и груш, однако правильность идентификации этих штаммов как представителей Gluconobacter подвергается сомнению (см. Micales et al., Int. J. Syst. BacterioL 35: 79-85; 1985). В отличие от штаммов Acetobacter, для которых предпочтительны среды с повышенным содержанием спирта, для штаммов Gluconobacter предпочтительны среды с повышенным содержанием Сахаров,
Типовой вид: Gluconobacter oxydans.
Дифференциация видов рода Gluconobacter: См. табл. 4.26.
Таблица 4.26. Дифференцирующие признаки трех видов рода Gluconobacter 1 
Род Halomonas
Клетки палочковидные или плеоморфные, первые обычно 0,6-0,8 x 1,6-1,9 мкм. В некоторых условиях образуют удлиненные гибкие нити. Спор не образуют Грамотрицательные. Подвижные или неподвижные; подвижность за счет нескольких латеральных или полярных жгутиков без чехлов. Метаболизм преимущественно дыхательного типа с использованием кислорода в качестве конечного акцептора электронов. Некоторые штаммы могут расти в анаэробных условиях с использованием нитрата. Возможен рост в анаэробных условиях в среде с глюкозой в отсутствие нитрата, что указывает на некоторую способность к брожению, однако для анаэробного роста в средах с другими углеводами или аминокислотами нуждаются в нитрате. Колонии от белых до желтых, но не красные. Каталазо- и оксидазоположительные. Большинство штаммов восстанавливает нитрат до нитрита. Хемоорганотрофы. В качестве единственных источников углерода в минеральной среде могут использовать углеводы, аминокислоты и некоторые многоатомные спирты, а в качестве единственного источника азота – сульфат аммония. Галотолерантные; способны расти при концентрациях NaCl от 0,05-20% (масса/объем) до более высоких. Выделены из оборудования для добычи грубой соли из рапы, выпариваемой под солнцем, из соленого озера в Антарктиде, а также из эстуария в шт. Нью-Гэмпшир.
Типовой вид; Halomonas elongata.
Дифференциация видов рода Halomonas: См. табл. 4.27.
Таблица 4.27. Дифференцирующие признаки видов рода Halomonas 1 
Род Hydrogenophaga
От редакции: Род Hydrogenophaga не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1989 г. (Willems et al,, Int. J. Syst, Bacteriol. 39: 319-333) и включает виды, ранее отнесенные к роду Pseudomonas как P. flava, Р. pseudoflava, P. palleronli, P. taeniosp trails и «Р. carboxy doflava».
Палочки, 0,3-0,6 х 0,6-5,5 мкм, от прямых до слегка изогнутых, одиночные или в парах. Подвижные за счет одного, редко двух, полярных или субполярных жгутиков. Грамотрицательные. Оксидазоположительные. По признаку каталазной активности виды различаются. Образуют недиффундирующнй желтый пигмент. Аэробы и факультативные Н2-автотрофы. Метаболизируют углеводы путем окисления с использованием кислорода в качестве конечного акцептора электронов. Некоторые виды (Н. pseudoflava и Н. taeniospiralis) способны к анаэробному нитратному дыханию, которое сопровождается денитрификацией. Хорошо растут на средах с органическими кислотами, аминокислотами или пептоном; углеводы используют слабее. Содержат циклопропановую жирную кислоту (С17:цик) и 3-гидроксиоктановую кислоту (3-ОН-С8:0), а также в некоторых случаях 3-гидроксидекановую кислоту (3-ОН-С10:0). Не содержат 2-гидроксилированных жирных кислот. Основной хинон – убихинон Q-8. Единственные или доминирующие полиамины – 2-гидроксипутресцин и путресцин, присутствующие примерно в эквимолярных количествах.
Типовой вид: Hydrogenophaga flava.
Дифференциация видов рода Hydrogenophaga: См. табл. 4.28.
Таблица 4.28. Дифференцирующие признаки видов рода Hydrogenophaga 1,2 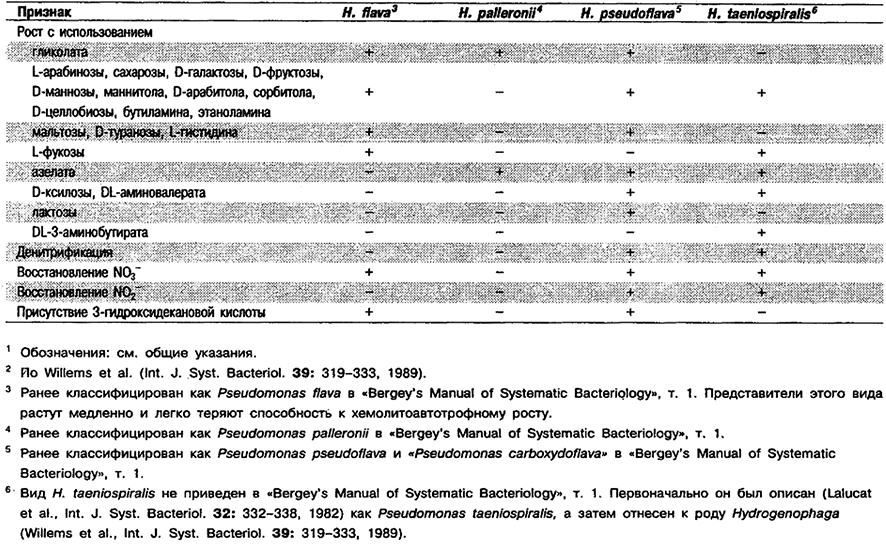
Род Janthinobacterium
Палочки, 0,8-1,2 x 2,5-6,0 мкм, с закругленными концами, иногда слегка изогнутые. Располагаются одиночно, но в отдельных случаях в виде коротких цепочек. Капсулы четко не выражены, хотя иногда образуют межклеточную слизь. Покоящиеся стадии неизвестны. Грамотрицательные; окрашивание иногда полосами или биполярное; присутствуют липидные включения. Подвижные за счет одного полярного плюс обычно от одного до четырех субполярных или латеральных жгутиков. Облигатные аэробы. Колонии на плотных средах слегка выпуклые, округлые, фиолетового цвета; на поверхности жидкой среды образуют фиолетовое кольцо по стенке сосуда. Оптимальная температура для роста 25°С, минимальная – 2°C, максимальная – 35°С. Оптимальный диапазон рН 7-8, при рН ниже 5 не растут. В среде с 6% и более NaCl не растут. Хемоорганотрофы; метаболизм чисто дыхательного типа с использованием кислорода в качестве конечного акцептора электронов. Из глюкозы и ряда других углеводов образуют кислоту, но не газ. Окисляют лактат до СО2. Тест Ковача (Kovacs, 1956) обычно обнаруживает оксидазную активность, хотя присутствие фиолетового пигмента может мешать интерпретации результатов. Каталазоположительные; индол не образуют; реакция Фогеса-Проскауэра отрицательная. Восстанавливают нитрат и нитрит, иногда с видимым выделением газа. Образуют аммиак из пептона. Содержат фосфатазную активность. Растут на обычных средах с пептоном; арилсульфатазу не образуют. Используют цитрат и аммоний в качестве единственных источников углерода и азота, при этом рост быстрый. В факторах роста не нуждаются. Устойчивы к бензилпенициллину (10 мкг/мл) и к вибриостагическому агенту О/129 (2,4-диамино-6,7-диизопропилптеридин, 30 мкг/диск). Почвенные и водные организмы, распространены в поясе умеренного климата. Иногда вызывают порчу пищевых продуктов.
Типовой (и единственный) вид: Janthinobacterium lividum.
Признаки вида: Как для рода.
Род Kingella
Прямые палочки длиной 1,0 мкм, с закругленными или квадратно очерченными концами, в парах и иногда коротких цепочках. Эндоспор не образуют. Грамотрицательные, но с более или менее выраженной устойчивостью к обесцвечиванию. По результатам обычных тестов клетки неподвижные, однако могут иметь фимбрии (пили) и обладать способностью к движению рывками (twitching motility). Аэробы или факультативные анаэробы; лучше всего растут в аэробных условиях, но способны также к слабому росту в анаэробных условиях на кровяном агаре. Оптимальная температура для роста 33-37°С. Колонии на кровяном агаре бывают двух типов; 1) распространяющиеся по поверхности, эродирующие агар; этот тип ассоциирован со способностью к движению рывками, наличием фимбрий и компетентностью к трансформации и 2) гладкие, выпуклые колонии, клетки в которых неподвижные, не имеют фимбрий и не компетентные. Оксидазоположительные (в тесте с тетраметилфенилендиамином; с диметиловым реагентом реакция может быть слабой или отрицательной). Каталазоотрицательные. Свернутую сыворотку не разжижают. Уреазу не образуют. Фенилаланиндезаминазная активность слабая или отсутствует. Хемоорганотрофы. Сбраживают глюкозу и ограниченное число других углеводов с образованием кислоты, но не газа. Чувствительны к пенициллину. Встречаются у человека в слизистой оболочке верхних дыхательных путей.
Типовой вид: Kingella kingae.
Дифференциация видов рода Kingella: См. табл. 4.29.
Таблица 4.29. Дифференцирующие признаки видов рода Kingella 1 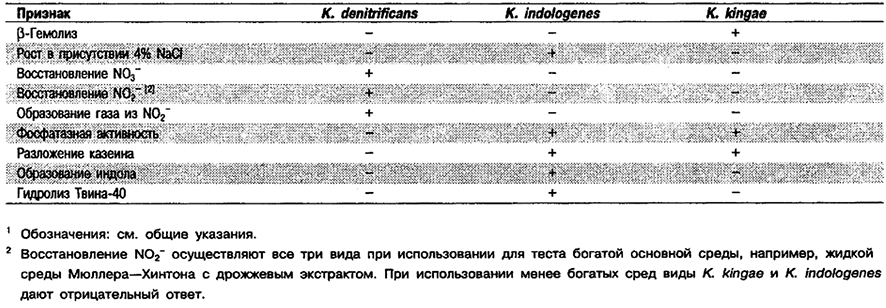
Род Lampropedia
Пластинки из округлых или почти кубических клеток, расположенных в виде квадратных табличек по 16-64 клетки, иногда разделенных на пары или тетрады. Деление клеток происходит синхронно в пластинке, поочередно в двух плоскостях. Клетки, образующие табличку, окружены сложной, структурированной оболочкой. Каждая клетка окружена клеточной стенкой грамотрицательного типа. Содержат хорошо заметные внутриклеточные гранулы поли-β-гидроксибутирата. Жгутиков не имеют. Во время активного роста наблюдается движение рывками небольших групп клеток. Облигатные аэробы; метаболизм чисто дыхательного типа с использованием кислорода в качестве конечного акцептора электронов. Рост как на плотной, так и в жидкой среде наблюдается в виде тонкой, гидрофобной, распространяющейся пленки. Непигментированные. Оптимальная температура для роста 30°С. Оптимальное значение рН 7,0. Оксидазо- и каталазоположительные, Хемоорганотрофы. Источниками энергии служат только интермедиаты цикла трккарбоновых кислот. Углеводы, спирты, глюкозиды и жирные кислоты не используют. Единственными источниками азота могут служить аммонийные соли некоторых аминокислот. Возможна потребность в витаминах для роста. Экологическая ниша неизвестна, но наблюдения и способ выделения указывают на среду, богатую органическими веществами.
Типовой (и единственный) вид: Lampropedia hyalina.
Признаки вида: Клетки 1,0-1,5 x 1,0-2,5 мкм. Морфологические признаки как для рода. В качестве единственного источника энергии используют пируват, лахтат, бутират, фумарат, малат или сукцинат (а также ацетат в присутствии пирувата в каталитической концентрации). В качестве единственного источника азота используют NH4Cl, аланин, аргинин или тирозин. Нуждаются для роста в биотине и тиамине. Диапазон температуры для роста 10-35°С, оптимальная температура 30°С. Диапазон рН для роста 6,0-8,6, оптимальное значение – 7,0.
Род Legionella
Палочки, 0,3-0,9 x 2-20 мкм или более. Эндоспор, микроцист и капсул не образуют. Некислотоустойчивые. Грамотрицательные. Подвижные за счет одного, двух или большего числа прямых или изогнутых, полярных или латеральных жгутиков; у отдельных штаммов клетки неподвижные. Аэробы. Нуждаются для роста в L-цистеингидрохлорнде и солях железа. Результаты теста на оксидазу отрицательные или слабо положительные. Нитрат не восстанавливают. Уреазу не образуют. Разжижают желатину. В составе клеточной стенки доминируют жирные кислоты с разветвленной цепью.
Хемоорганотрофы; в качестве источников углерода и энергии используют аминокислоты. Углеводы ие сбраживают и не окисляют. Выделены с поверхности воды, из ила, а также термально загрязненных озер и источников. Сообщений о выделении из почвы или животных не имеется. Патогенны для человека: служат возбудителями пневмонии («болезнь легионеров») и лихорадки Понтиака.
Типовой вид: Legionella pneumophila.
Дифференциация видов рода Legionella: В настоящее время род Legionella включает 30 видов и 47 серологически различающихся групп (Thacker et at., J. Clin. Microbiol. 27: 1831-1834; 1989; Bornstein et al., Res. Microbiol. 140: 541–552, 1989). Идентификацию проводят по серологическим признакам, позволяющим определить наиболее часто встречающиеся виды и серовары, однако антисыворотки для многих видов не поступают в продажу. Существуют тесты, потенциально пригодные для разграничения видов Legionella, однако они еще не опробованы на всех видах или на достаточном числе штаммов многих видов, кроме того, многие из этих тестов не подходят для постоянного использования в лаборатории (Vasey et al., J. AppL Bacteriol. 65: 339-345, 1988: Fox, Brown, J. Clin. Microbiol. 27: 1952-1955, 1989). В связи с этим, идентификация большинства видов Legionella должна производиться в справочной лаборатории.
Род Marinobacter
От редакции: Род Marinobacter не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1992 г. (Gauthier et al., Int. J. Syst. Bacteriol. 42: 568–576) и включает единственный вид М. hydrocarbonoclasticus.
Клетки палочковидные, 0,3-0,6 x 2,0-3,0 мкм. Поверхность клеток, выращенных в минеральной среде с эйкозаном, покрыта многочисленными вздутиями. Грамотрицательные, Спор не образуют. В средах, содержащих 0,2-1 М NaCl, подвижные за счет единственного, не покрытого чехлом полярного жгутика. При росте в средах с более высокой или низкой концентрацией NaCl жгутиков не имеют. Молодые колонии на агаризованной среде окрашены в белый цвет, после 48-часовой инкубации в розовато-бе же вый. Растут при температуре 10-4 5°С (мезофилы), оптимальная температура 32°С. Экстремально галотолерантные; могут расти при концентрациях NaCl от 0,08 до 3,5 М. Облигатно нуждаются в ионах натрия. Аэробы; метаболизм не бродильного типа. Растут в анаэробных условиях с использованием нитрата или сукцината, цитрата либо ацетата, но не глюкозы; осуществляют денитрификацию с выделением N2. Обладают оксидазной, цитохромоксидазной, каталазной, твиназной и лецитиназной активностями. Используют как единственные источники углерода ацетат, бутират, капронат, сукцинат, фумарат, адипинат, DL-лактат и цитрат, но не углеводы и аминокислоты (за исключением L-пролина и L- глутамата). Разлагают разнообразные алифатические и ароматические углеводороды; при росте в средах с углеводородами образуют недиализируемый биоэмульгатор. Устойчивы к новобиоцину, тетрациклину, олеандомицину, стафиломицину и вибриостатическому агенту 0/129; чувствительны к пенициллину G, канамицину, стрептомицину, хлорамфениколу, эритромицину, цефалоридину, гентамицину и налидиксовой кислоте.
Типовой (и единственный) вид: Marinobacter hydrocarbonoclasticus.
Признаки вида: Как для рода.
Род Marinomonas
От редакции: Род Marinomonas не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1983 г. (Landschoot, De Ley, J. Gen. Microbiol. 129: 3057–3074, Int. J. Syst. Bacteriol. 34: 91–92, 1984) и включает два вида: М. communis (прежнее название – Alteromonas communis) и М. vaga (прежнее название – Alteromonas vaga). М. communis описан также в Группе 2.
Прямые или изогнутые палочки, 0,7–1,5 х 1,8–3,0 мкм. Поли-β-гидроксибутират не накапливают. Микроцист и эндоспор не образуют. Грамотрицательные. Подвижные за счет одного полярного жгутика. Хемоорганотрофы; метаболизм дыхательного, не бродильного, типа. Универсальным конечным акцептором электронов служит кислород. Нитрат до нитрита не восстанавливают и денитрификацию не осуществляют. Ни один из штаммов не обладает конститутивной аргининдигидролазной системой. Все штаммы нуждаются в морской воде как основе среды для роста. Используют D-сорбитол, DL-малат, α-кетоглутарат, D-маннозу, D-фруктозу, сукцинат, фумарат, глицерол и м-гидроксибензоат. Внеклеточную желатиназу и липазу не образуют.
Типовой вид: Marinomonas communis.
Дифференциация видов рода Marinomonas: См. табл. 4.30.
Таблица 4.30. Дифференциация видов рода Marinomonas 1 
Род Mesophilobacter
От редакции: Род Mesophilobacter не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1989 г. (Nishimuraet al., Int. J. Syst. Bacteriol. 39: 378–381) и включает единственный вид М. marinus,
Плеоморфные палочки, 0,5–0,6 х 1,0–2,0 мкм. В молодых культурах встречаются налитые (утолщенные) формы (0,8–1,0 x 1,5–3,0 мкм) и удлиненные клетки (15 мкм или более), а в старых культурах кокковндные клетки (диаметром 0,6–0,9 мкм). Грамотрицательные; капсул не имеют. Неподвижные. Каталазо- и оксидазоположительные. Умеренные галофилы. Оптимальная температура для роста 33–37°С. Растут при 5°С, но не при 0°С. Аэробы; метаболизм чисто дыхательного типа. Устойчивы к пенициллину. Выделены из морской воды.
Типовой (и единственный) вид: Mesophilobacter marinus.
Признаки вида: Колонии на МПА обычно округлые, иногда неправильной формы, гладкие, слегка выпуклые, с цельным краем, блестящие, непрозрачные, бледного желтовато-коричневого цвета, В МП Б рост умеренный, в лакмусовом молочке – со слабым осадкообразованием и без реакции, По признакам образования индола, наличия желатиназы и уреазы штаммы различаются. Проба с метиловым красным положительная; реакция Фогеса-Проскауэра отрицательная. Фенилаланиндезаминазная активность отсутствует. Гемолиз не осуществляют. Восстанавливают нитрат до нитрита. При использовании D-рибозы, D-глюкозы, D-фруктозы, D-маннитола и целлобиозы образуют кислоту, но не газ. Ассимилируют D-рибозу, D-глюкозу, D-фруктозу, мальтозу, сахарозу, трегалозу, ацетат, цитрат, сукцинат, фумарат, малат и глюконат. Растут в присутствии 7% NaCl. Основной убихинон – Q–8. В составе клеточных жирных кислот преобладают нераз- ветвленные кислоты С16:0, C18:1 и C16:1.
Род Methylobacillus
От редакции: Род Methylobacillus не приведен в «Bergey's Manual of Systematic Bacteriology». Этот
род описан в 1986 г, (Urakami, Komagata, Int. J. Syst, Bacteriol, 36: 502-511) и включает единственный вид М. glycogenes.
Короткие грамотрицательные палочки, подвижные за счет одного полярного жгутика или неподвижные. Облигатные метилотрофы; растут за счет использования метанола или других одноуглеродных соединений, но не метана. По способности использовать D-фруктозу штаммы различаются. Строгие аэробы; метаболизм дыхательного типа. В составе жирных кислот преобладают неразветвленная насыщенная кислота С16:0 и ненасыщенная кислота C16:1. Основной хинон – Q-8, минорные – Q-7 и Q-9.
Типовой (и единственный) вид; Methylobacillus glycogenes.
Признаки вида. Неспорообразующие палочки с закругленными концами, 0,3-0,5 x 0,8-2,0 мкм, одиночные, редко в парах, подвижные за счет одного полярного жгутика; иногда неподвижные. Капсул не образуют. Гранул поли-β-гидроксибутирата в клетках не накапливают. В МПБ и жидкой среде с пептоном не растут. На агаризованной среде с метанолом колонии блестящие, гладкие, приподнятые, с цельным краем, от белых до светло-желтых, диаметром 1-3 мм через 3 сут инкубации при 30°С. Водорастворимые пигменты не образуют. Восстанавливают нитрат до нитрита. Проба с метиловым красным и реакция Фогеса-Проскауэра отрицательные. Индолотрицательные. H2S и аммиак не образуют. Желатину и крахмал не гидролизуют. Денитрификацию не осуществляют. Кислоту из D-глюкозы и D-фруктозы не образуют. В качестве единственного источника углерода используют метанол, но не метан. Облигатные метилотрофы. Не используют L-арабинозу, D-глюкозу, D-ксилозу, D-маннозу, галактозу, мальтозу, глицерол, растворимый крахмал, янтарную, лимонную и уксусную кислоты, а также этанол и водород. По способности использовать D-фруктозу и метиламин штаммы различаются. В витаминах и аминокислотах для роста не нуждаются, однако несколько штаммов нуждаются в тиамине. В качестве единственного источника азота используют аммиак, мочевину и нитрат. Уреазоотрицательные. Оксидазоотрицательные. Большинство штаммов образует каталазу. Аэробы. Метаболизм чисто дыхательного типа. Хорошо растут при рН от 6,0 до 8,0. Все штаммы растут при 30°С и примерно 50% штаммов – при 42°С. В присутствии 3% NaCl большинство штаммов не растет.
Род Methylobacterium
От редакции: Род Methylobacterium не приведен в «Bergey's Manual of Systematic Bacteriology», хотя включен в «Одобренный список названий бактерий» (1980). Типовой вид – М. organophilum. В 1983 г. к роду добавлены три вида: М. rhodinum (прежнее название – Pseudomonas rhodos), М. radiotolerans (прежнее название – Pseudomonas га~ diora) и М. mesophilicum (прежнее название Pseudomonas mesophilica), и внесены уточнения в его описание (Green, Bousfield, Int. J. Syst. Bacteriol. 33: 875-877). В 1984 г. описан род Protomonas, включающий один вид Р, extorquens (прежнее название – Pseudomonas extorquens) (Urakami, Komagata, Int. J. Syst. Bacteriol. 34: 188-201), однако в 1985 г. представители Protomonas классифицированы и перенесены в род Methylobacterium (Bousfield, Green, Int. J. Syst. Bacteriol. 35: 209). Таким образом, P. extorquens стал M. extorquens. В 1988 г. к роду Methylobacterium отнесены еще три вида: М. rhodesianum, М. zatmanii и М. fujisawaense (Greenetal., Int. J, Syst. Bacteriol., 38: 124-127).
Палочки, 0,8-1,0 x 1,0-8,0 мкм, одиночные или иногда в розетках. В некоторых случаях клетки разветвленные или плеоморфные. Подвижные за счет единственного полярного, субполярного или латерального жгутика, хотя у некоторых штаммов подвижность слабо выражена. Клетки часто содержат крупные включения, окрашивающиеся Суданом, и иногда гранулы волютина. Грамотрицательные, однако многие штаммы грамвариабельные; для репрезентативных штаммов характерна многослойная клеточная стенка и тип цитратсинтазы, свойственный грамотрицательным бактериям. На МПА большинство штаммов растет медленно, а некоторые не растут совсем. На глицерол-пептонном агаре колонии диаметром 1-3 мм, от бледно-розовых до ярких оранжево-красных; на метанол-солевом агаре колонии более однородные, бледно-розового цвета. Пигмент нерастворимый, вероятно каротиноидный. При культивировании в жидкой среде без перемешивания растут в виде розового кольца или пленки на поверхности среды. Облигатные аэробы; каталазо- и оксидазоположительные (активность последнего фермента часто низкая). Хемоорганотрофы, факультативные метилотрофы и в ряде случаев факультативные метан отрофы. Способность некоторых штаммов использовать метан в качестве единственного источника углерода и энергии легко утрачивается, если их не поддерживать на неорганической среде в атмосфере метана. Репрезентативные штаммы ассимилируют одноуглеродные соединения по гомоизоцитратному пути; при росте на сложных органических средах действует полный цикл трикарбоновых кислот. Выделены из почвы, пыли, пресной воды, озерных осадков, с поверхности листьев, из клубеньков, зерен риса, воздуха и больничной среды. Оптимальная температура для роста 25-30°С.
Типовой вид: Methylobacterium organophilum.
Дифференциация видов рода Methylobacterium: См. табл. 4.31.
Таблица 4.31. Дифференцирующие признаки видов рода Methylobacterium 1,2 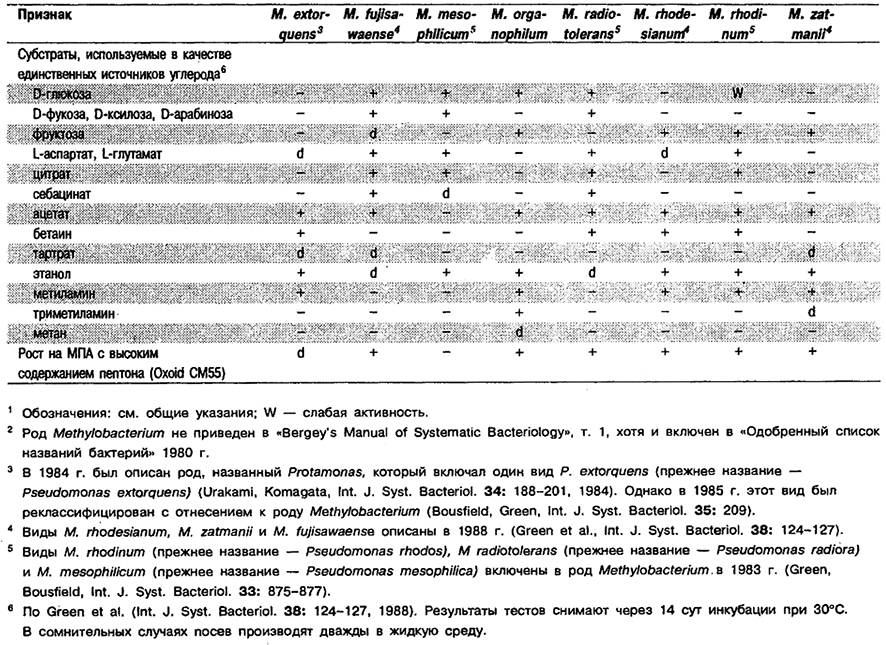
Род Methylococcus
Клетки сферические, обычно в парах. Неподвижные. Имеют покоящуюся стадию в виде цист. Грамотрицательные. Аэробы; метаболизм чисто дыхательного типа с использованием кислорода в качестве конечного акцептора электронов. Как единственные источники углерода и энергии известны только метан, метанол и формальдегид. В органических факторах роста не нуждаются.
Типовой вид: Methylococcus capsulatus.
Признаки вида: Кокки диаметром 1,0 мкм. Образуют капсулы. Поли-β-гидроксибутират не образуют. Клетки непигментированные. В качестве источника азота нитрит предпочтительнее солей аммония и гидролизата казеина. В органических факторах роста не нуждаются, хотя рост колоний на агаризованной среде стимулируют экстракты сложного состава, такие как гидролизат казеина. Оптимальная температура 37°С. Растут в диапазоне температуры 30-50°С, но не при 55°С. Основной путь ассимиляции углерода – рибулозомонофосфатный (содержат 3-гексулозофосфатсинтазу). Цикл трикарбоновых кислот неполный (не содержат 2-кетоглутаратдегидрогеназу). Обладают только NAD-зависимой изоцитратдегидрогеназой. Способны к автотрофной фиксации СО2 (содержат рибулозобисфосфаткарбоксилазу). Могут обладать или не обладают нитрогеназной активностью. Доминирующие жирные кислоты – C16.
Род Methylomonas
Прямые, изогнутые или разветвленные, но не спиральные палочки, 0,5–1,0 х 1,0–4,0 мкм. Подвижные за счет единственного полярного жгутика. Чехлы или про стеки не обнаружены. Один вид образует инсты. Грамотрицательные. Аэробы; метаболизм чисто дыхательного типа с использованием кислорода в качестве конечного акцептора электронов. Как соединения, используемые в качестве единственных источников углерода и энергии, известны только метан, метанол и формальдегид. В органических факторах роста не нуждаются. Одному виду для роста необходим NaCl. Диапазон температуры 20–35°С.
Типовой вид: Methylomonas methanica.
Дифференциация видов рода Methylomonas: См. табл. 4.32.
Таблица 4.32. Дифференцирующие признаки видов рода Methylomonas 1 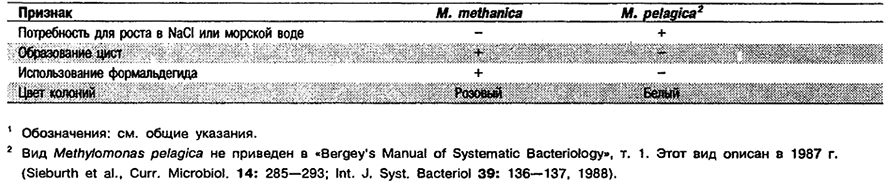
Род Methylophaga
От редакции: Род Methylophaga не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1985 г. (Janvier et al,, Int. J. Syst, Bacteriol. 35: 131–139) и включает два вида: М. marina и М. thalassica.
Короткие прямые палочки диаметром 0,2 мкм. Подвижные за счет единственного полярного жгутика. Характерно обширное периплазматическое пространство (20-30 км). Клетки могут разрушаться в результате осмотического шока после отмывки 0,5 М NaCl. Облигатные аэробы и умеренные галофилы. Ауксотрофы по витамину В12. Не растут в среде с пептоном и дрожжевым экстрактом, содержащей (или не содержащей) NaCl. Кроме фруктозы используют в качестве субстратов для роста только одноуглеродные соединения, такие как метанол и метиламин, которые метаболизируют по рибулозомонофосфатному пути. С использованием метана не растут. Выделены из морской среды.
Типовой вид: Methylophaga marina.
Дифференциация видов рода Methylophaga: См. табл. 4.33.
Таблица 4.33. Дифференцирующие признаки видов рода Methylophaga 1,2 
Род Methylophilus
От редакции: Род Methylophilus не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1987 г. (Jenkins et al., Int. J. Syst. Bacteriol. 37: 446-448) и включает один вид М. methylotrophus.
Клетки, выращенные на минеральной агаризованной среде с метанолом или в жидкой минеральной среде с метанолом, имеют форму прямых или слегка изогнутых палочек, обычно 0,3-0,6 х 0,8-1,5 мкм, расположенных одиночно или парами. Грамотрицательные, однако часто плохо обесцвечиваются. Подвижные за счет полярных жгутиков или неподвижные. Эндоспор не образуют. Внутриклеточных включений не содержат. Чехлов или простек не обнаружено. Капсул не образуют, однако клетки некоторых штаммов выделяют слизь. Колонии на агаризованной среде с метанолом и минеральными солями (2 сут инкубации при 30°С или 37°С) округлые, диаметром 1-2 мм, с цельным краем, выпуклые, от просвечивающих до непрозрачных. Пиоцианин и флуоресцеин не образуют. На МПА или в МП Б (2 сут при 30 или 37°С) не растут или рост крайне слабый. На кровяном агаре не растут или растут очень слабо; гемолиз не осуществляют. Оптимальная температура для роста 30-37°С, при 4 и 45°С не растут. Оптимальный диапазон рН 6,5-7,2. Аэробы. Метаболизм дыхательного типа; кислотообразование при использовании глюкозы очень незначительное или отсутствует. Все штаммы окисляют метанол в качестве единственного источника углерода и энергии. Кроме того, единственными источниками углерода и энергии служат некоторые другие углеродные соединения: метиламины, формиат, глюкоза и фруктоза. Потребности в питательных веществах простые; источниками азота служат нитрат и соли аммония. Каталазо- и оксидазоположитсльные. В составе жирных кислот преобладают негидроксилированные неразветвленные насыщенные и мононенасыщенные кислоты С16:0 и С16:1. Основные изопреноидные хиноны – убихиноны с восемью изопреновыми единицами (Q-8). Выделены из активного ила, ила, речной и прудовой воды.
Типовой (и единственный) вид: Methylophilus теthylotrophus.
Признаки вида: Колонии на агаризованной среде с метанолом и минеральными солями серовато-белые. Клетки подвижные за счет единственного жгутика. Кроме роста на среде с метанолом в качестве единственного источника углерода и энергии хороший рост отмечен на среде с глюкозой и в некоторых случаях с метиламинами как единственными источниками углерода и энергии. По способности использовать в этом качестве фруктозу штаммы различаются. Могут давать слабый рост, варьирующий в зависимости от штамма, на средах с лактозой, сахарозой, D-рибозой, D-ксилозой, этанолом, пропанолом, бутанолом, ацетатом и формиатом. Кислоту из глюкозы не образуют. Ацетоин образуют или не образуют (реакция Фогеса-Проскауэра). Гидролизуют Твин-20, -40 и -60, но не -80. Образуют уреазу и лейцинариламидазу. Синтез фосфатазы слабый и различается у разных штаммов, Сульфатазу и H2S не образуют. Желатину не разжижают. Внеклеточную ДНКазу и РНКазу не продуцируют. Восстанавливают 2,3,5-трифенилтетразолийхлорид (0,01%), В присутствии теллурита калия (0,01%) или NaCl (5%) не растут. Устойчивы к пенициллину и олеацдомицину; чувствительны к налидиксовой кислоте, стрептомицину и многим другим антибиотикам.
Род Methylovorus
От редакции: Род Methylovorus не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1991 г. (Govorukliina, Trotsenko, Int. J. Syst. Bacteriol. 41: 158–162) и включает единственный вид М. glucosotrophus.
Клетки, выращенные на среде с метанолом и минеральными солями, имеют форму .прямых или слегка изогнутых палочек, обычно 0,5–0,6 x 1,0–1,3 мкм, расположенных одиночно или парами. Грамотрицательные. Эндоспор не образуют. Не содержат внутриклеточных мембранных структур. Чехлов или простек не обнаружено. Капсул не образуют, однако некоторые штаммы могут образовывать слизь. Колонии на агаризованной среде с метанолом и минеральными солями (2 сут при 30°С) округлые, диаметром 1–2 мм, с цельным краем, выпуклые, от просвечивающих до непрозрачных, розового, кремового или молочного цвета. Пиоцианин и флуоресцеин не образуют. Размножаются бинарным делением. В жидкой среде клеточные агрегаты не формируют, пигменты не образуют. На МПА или в МП Б при 30–37°С не растут или рост крайне слабый; в атмосфере СН4 + О2 или Н2 + СО2 + О2 не растут. Оптимальный диапазон рН для роста 7,0–7,2, температуры – 35–37°С. Облигатные аэробы; метаболизм дыхательного типа. В качестве источников углерода и энергии используют метанол и глюкозу. Помимо этого некоторые штаммы способны слабо расти с использованием метилированных аминов, инулина и бетаина. Источниками азота служат нитраты, соли аммония, метилированные амины, глутамат и пептон. Ацетоин, H2S, индол и NH3 в тестовых средах не образуют. Молоко не гидролизуют. Уреазо-, каталазо- и оксидазоположитсльные. По пероксидазе вариабельные. Аргининдигидролазная активность отсутствует. Могут гидролизовать крахмал, но не целлюлозу, желатину или Твин-80. Из глюкозы образуют кислоту (но не газ).
Все штаммы ассимилируют углерод метанола по рибулозомонофосфатному пути и аммиак в глутаматном цикле (глутаматсинтаза и глутаминсинтетаза). α-Кетоглутаратдегидрогеназа и ферменты глиоксилатного шунта отсутствуют. В составе жирных кислот преобладают негидроксилированные неразветвленные насыщенные и мононенасышенные кислоты С16:0 и С 16:1ω7. Разветвленных жирных кислот С17 не содержат. Основные фосфолипиды – фосфатидилэтаноламин и фосфатидилглицерол. Некоторые штаммы содержат также ди фосфатидилглицерол. Выделены из активного ила, ила, почвы и прудовой воды.
Типовой (и единственный) вид: Methylovorus glucosotrophus.
Признаки вида: Как для рода.
Род Moraxella
Палочки (подрод Moraxella) или кокки (подрод Branhamella). Палочки часто очень короткие и толстые, приближающиеся по форме к коккам (1,0–1,5 x 1,5–2,5 мкм), обычно в парах и коротких цепочках (деление происходит в одной плоскости). В культурах часто наблюдается изменчивость по признакам размеров и формы клеток, а также образования нитей или цепочек; плеоморфизм усиливается в отсутствие кислорода и при температуре выше оптимальной. Кокки обычно более мелкие (диаметром 0,6–1,0 мкм) и расположены одиночно или парами, где соприкасающиеся стороны клеток уплощены (разные плоскости деления); деление происходит в двух взаимоперпендикулярных плоскостях, иногда с образованием тетрад. Клетки могут быть окружены капсулой. Грамотрицательные, но часто с более или менее выраженной устойчивостью к обесцвечиванию. Жгутики отсутствуют. Как палочковидные, так и кокковидные бактерии могут иметь фимбрии. Способностью к движению путем плавания не обладают, но у некоторых видов палочковидных бактерий описано движение рывками по твердой поверхности. Аэробы, но отдельные штаммы могут слабо расти в анаэробных условиях. Для большинства видов (за исключением М. [M.] osloensis) характерны сложные потребности в питательных веществах, но конкретные ростовые потребности неизвестны. Оптимальная температура для роста 33–35°С. Обычно каталазоположительные. Хемоорганотрофы. При использовании углеводов кислотообразования не происходит. Обычно высокочувствительны к пенициллину. Паразитические организмы; встречаются на слизистых оболочках у человека и пойкилотермных животных.
Типовой вид: Moraxella (Moraxella) lacunata,
Дифференциация видов рода Moraxella: См. табл. 4,34.
Таблица 4.34. Дифференцирующие признаки видов рода Moraxella 1 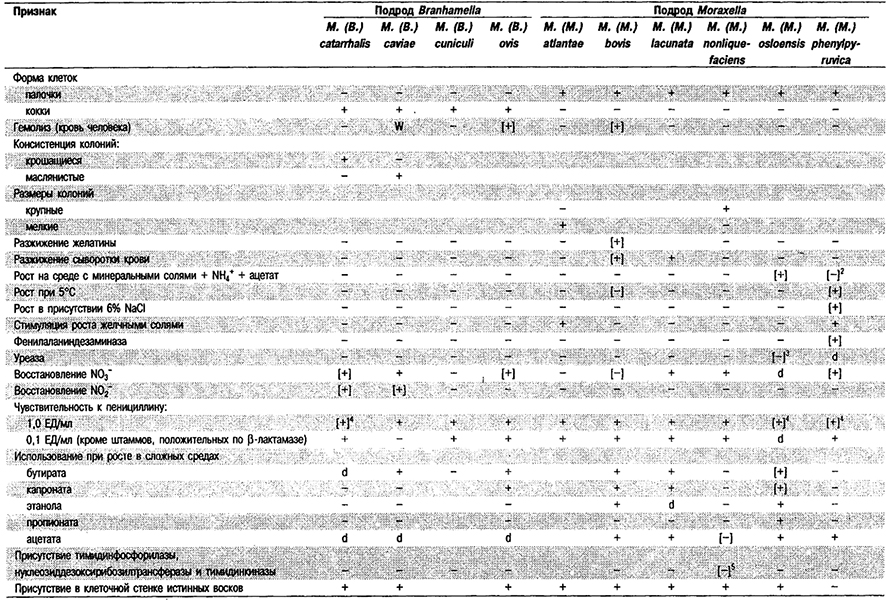

Род Morococcus
От редакции. Род Morococcus не приведен в «Bergey's Manual of Systematic Bacteriology)». Этот род описан в 1981 г. (Long et al, Int. J. Syst. Bacteriol. 31: 294-301) и включает единственный вид М. cerebrosus.
Кокки диаметром <1 мкм, образующие плотно упакованные, похожие на тутовые ягоды агрегаты из 10-20 клеток. Неподвижные и неспорообразующие, Поли-β-гидроксибутират не образуют. Аэробы. В химически сложных факторах роста не нуждаются. Колонии на сахарозном агаре окрашиваются в черный цвет при реакции с иодом. Диапазон температуры для роста 23–42°С, рН 5,5–9,0. Каталазо- и оксцдазоположительные. Восстанавливают нитрат. Образуют H2S. Образуют кислоту из углеводов. Первоначально выделены у человека из абсцесса мозга.
Типовой (и единственный) вид; Morococcus cerebrosus.
Признаки вида: При проведении теста на окисление–брожение (O/F-тест) по Хью-Лейфсону могут давать слабую реакцию в обеих пробирках. Образуют кислоту из глюкозы, фруктозы, сахарозы и мальтозы, но не из арабинозы, рибозы, ксилозы, рамнозы, галактозы, маннозы, сорбозы, салицина, целлобиозы, лактозы, мелибиозы, трегалозы, мелецитозы, рафинозы, декстрина, инулина или крахмала. Восстанавливают лакмусовое молочко. Образуют H2S из цистеина. Проба с метиловым красным положительная. По ДНКазе и орнитиндекар- боксилазе положительные; по лецитиназе, фосфатазе, фенилаланиндезаминазе и уреазе отрицательные. Индол не образуют. Цитрат и малонат не окисляют. Крахмал, эскулин, желатину, казеин и Твин-80 не гидролизуют. Патогенны для мышей.
Отличия М, cerebrosus от видов Neisseria: См. табл. 4.35.
Таблица 4.35. Дифференцирующие признаки видов
рода Neisseria и Morococcus cerebrosus 1,2 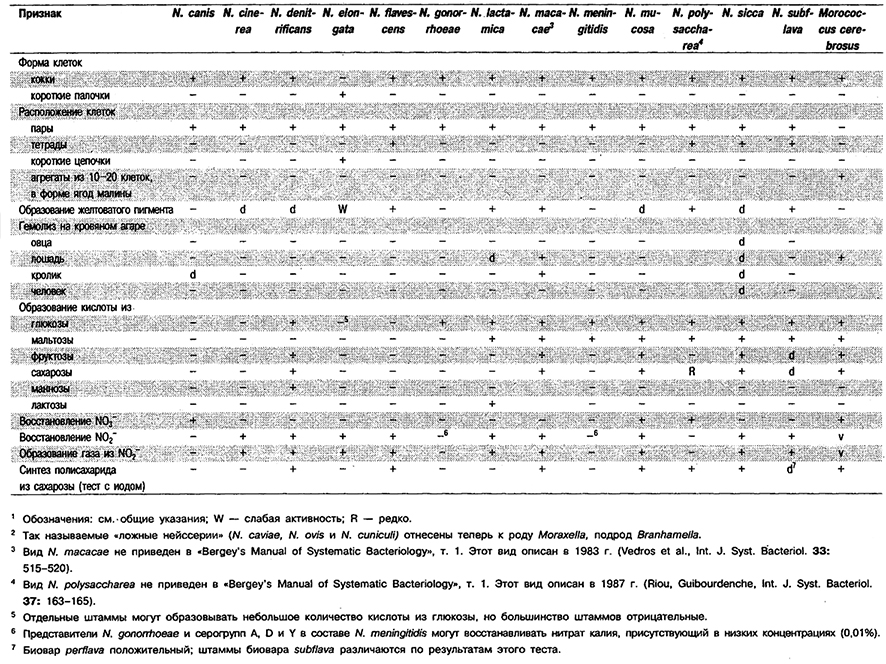
Род Neisseria
Кокки диаметром 0,6-1,0 мкм, одиночные или, чаще, в парах (соприкасающиеся стороны клеток при этом уплощены); один вид (N. elongata) составляет исключение – относящиеся к нему бактерии представляют собой короткие палочки шириной 0,5 мкм, часто в виде диплобацилл или в коротких цепочках. Деление у видов кокковидных бактерий происходит в двух взаимноперпендикулярных плоскостях, иногда с образованием тетрад. Могут иметь капсулы и фимбрии (пили). Эндоспоры отсутствуют. Грамотрицательные, но с более или менее выраженной устойчивостью к обесцвечиванию. Способностью к плаванию не обладают и жгутиков не имеют. Аэробы. Представители некоторых видов образуют зеленовато-желтый каротиноидный пигмент. Некоторые виды высокотребовательны в отношении питательных веществ и обладают гемолитической активностью. Оптимальная температура 35-37°С. Оксндазоположительные. Каталазоположительные, за исключением N. elongata. Образуют карбоангидразу. Все представители, за исключением N. gonorrhoeae и N. can is, восстанавливают нитрит. Хемоорганотрофы. Некоторые виды сахаролитические. Присутствуют в микрофлоре слизистых оболочек млекопитающих. Некоторые виды вызывают первичные инфекции у человека,
Типовой вид: Neisseria gonorrhoeae.
Дифференциация видов рода Neisseria: См. табл. 4.35.
Род Oceanospirillum: Oceanospirillum kriegii и Oceanospirillum jannaschii
От редакции: Эти виды не приведены в «Bergey's Manual of Systematic Bacteriology». Они описаны в 1984 г. (Bowdich et al., Curr. Microbiol. 10: 221-230; Int. J. Syst. Bacteriol, 34: 503, 1984) и отличаются от других видов рода Oceanospirillum тем, что относящиеся к ним бактерии имеют палочковидную форму, а не спиральную (см. также Pot et al., Int. J. Syst. Bacteriol. 39: 23-34, 1989).
Признаки Oceanospirillum kriegii: Прямые палочки, 0,8-1,2 x 2,6–3,6 мкм. Подвижные за счет одного единственного полярного жгутика. Накапливают поли-β-гидроксибутнрат в качестве запасного внутриклеточного продукта. Аэробы; метаболизм чисто дыхательного типа. Для роста необходимы ионы натрия или среда на основе морской воды. Растут при 20-35°С, но не при 4 или 40°С. Используют 29-33 органических соединения, в том числе D-глюкозу, D-фруктозу и маннитол, но не другие пентозы, гексозы или дисахариды. В качестве единственного источника углерода и энергии используют ацетат, сукцинат, фумарат, DL-малат, пируват, α-кетоглутарат, n-гидроксибензоат, L-аланин, D-аланин, L-пролин, этанол, пропанол и хиннат. Не используют изобутанол, сахарат, гликолат, W-аминовалерат, L-тирозин, гистамин и саркозин. Образуют внеклеточную липазу. Нитрат до нитрита не восстанавливают.
Признаки Oceanospirillum jannaschii: Прямые палочки, 1,0–1,4 x 2,4–3,2 мкм. Подвижные за счет 1–2 жгутиков на одном из полюсов. Накапливают полигндроксибутрат в качестве запасного внутриклеточного продукта. Аэробы; метаболизм чисто дыхательного типа. Для роста необходимы ионы натрия или среда на основе морской воды. Растут при 20-30°С, но не при 4 или 35°С, Используют 39-46 органических соединений, но не используют углеводы. В качестве единственного источника углерода и энергии используют ацетат, сукцинат, фумарат, DL-малат, пируват, L-аланин, D-аланин, L-пролин, W-аминовалерат, гистамин, саркозин, этанол, пропанол и изобутанол. Не используют сахарат, гликолат, хиннат, L-тирозин, и n-гидроксифенилбензоат. Внеклеточную липазу не образуют. Восстанавливают нитрат до нитрита.
Род Ochrobactrum
От редакции: Род Ochrobactrum не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1988 г. (Holmes et al., Int. J. Syst. Bacteriol. 38: 406-416) и включает единственный вид О. anthropi.
Палочки с параллельными сторонами и закругленными концами, как правило одиночные. Покоящиеся стадии неизвестны. Грамотрицательные. Подвижные за счет лернтрихиальных жгутиков. Облигатные аэробы; метаболизм чисто дыхательного типа с использованием кислорода в качестве конечного акцептора электронов. Оптимальная температура для роста 20-37°С. Колонии на МПА неокрашенные. Оксидазо- и каталазоположительные. Индол отрицательные. Эскулин, желатину и ДНК не гидролизуют. Хемоорганотрофы; используют в качестве источников углерода разнообразные аминокислоты, органические кислоты и углеводы. Встречаются в клиническом материале. Прежде были известны как «CDC Group Vd».
Типовой (и единственный) вид: Ochrobactrum anthropi.
Признаки вида: Растут на агаре Макконки. В аэробных условиях на среде с солью аммония образуют кислоту из глюкозы, арабинозы, этанола, фруктозы, рамнозы и ксилозы. Растут за счет использования β-гидроксибутирата. Отрицательные по следующим признакам: образование пигментов, окисление глюконата, лизин и орнитиндекарбоксилазная активности, образование газа и кислоты в среде на основе пептонной воды, восстановление селенита, разложение казеина, внеклеточная ДНКаза, образование кислоты из рафинозы в аэробных условиях в среде с солью аммония, аргининдезимидазная активность, гидролиз Твина-20 и -80, образование на агаре с тирозином коричневого меланиноподобного пигмента, накопление в клетках липидов при росте с использованием β-гидроксибутирата, флуоресценция на среде В Кинга, рост при 5°С, образование 3-кетолактозы и лецитиназы, гидролиз крахмала, образование кислоты из лактозы (10%).
Род Oligella
От редакции: Род Oligella не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1987 г. (Rossau et al, Int. J. Syst. Bacteriol. 37: 198-210) и включает два вида: О. urethralis (прежнее название – Moraxella urethralis) и О. ureolytica (прежде эти бактерии были известны как «СDC Group IVe»).
Мелкие палочки, обычно длиной <1 мкм, часто в парах. Клетки не имеют утолщенного вида, типичного для моракселл. Капсул и спор не образуют. В большинстве неподвижные, однако для некоторых штаммов О. ureolytica характерно перитрихиальное жгутикование. Аэробы. Хемоорганотрофы, умеренно требовательные в отношении питательных веществ. Растут на МПА, однако добавление, например, дрожжевого автолизата, сыворотки крови или крови стимулирует рост. На кровяном агаре колонии развиваются довольно медленно и по цвету более ярко-белые, чем у всех признанных видов Moraxella. Пигментов и пахучих соединений не образуют. Гемолиз не осуществляют. Биохимически довольно инертные; в качестве единственных источников углерода используют лишь небольшое число органических кислот и аминокислот. Углеводы не сбраживают и не окисляют. Оксидазоположительные и обычно каталазоположительные. Индол и H2S не образуют Желатину не гидролизуют. Выделены главным образом из мочеполовых путей человека. О патогенности неизвестно; возможно, слабо патогенные.
Типовой вид: Oligella urethralis.
Дифференциация видов рода Oligella: См. табл. 4.36.
Таблица 4.36. Дифференцирующие признаки видов рода Oligella 1,2 
Род Paracoccus
Клетки сферические (диаметром 0,5–0,9 мкм) или в форме коротких палочек (длиной 0,9–1,2 мкм), одиночные, в парах или группах. Содержат внутриклеточные гранулы поли-β-гидроксибутирата. Покоящиеся стадии неизвестно Грамотрицательные. Неподвижные. Аэробы; метаболизм чисто дыхательного типа; могут расти в анаэробных условиях в присутствии нитрата, нитрита или оксида азота (NO) как конечного акцептора электронов. В анаэробных условиях восстанавливают нитрат до оксцда азота (NO) и молекулярного азота. Представители одного из видов могут расти автотрофно в присутствии Н2 + СО2 или гетеротрофно за счет использования в качестве единственного источника углерода одного из разнообразных органических соединений; этот вид не галофильный. Представители другого вида не способны к автотрофному росту, но галофилы. Оптимальная температура 25-30°С. Оксндазо- и каталазоположительные. Встречаются в почве и, предположительно, в природной и искусственной рапе.
Типовой вид: Paracoccus denitrificans.
Дифференциация видов рода Paracoccus: См. табл. 4.37.
Таблица 4.37. Дифференцирующие признаки видов рода Paracoccus 1

Род Phenylobacterium
От редакции: Род Phenylobacterium не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1985 г. (Makkar Lingens et al. Int. J. Syst. Bacteriol. 35: 26-39) и включает единственный вид P. immobile.
Палочки, кокковидные палочки или кокки, 0,7-1,0 x 1,0-2,0 мкм, одиночные, в парах или коротких цепочках, У некоторых штаммов более или менее выражено образование комков клеток. В старых культурах могут присутствовать плеоморфные формы: длинные палочки (1,0 x 2,0-4,0 мкм), соединенные нитями длинные цепочки (10-50 мкм) и эллиптические формы. Клетки неподвижные. Чехлов и простек не образуют. Покоящиеся стадии неизвестны. Капсулы при окраске не выявляются, но гибкая капсула имеется. Грамотрицательные. Рост на агаре слабый, колонии мелкие (диаметром 1-2 мм через 2-3 нед инкубации). Колонии гладкие, выпуклые, влажные, с блестящей поверхностью и цельным краем, легко эмульгирующие в растворе соли либо скорее шероховатые, сухие и не эмульгирующие в растворе соли. В жидкой среде образуют в небольшом количестве зеленовато-желтый флуоресцирующий пигмент. Хемоорганотрофы; метаболизм дыхательного, никогда не бродильного, типа. Осмотически чувствительные. Нуждаются в витамине B12 как факторе роста. Характерны очень специфические потребности в питательных веществах. Хороший рост наблюдается на средах с хлоридазоном, антипирином и L-фенилаланином. Слабо растут на средах с L-глутаматом, пируватом, фумаратом, сукцина- том и малатом и на разбавленных сложных средах. Большинство сахаров, спиртов, аминокислот, карбоновых кислот, а также обычных питательных сред сложного состава не используют. Оптимальная температура для роста 29-30°С. При 4 и 37°С не растут. При 37°С культуры погибают в течение нескольких дней. Оптимальный рН 6,8-7,0. Выделены из почвы путем накопления в минеральных средах, содержащих хлоридазон, антипирин или пирамидон. Деннтрифнкацию не осуществляют; нитрит из нитрата не образуют. В качестве единственных источников азота используют NH4+ и NO3–. Не способны расти за счет азотфиксации. Каталазоположительные, слабо оксидазоположительные. Желатину, казеин, крахмал и эскулин не гидролизуют. Уреазоотрицательные. Реакция с лакмусовым молочком отрицательная. Характерно слабое образование H2S из тиосульфата или цистеина. Проба с метиловым красным и реакция Фогеса-Проскауэра отрицательные. Индол отрицательные. Из сахаров и спиртов кислоту и газ не образуют. Не патогенны для крыс и кроликов.
Типовой (и единственный) вид: Phenylobacterium immobile.
Признаки вида: Как для рода.
Род PhylIobacterium
Прямые палочки (in vitro), 0,4–0,8 х 0,8–2,0 мкм. Грамотрицательные. Подвижные за счет едннственного полярного жгутика или нескольких полярных либо латеральных жгутиков с большой длиной волны. Аэробы; метаболизм чисто дыхательного типа с использованием кислорода в качестве конечного ацептора электронов. Оптимальная температура для роста 28–34°С. Колонии на агаризованной среде с глюкозой и дрожжевым экстрактом прозрачные, бесцветные или бежевого цвета, слизистые. Оксидазоположительные. Хемоорганотрофы; используют в качестве источников углерода разнообразные сахара или соли органических кислот. Не гидролизуют крахмал, пектин и целлюлозу. Встречаются в узелках на листьях высших растении (виды сем. Myrsinaceae [мирзнновые] и Rubiaceae [мареновые]).
Типовой вид; Phyllobacterium myrsinacearum.
Дифференциация видов рода Phyllobacterium: См. табл. 4.38.
Таблица 4.38. Дифференцирующие признаки видов рода Phyllobacterium 1 
Род Pseudomonas
Прямые или слегка изогнутые, но не спиральные, палочки, 0,5-1,0 x 1,5-5,0 мкм. У многих видов накапливается в качестве запасного источника углерода поли-β-гидроксибутират, который виден как включения после окраски суданом. Простек и чехлов не образуют. Покоящиеся стадии неизвестны. Грамотрицательные. Подвижные за счет одного или нескольких полярных жгутиков; в отдельных случаях неподвижные. У некоторых видов возможно также образование латеральных жгутиков с более короткой длиной волны. Аэробы; метаболизм чисто дыхательного типа с использованием кислорода в качестве конечного акцептора электронов; в некоторых случаях альтернативным акцептором электронов может служить нитрат, что обеспечивает анаэробный рост. Ксантомонадины не образуют. Большинство, если не все, виды не растут в кислой среде (рН 4,5). Большинство видов в органических факторах роста не нуждается. Оксидазоположительные или отрицательные. Каталазоположительные; хемоорганотрофы; некоторые виды – факультативные автотрофы, способные использовать в качестве источника энергии Н2 или СО. Широко распространены в природе. Некоторые виды патогенны для человека, животных или растений.
Типовой вид: Pseudomonas aeruginosa.
Дифференциация видов рода Pseudomonas: См. табл. 4.39-4.42.
Таблица 4.39. Дифференциация некоторых хорошо изученных
видов рода Pseudomonas 1,2 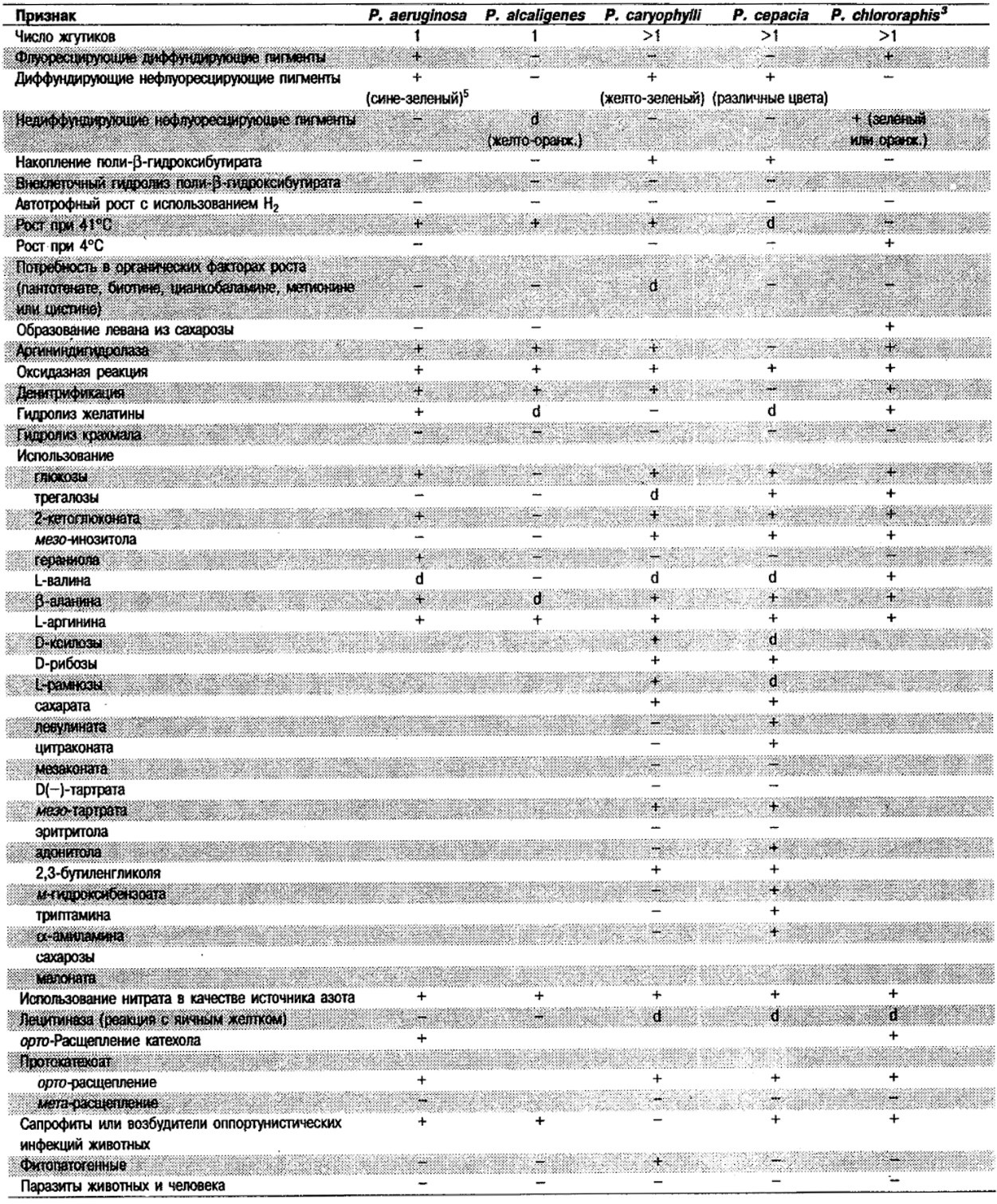
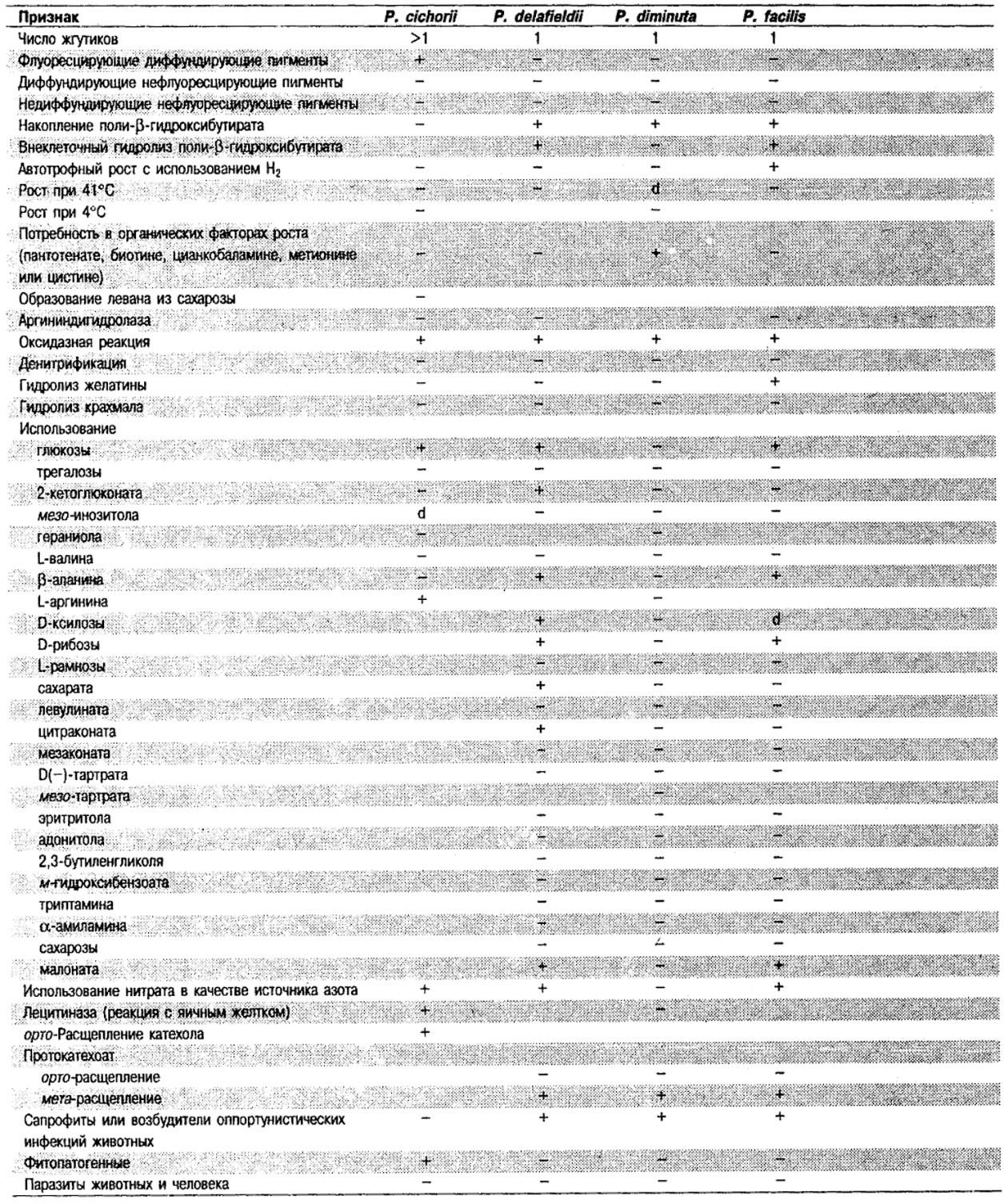

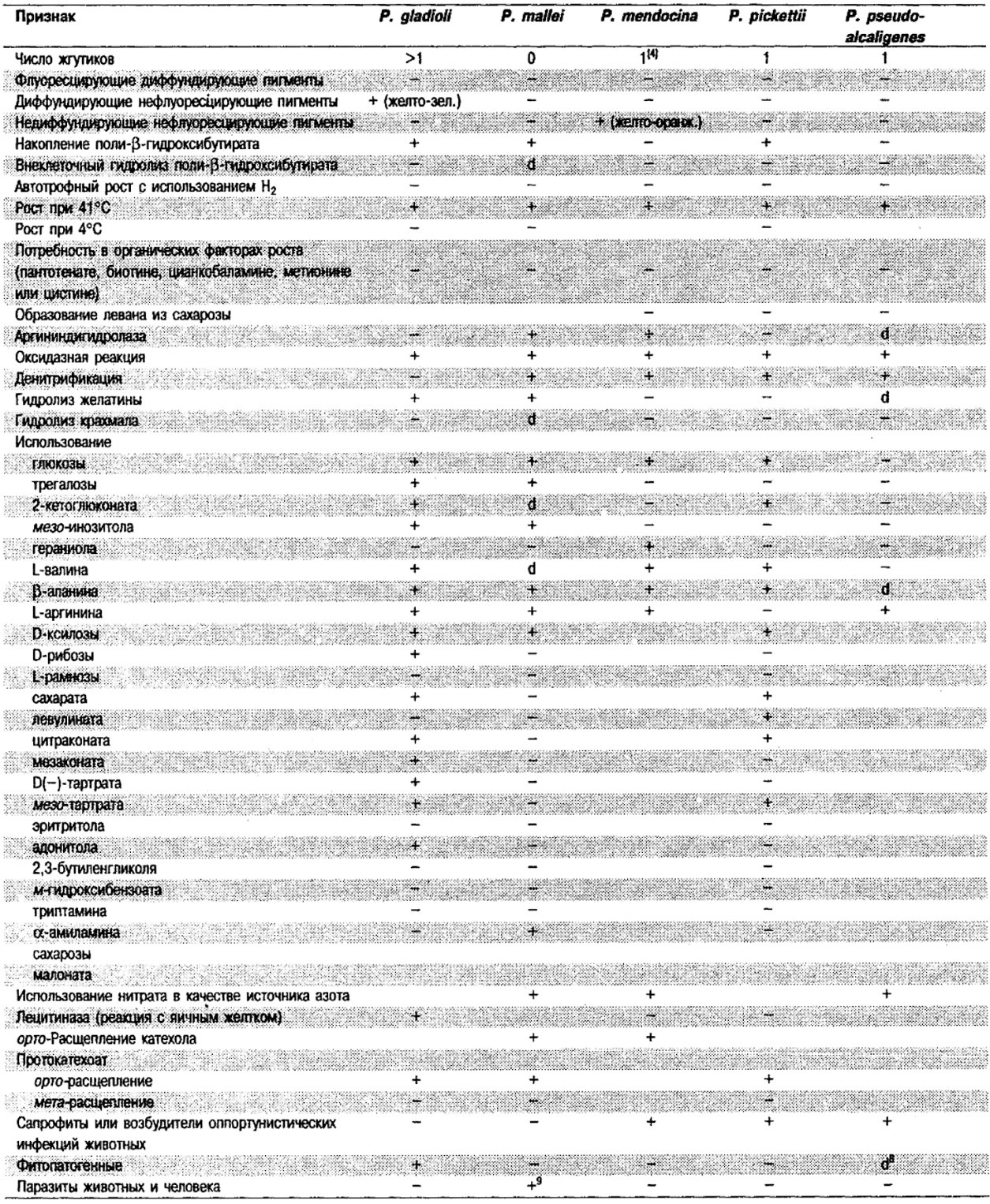

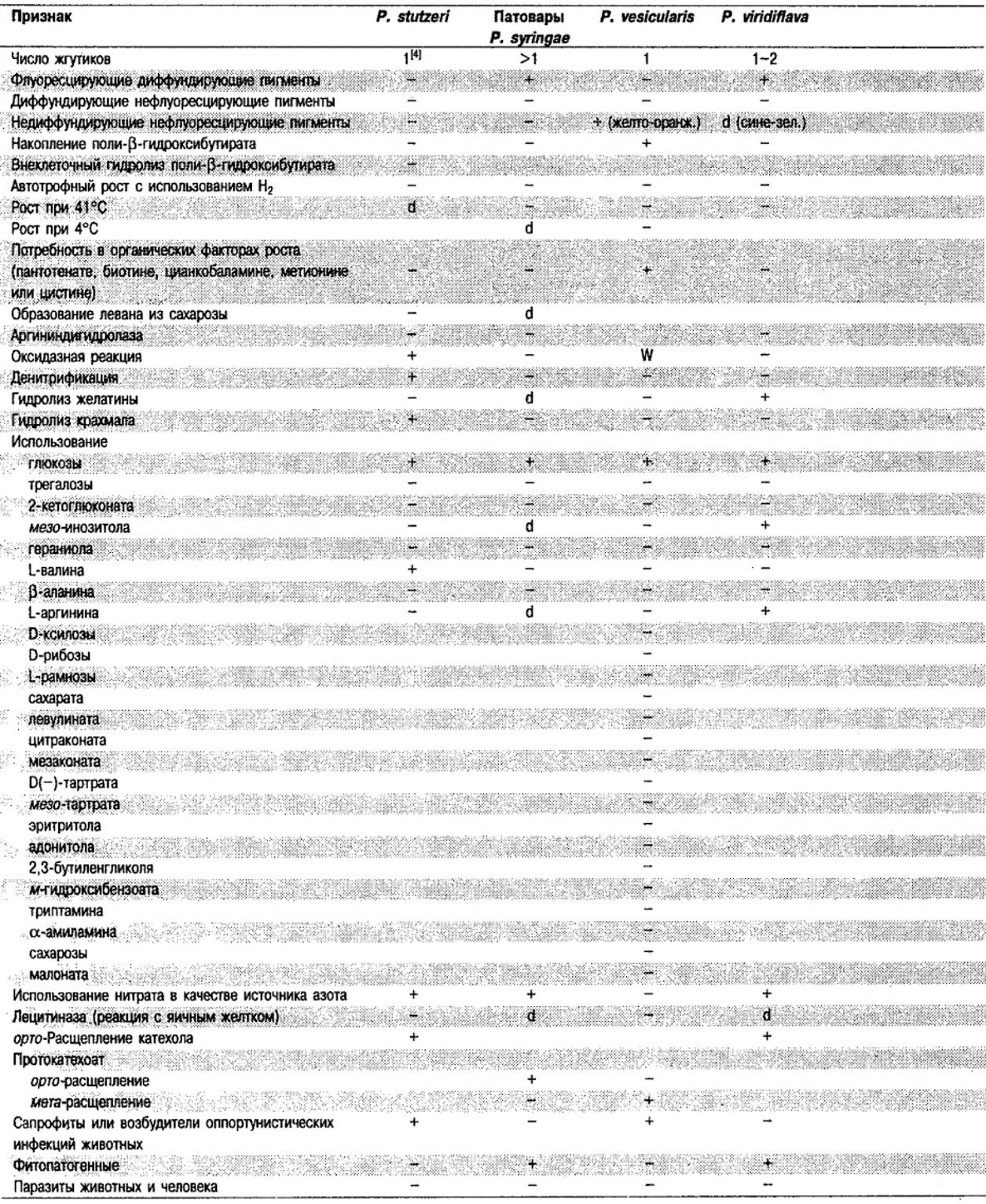

Таблица 4.40. Источники выделения
и признаки дополнительных видов Pseudomonas 1 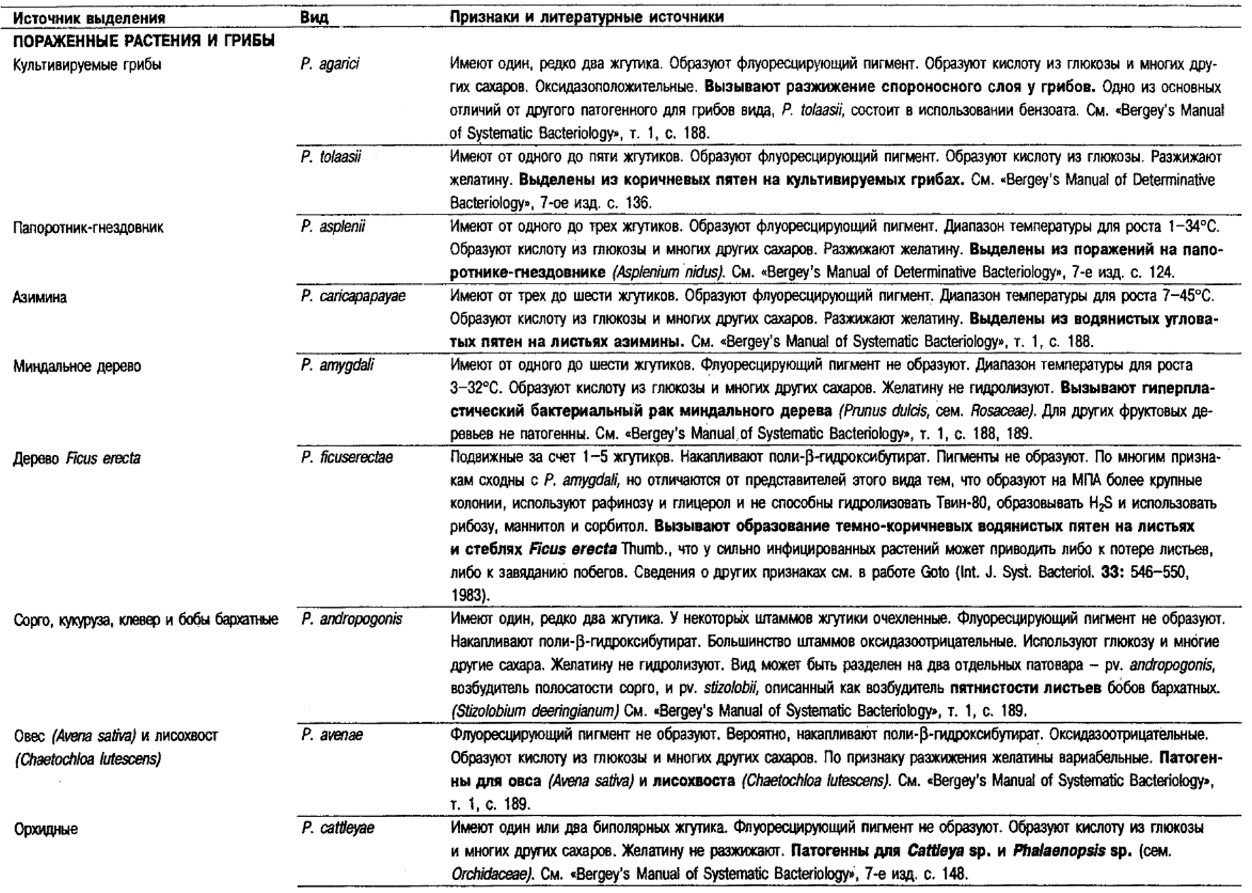

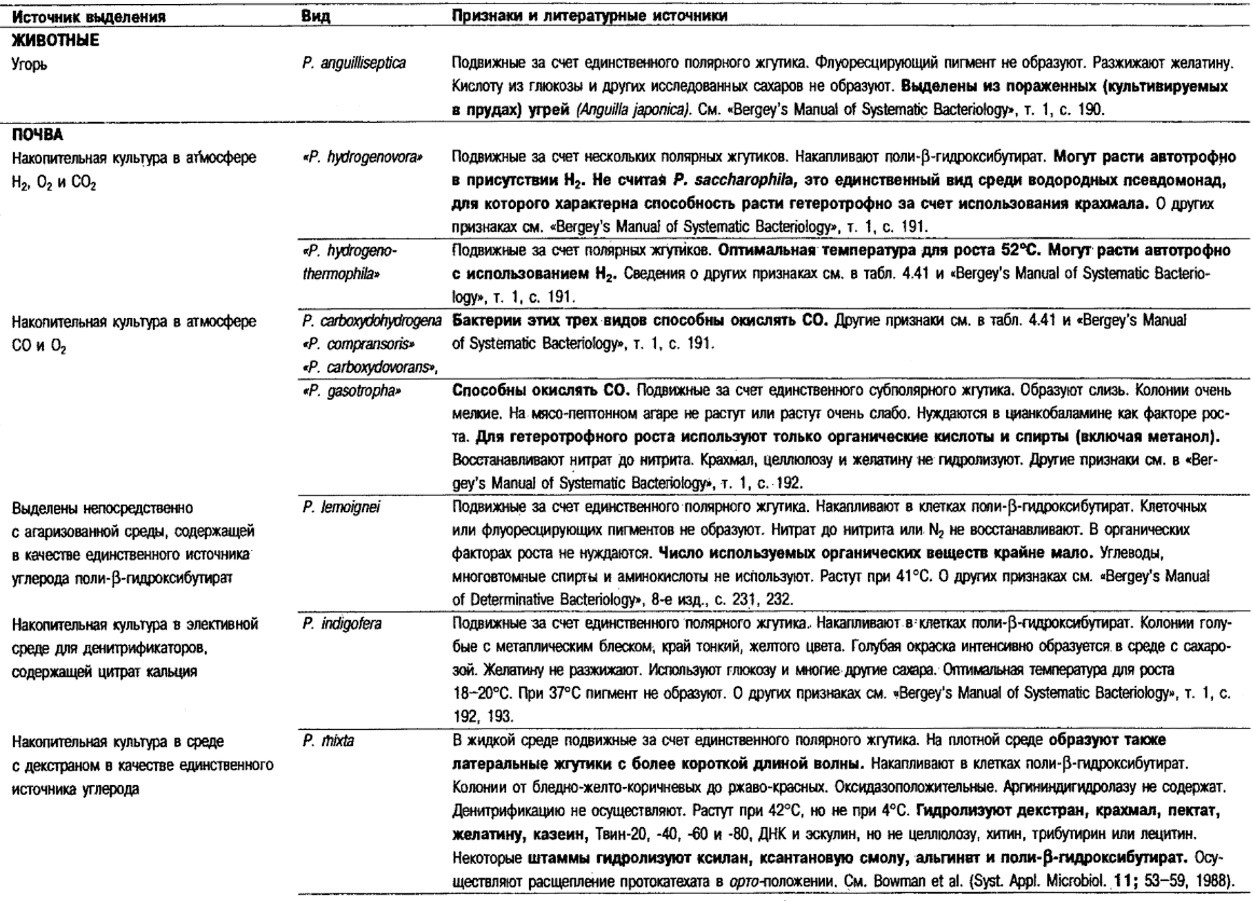



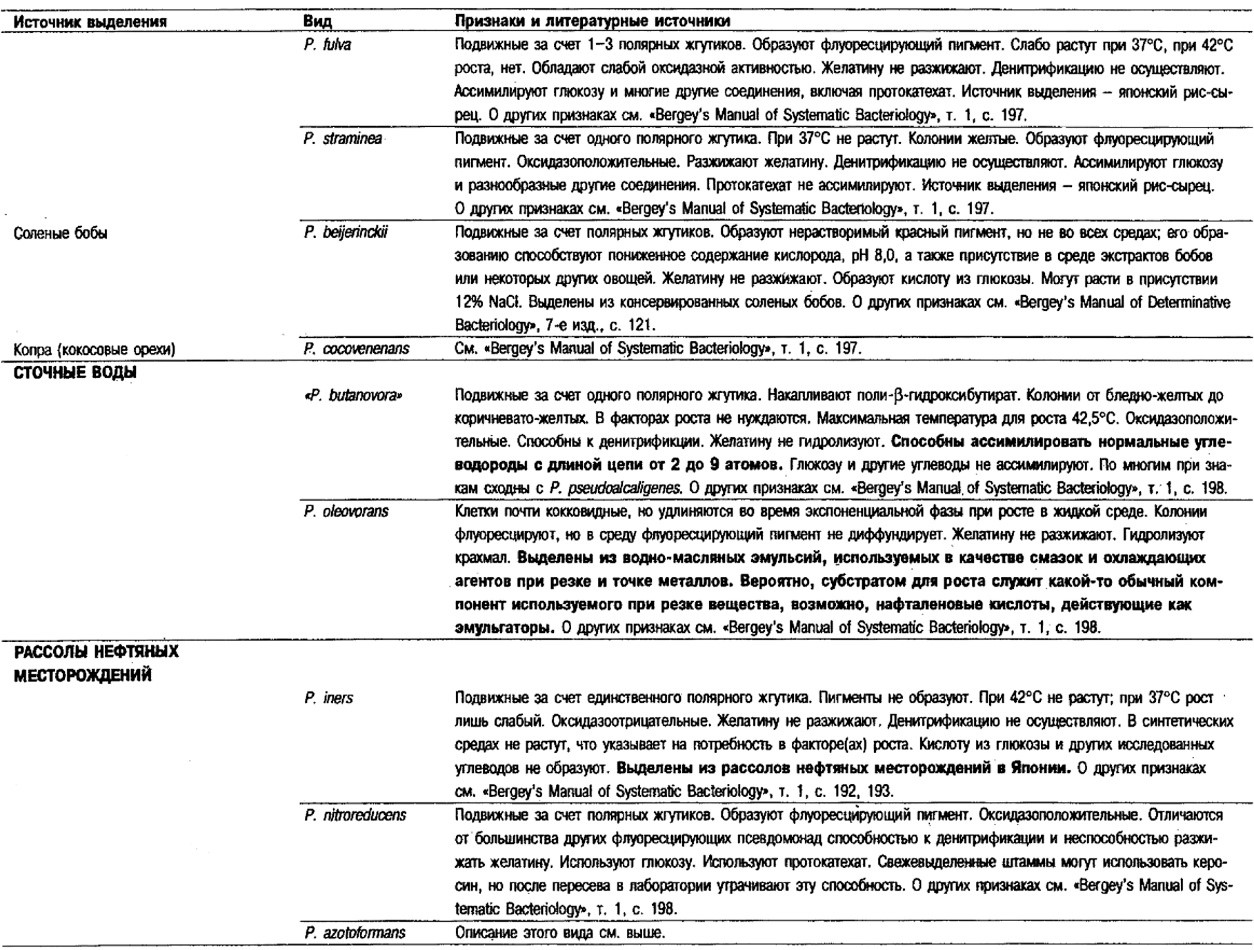

Таблица 4.41. Дифференцирующие признаки различных псевдомонад,
использующих водород и СО1 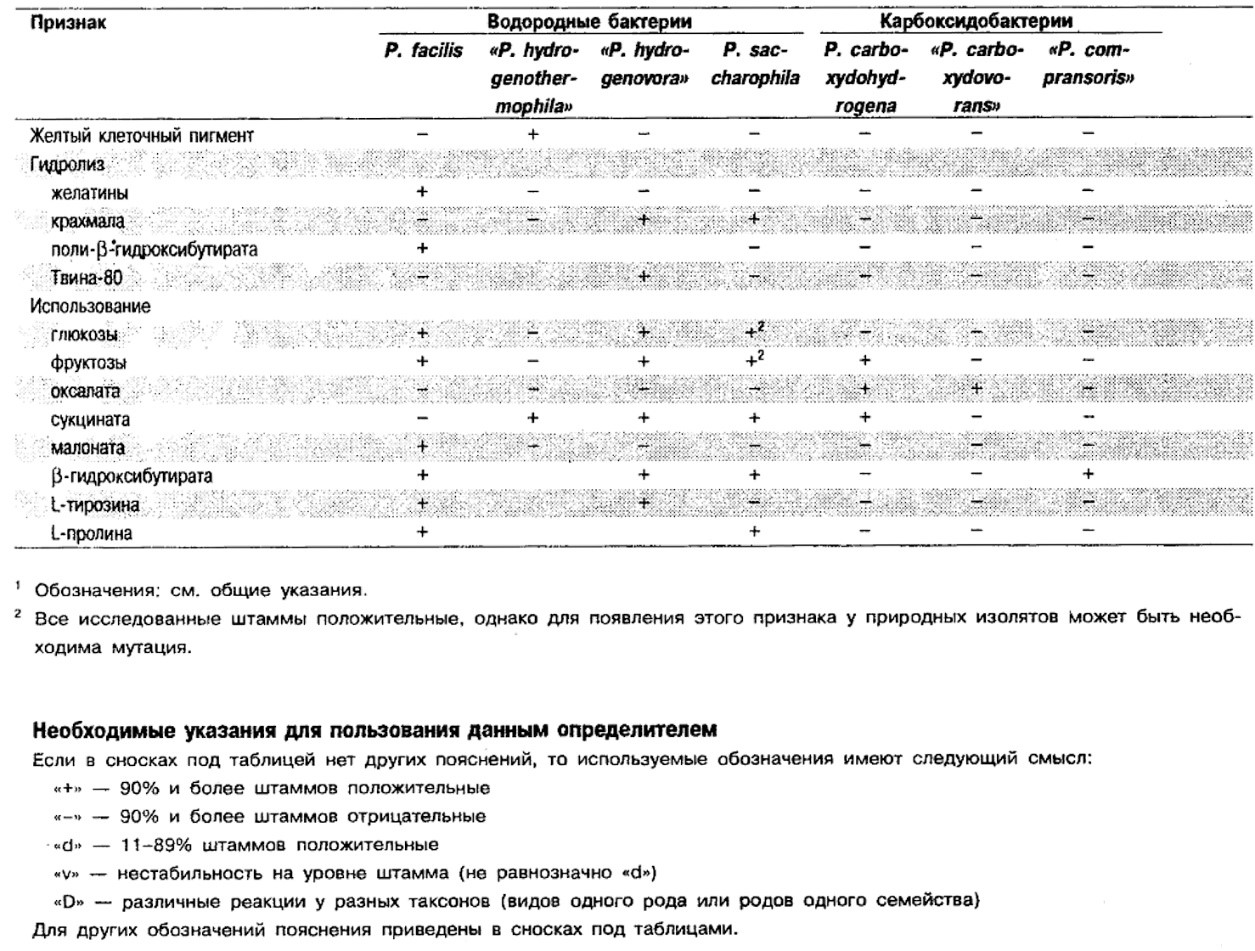
Таблица 4.42. Признаки, полезные для дифференциации
различных денитрифицирующих псевдомонад1 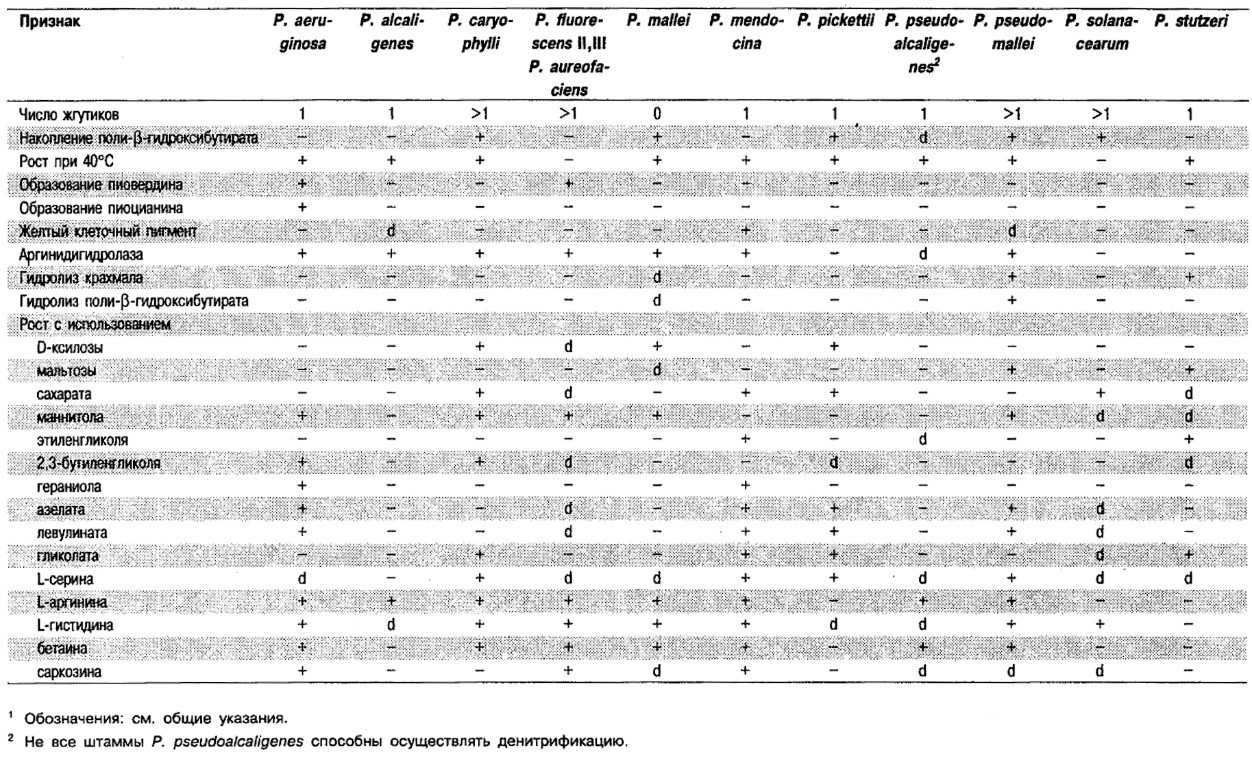
Род Psychrobacter
От редакции: Род Psychrobacter не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1986 г. (Juni, Heym, Int. J. Syst. Bacteriol. 36: 388-391) и включает единственный вид P. immobilis.
Коккобациллы, 0,9–1,3 x 1,5–3,8 мкм. Кокки часто имеют более или менее выраженную овальную форму, тогда как палочки могут варьировать по длине от очень коротких до относительнодлинных. У некоторых штаммов палочки могут быть несколько раздуты. Грамотрицательные. Неподвижные; аэробы; не пигментированные. Неспорообразующие. Каталазо- и оксидазоположительные. Большинство штаммов психрофилы, способные расти при 5°С (оптимальная температура 20°С); при 35–37°С, как правило, расти не способны. Штаммы, способные расти прн 35–37°С, обычно не могут расти при 5°С. Колонии на агаре с сердечной вытяжкой обычно гладкие и непрозрачные. Встречаются в рыбе, мясных и куриных продуктах. Многие штаммы образуют в аэробных условиях кислоту из глюкозы и ряда других сахаров. Штаммы, способные расти при 35-37°С, выделены из клинического материала от человека и животных.
Типовой (и единственный) вид: Psychrobacter immobilis.
Признаки вида: Большинство штаммов образует в аэробных условиях кислоту из глюкозы и ряда других сахаров, дезаминирует фенилаланин и триптофан, содержит уреазу, восстанавливает нитрат и гидролизует Твин-80. Не образуют кислоту из фруктозы и сахарозы; не гидролизуют крахмал.
Род Rhizobacter
От редакции. Род Rhizobacter не приведен в «Sergey's Manual of Systematic Bacteriology». Этот род описан в 1988 г. (Goto, Kuwata, Int. J. Syst. Bacteriol. 38: 233-239) и включает единственный вид R. daucus.
Грамотрицательные палочки, от прямых до слегка изогнутых. Подвижные за счет полярных или/и латеральных жгутиков. Образуют гранулы поли-β-гидроксибутирата. Хемоорганотрофы. Аэробы; метабол и зируют глюкозу по пути окисления, Используют D-глюкозу в качестве единственного источника углерода и энергии. На агаризованной среде образуют колонии, белые или желтовато-белые, складчатые, плотные или вязкие. Желтый пигмент отличен от ксантомонадинов. В жидких средах растут в виде хлопьев, состоящих из единиц-шариков; пальцеобразные выросты не отмечены. По следующим признакам положительные: окендаза, каталаза, образование H2S из цистеина, восстановление нитрата, о-нитрофенил-β-В-галактопиранозид, ингибирование KCN. Отрицательные по следующим признакам: денитрификация, проба с метиловым красным, аргининдигидролаза, нитрогеназа, флуоресцирующий пигмент, индол и ацетоин. Чувствительны к 10 мкг фосфата вибриостатического агента 0/129. Гидролизуют крахмал, декстрин и гликоген. Производные бензола в качестве источников углерода не используют. В NaCl и факторах роста не нуждаются. Содержат убихинон Q-8. Типовой вид встречается в почве и патогенен для растений: в естественных условиях вызывает образование галлов на корнях моркови.
Типовой вид: Rhizobacter daucus.
Признаки вида: Грамотрицательные капсулированные палочки, от прямых до слегка изогнутых, 0,9-1,3 х 2,1-2,5 мкм. Обычно небольшое число клеток в популяции обладает подвижностью за счет полярных или/и латеральных жгутиков. Псевдоолиготрофы. Колонии на картофельном агаре с пептоном и глюкозой (PPG-arap) плотные, белые, складчатые, а на агаре с дрожжевым экстрактом, пептоном и глюкозой (DYPG-arap) желтовато-белые. В жидких средах дают обильный рост в виде хлопьев, состоящих из шарообразных клеточных агрегатов, Положительные по уреазе, липазе (гидролиз Твина-80) и лецитиназе. Отрицательные по глкжонату, гидролизу казеина, нитрогеназе, ацетоину и 3-кетолактозе. Максимальная устойчивость к NaCl 0,7%. В качестве источника углерода используют D-арабинозу, рибозу, ксилозу, лактозу, целлобиозу, мелибиозу, сахарозу, рафинозу, крахмал, декстрин, гликоген, инулин, адонитол, инозитол, маннитол, сорбитол, салицин, галактуронат, лактат, мукат, хиннат, тартрат и метанол. Не используют как источник углерода трегалозу, мелецитозу, дульцитол, эритритол, ацетат, цитрат, малонат и бензоат. Проба на гиперчувствительность к табаку отрицательная.
Род Rhizobium
Палочки, 0,5-0,9 x 1,2-3,0 мкм. В неблагоприятных для роста условиях обычно плеоморфные. Как правило, содержат гранулы поли-β-гидрокенбутирата, преломляющие свет при фазово-контрастной микроскопии. Грамотрицательные. Подвижные за счет единственного полярного moih субполярного жгутика либо перитрихиальных жгутиков в количестве от двух до шести. У небольшого числа штаммов обнаружены фимбрии. Аэробы; метаболизм дыхательного типа с использованием кислорода в качестве конечного акцептора электронов. Во многих случаях способны хорошо расти при парциальном давлении кислорода ниже 1,0 кПа. Оптимальный диапазон температуры 25–30°С, рН 6-7. Колонии округлые, выпуклые, полупрозрачные, приподнятые, слизистые, обычно диаметром 2–4 мм через 3-5 сут инкубации на агаре с дрожжевым экстрактом» маниитолом и минеральными солями. При росте в жидкой среде с перемешиванием культуры достигают значительной плотности через 2 или 3 сут инкубации. Хемоорганотрофы; используют в качестве источников углерода разнообразные углеводы и соли органических кислот; газ не образуют. Целлюлозу и крахмал не используют. Среду с минеральными солями и манннтолом или другими углеводами при росте подкисляют. Рост на среде с углеводами обычно сопровождается обильным образованием внеклеточной слизи полисахаридной природы. Источниками азота могут служить соли аммония, нитрат, нитрит и большинство аминокислот. Некоторые штаммы могут расти в простой синтетической среде с не содержащим витаминов гидролизатом казеина в качестве источника и углерода и азота. Пептон используют слабо. Казеин и агар не гидролизуют. Некоторые штаммы нуждаются в биотине или других водорастворимых витаминах. 3-Кетогликозиды не образуют. Характерная особенность этих организмов – способность проникать в корневые волоски бобовых растений (сем. Legumittosae) умеренного пояса, а также некоторых представителей бобовых тропического пояса и вызывать образование корневых клубеньков, внутри которых бактерии присутствуют как внутриклеточные симбионты. Все штаммы обнаруживают сродство к определенному кругу хозяев («специфичность» по хозяину). В корневых клубеньках бактерии присутствуют в виде плеоморфных форм – бактероидов, обычно осуществляющих фиксацию атмосферного азота с превращением его в связанную форму (аммиак), доступную для использования растением-хозяином.
Типовой вид: Rhizobium leguminosarum.
Дифференциация видов рода Rhizobium: См. табл. 4.43 и 4.44.
Таблица 4.43. Дифференцирующие признаки видов рода Rhizobium1 
Таблица 4.44. Признаки видов Rhizobium, связанные с образованием клубеньков1 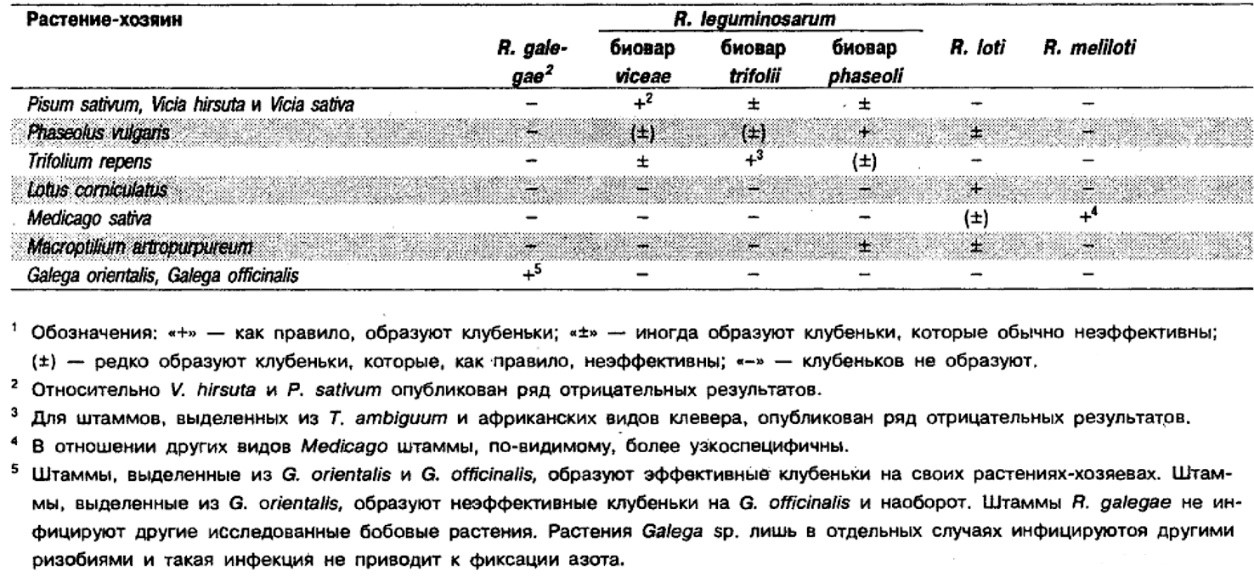
Род Rhizomonas
От редакции: Род Rhizomonas не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1990 г. (Van Bruggen et al., Int. J. Syst. Bacteriol. 40: 175-188) и включает единственный вид R. suberifaciens.
Прямые или слегка изогнутые палочки, 0,5-1,2 мкм. Грамотрицательные. Если подвижные, имеют один латеральный, субполярный или полярный жгутик. Покоящиеся стадии неизвестны. Размножаются путем бинарного деления. Накапливают внутриклеточные гранулы поли-β-гидроксибутирата; не содержат аргининдигидролазу. Колонии не флуоресцирующие, белые или желтоватые, гладкие или морщинистые. Облигатные аэробы; метаболизм окислительный. Оптимальный диапазон температуры примерно 28-33°С, максимальная температура – между 36 и 42°С, Этанол в уксусную кислоту не превращают, Оксидазо- и каталазоположительные. Денитрификацию с образованием N2 не осуществляют. Содержат убихинон Q-10. В составе клеточных жирных кислот преобладают четные ненасыщенные (C18:1 и С16:1) и насыщенные (С16:0) неразветвленные жирные кислоты, а также дигидроксилированные жирные кислоты С14:0 и C 17:1. Жирная кислота С18:1 составляет не менее 50% от общего количества жирных кислот. Вызывают опробковение корней у латука.
Типовой (и единственный) вид: Rhizomonas suberifaciens.
Признаки вида: Как для рода.
Вид Rochalimaea henselae
От редакции: Вид Rochalimaea henselae не приведен в «Bergey's Manual of Systematic Bacteriology». Этот вид описан в 1992 г. (Regnery et al., J. Clin. Microbiol. 30: 265-274; Int. J. Syst. Bacteriol. 42: 511, 1992).
Слегка изогнутые палочки, 0,5-0,6 x 1,0-2,0 мкм, Грамотрицательные. Легко окрашиваются красителем Хименеса. Жгутики не обнаружены. Оксидазо-, уреазо- и каталазоотрицательные. Лучше всего растут на богатых средах с эритроцитами, таких как среда с сердечно-мозговой вытяжкой (BHIA) и среда с триптиказой и продуктом обработки папаином соевых бобов (TSA) плюс 5% крови барана. В тестах на использование углеводов активности не обнаруживают. Колонии на начальных этапах выделения твердые, с неровной поверхностью, по форме напоминающие цветную капусту, вросшие в агар. Предположительно вызывают бактериальный ангиоматоз и, возможно, болезнь от кошачьих царапин у человека.
Род Roseobacter
От редакции. Род Roseobacter не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1991 г. (Shiba, Syst. Appl. Microbiol. 14: 140-145; Int. J. Syst. Bacteriol. 41: 331, 1991) и вхлючает два вида: R litoralis и R. denitrificans.
Клетки овальные или палочковидные, 0,6-0,9 х 1,0-2,0 мкм. Подвижные за счет субполярных жгутиков. Грамотрицательные. Размножаются бинарным делением. Растут гетеротрофно в аэробных условиях. Обнаруживают фотосинтетическую активность в аэробных условиях. Содержат бактериохлорофилл α, этерифицированный фитрлом. В анаэробных условиях бактериохлорофилл не синтезируют. Спектр поглощения клеток включает полосу интенсивного поглощения при 805-807 нм и меньшую по интенсивности полосу при 868-873 нм в ближней инфракрасной области. Основной каротиноид – сфероиденон. Основной хинон – убихинон-10. Менахиноны отсутствуют. В составе клеточных жирных кислот преобладает жирная кислота С18:1. Для роста необходимы ионы Na+, биотин, тиамин и никотиновая кислота. Оптимальный диапазон рН 7,0-8,0, температуры – 20-30°С, Единственными источниками органического углерода могут служить некоторые органические кислоты. Метанол не используют. Чувствительны к хлорамфениколу, пенициллину, тетрациклину, стрептомицину и полимиксину В. Гидролизуют желатину и Твин-80. Содержат каталазу и оксидазу. Содержаниие G+C в ДНК 56-60 мол.%.
Типовой вид: Roseobacter litoralis.
Дифференциация видов рода Roseobacter и Erythrobacter longus: См. табл. 4.45.
Таблица 4.45. Дифференцирующие признаки видов Roseobacter и Erythrobacter longus1 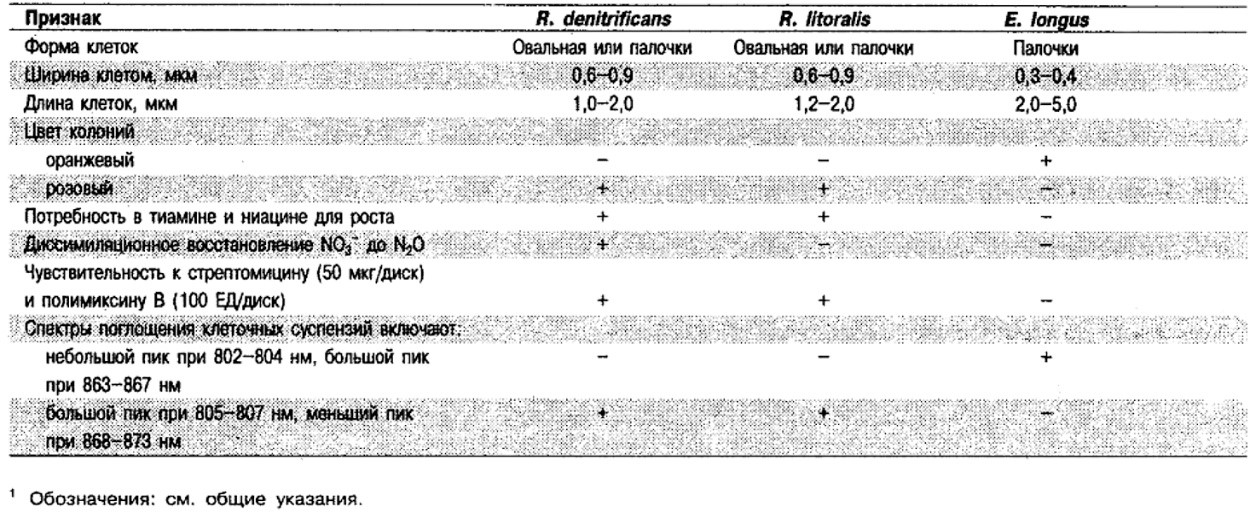
Род Rugamonas
От редакции: Род Rugamonas не приведен в «Sergey's Manual of Systematic Bacteriology». Этот род описан в 1986 г. (Austin, Moss, J. Gen. Microbiol. 132: 1899-1909; Int. J. Syst. Bacteriol. 37: 179-180, 1987) и включает единственный вид R. rubra.
Грамотрицательные палочки с закругленными концами. Подвижные за счет одного или большего числа субполярных или полярных жгутиков. Колонии на агаризованной среде (5 сут при 20°С) морщинистые, приподнятые, резиновой консистенции. В жидкой среде через 7 сут инкубации образуют хлопья, через 5 сут клетки становятся плеоморфными. Хемоорганотрофы; метаболизм дыхательного, никогда не бродильного, типа. Каталазо- и оксидазоположительные.
Типовой (и единственный) вид; Rugamonas rubra.
Признаки вида: Размер клеток 0,8-0,9 х 2,4-4,0 мкм. Примерно через 7 сут роста образуют внутриклеточные гранулы продигиозина. Молодые клетки подвижны за счет одного или более полярных или субполярных жгутиков. В жидкой среде Беннета через 7 сут инкубации образуют розовые/красные хлопья. Колонии на агаризованной среде Беннетта (20°С) через 2-3 сут белые, блестящие, округлые, через 4 сут становятся розовыми, а через 5 сут темно-красными, морщинистыми, резиновой консистенции, диаметром примерно 3 мм. Восстанавливают нитраты до N2. Температура для роста 4-30°С, но не 37°С. Оптимальная температура примерно 25°С. Растут в присутствии NaCl (0,5%) при рН 5-9. Аргинин-декарбоксилазу не образуют; некоторые штаммы синтезируют лизин- и орнитиндекарбоксилазу. Разлагают эскулин, хитин, ДНК, РНК, желатину, лецитин, Твин-20, -40, -60 и -80, тирозин и мочевину, но не аллантоин, целлюлозу, эластин, гипоксантин, крахмал и ксантин. Содержат β-галахтозидазу и фосфатазу. H2S и фенилаланиндезаминазу не образуют. Результаты тестов с глюконатом и на среде Козера с цитратом положительные; реакция Фогеса-Проскауэра, проба с метиловым красным и малонатом отрицательные. Образуют следующие ферменты: щелочную фосфатазу, кислую фосфатазу, эстеразу (некоторые штаммы), эстеразу-липазу, лейцинариламидазу и фосфоамидазу. В качестве единственного источника углерода используют D-аланин, L-apaбинозу, DL-аргинин, м-инозитол, DL-лактат натрия, целлобиозу, D-фруктозу, D-галактозу, L-гистидин, мальтозу, D-маннозу, D-рафинозу, D-рибозу, DL-серин, ацетат натрия, трехзамещенный цитрат натрия, D-сорбитол, сахарозу и трегалоэу. Не используют адонитол, i-эритритол, малонат натрия, D-мелибиозу. D-мелецитозу, D-рамнозу, DL-валин и D-ксилозу. Некоторые штаммы используют лактозу. Выделены из речной воды.
Род Serpens
Палочки, 0,3-0,4 x 8-12 мкм, одиночные или в парах. Цист или кокковидных тел не образуют. В стационарной фазе роста клетки увеличиваются в длину до 16-25 мкм и часто имеют пузырьки или сферические выросты. Грамотрицательные. Чрезвычайно гибкие и способны к змеевидному движению в агаровых гелях. Имеют биполярные пучки из 4-10 жгутиков. Поли-β-гидроксибутират или другие внутриклеточные включения не образуют. Метаболизм чисто дыхательного типа с использованием кислорода в качестве конечного акцептора электронов. Растут в аэробных условиях, но предпочтительно при более низких, чем атмосферная, концентрациях кислорода. Каталазо- и оксидазоположительные. Хемоорганотрофы. Единственным эффективным источником углерода и энергии служит лактат, хотя очень слабый рост возможен за счет использования ацетата или α-кетоглутарата. Углеводы, жирные кислоты и сахароспирты не катаболнзируют. Источниками азота могут служить гидролизат казеина, пептон, дрожжевой экстракт и для большинства штаммов хлористый аммоний; нитрат и нитрит не используют. Витамины стимулируют рост, но их присутствие не обязательно. Оптимальная температура 28-30°С. На средах с 1,8-2,0% агара образуют колонии кремового цвета, округлые, диаметром 3-6 мм, с нитевидным краем. На средах, содержащих менее 1,5% агара, образуют только подповерхностные распространяющиеся колонии. Обнаружены в осадках эвтрофных пресноводных водоемов.
Типовой вид: Serpens ftexibilis.
Признаки вида: Как для рода.
Род Sinorhizobium
От редакции: Род Sinorhizobium не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1988 г. (Chen et al., Int. Syst. Bacteriol. 38: 392-397) и включает два вида: S. fredii и S. xinjiangensis.
Палочки 0,5-0,9 x 1,2-3,0 мкм. Обычно содержат гранулы поли-β-гидроксибутирата. Спор не образуют. Грамотрицательные. Подвижные за счет единственного полярного жгутика или перитрихнальных жгутиков в количестве от 1 до 3 и более. Аэробы; метаболизм дыхательного типа с использованием кислорода в качестве конечного акцептора электронов. Колонии округлые, выпуклые, полупрозрачные, приподнятые, слизистые, обычно диаметром 2-4 мм через 3-5 сут инкубации на агаризованной среде с дрожжевым экстрактом и маннитолом (YMA). В жидкой среде заметное помутнение наблюдается через 3-5 сут культивирования на качалке. Оптимальная температура 25-30°С, но большинство штаммов растут и при 35°С, а некоторые при 10°С. Оптимальный диапазон рН 6-8. Некоторые штаммы растут при рН 5,0, другие при рН 10,5. Большинство штаммов растет в присутствии 1%, но не 1,5% NaCl, хотя отдельные штаммы хорошо растут на среде YMA, содержащей 4,5% NaCl. В качестве единственного источника углерода используют следующие субстраты: D-арабинозу, целлобиозу, фруктозу, D-галактозу, глюкозу, L-глутамин, лактозу, D-маннозу, маннитол, D-рибозу, сукцинат натрия, ксилозу и D-туранозу. Не используют следующие субстраты: тартрат аммония, оксалат аммония, целлюлозу, дульцитол, фукозу, глицин, сорбозу, альгинат натрия и ванилиновую кислоту. Все штаммы образуют кислоту при росте на среде YMA. Предпочтительные источники азота – хлористый аммоний и нитраты, а не аминокислоты, но некоторые штаммы используют отдельные аминокислоты. Пептон используют слабо. Все штаммы нуждаются в пантотенате и никотиновой кислоте, потребности в других витаминах варьируют. Оксидазо- и каталазоположительные. Симбиотические азотфиксаторы. Не имеют широкого спектра хозяев, но эффективно вызывают образование клубеньков у Glycine soja, Glycine max с v. TGmll9, G. max cv. TGml20, G. max cv, TGm344, G. max cv. Kai-Yu No. 8, Vigna unquiculata cv. VITA-3 и Cajanus cajan cv. CITA-l Неэффективные симбиозы образуют с Vigna radiata, Macropti-Hum atropurpureum, Macroptilium lathyroides и Sesba-nia cannabina.
Типовой вид: Sinorhizobium fredii.
Дифференциация видов рода Sinorhizobium: См. табл. 4.46.
Таблица 4.46. Дифференцирующие признаки видов рода Sinorhizobium 1,2 
Род Sphingobacterium
От редакции: Род Sphingobacterium не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1983 г. (Yabuuchi et al., Int. Syst. Bacteriol. 33: 580-598) и включает три вида: S. spiritivorum, (прежнее название – Flavobacterium spiritivorum), S. multivorum (прежнее название – Flavobacterium multivorum) и S. mizutae.
Прямые палочки. Спор не образуют. Грамотрицательные. Жгутиков не имеют, однако некоторые виды на полужидких средах обнаруживают скользящее движение. Каталазоположительные. Хемоорганотрофы; специфических потребностей в факторах роста не имеют. Колонии через несколько суток инкубации при комнатной температуре обычно становятся желтыми. Индол и ацетилметилкарбинол не образуют, Непротеолитические; желатину не гидролизуют. Образуют кислоту из углеводов в результате их окисления, но не брожения. В состав клеточных липидов входят сфинголипиды, церамидные составляющие которых представлены, главным образом, разветвленным дигидронасыщенным сфингозином C17:0, а основная кислота – i-2-OH-C15:0.
Типовой вид: Sphingobacterium spiritivorum.
Дифференциация видов рода Sphingobacterium: См. табл. 4.47.
Таблица 4,47. Дифференцирующие признаки видов рода Sphingobacterium 1,2 
Род Thermoleophilum
От редакции: Род Thetmoleophilum не приведен в «Beigey's Manual of Systematic Bacteriology». Этот род описан в 1984 г. (Zarilla, Perry, Arch. Microbiol. 137: 286-290; Int. J-Syst. Bacteriol. 36: 354-356, 1986) и исходно включал единственный вид Т. album. В 1986 г. добавлен второй вид Т, minutum (Zarilla, Perry, Int. J. Syst. Bacteriol. 36: 13-16).
Короткие палочки, 0,4 x 0,7-1,5 мкм. Спор не образуют. Аэробы. Образуют очень мелкие просвечивающие колонии на поверхности агара. Растут только в присутствии избытка н-алканов из 13-20 атомов углерода в синтетической среде с минеральными солями. В факторах роста не нуждаются. Оптимальная температура для роста 60°С, диапазон температуры 45-70°С. Оптимальное значение рН близко к нейтральному, диапазон рН 5,8-8,0. Могут быть выделены из образцов ила и воды, главным образом из термальных сред, однако обнаружены также и в нетермальных местообитаниях.
Типовой вид: Thermoleophilum album.
Дифференциация видов рода Thermoleophilum: В настоящее время не существует рутинных фенотипических тестов, позволяющих отличить Т. album от Т. minutum. Эти два вида можно различить на основании данных по гибридизации ДНК, а также по разнице в электрофоретической подвижности белков, высвобождаемых из клеток при обработке ультразвуком.
Род Thermomicrobium
Короткие палочки неправильной формы, 1,3-1,8 х 3,0-6,0 мкм. Плеоморфные клетки могут иметь колокодообразную форму или варьирующий диаметр, располагаться одиночно или парами. Ни покоящихся стадий (например, «округлых тел»), ни эндоспор не образуют. Грамотрицательные. Клеточная стенка не содержит в значительных количествах диаминокислоту в составе пептндогликана. Неподвижные. Облигатные аэробы. Оптимальная температура для роста 70-75°С, максимальная – 80°С, минимальная – 45°С, Оптимальный диапазон рН 8,2-8,5, однако хорошо растут в диапазоне рН 7,5-8,7. Хемоорганотрофы; метаболизм чисто дыхательного типа с использованием кислорода в качестве конечного акцептора электронов. Каталазоположительные. Колонии розовые. Максимальный рост обнаруживается на среде с дрожжевым экстрактом и пептоном (концентрация по 0,5%). Глюкозу в качестве единственного источника углерода и энергии не используют, н-Алканы не используют. Время генерации 5,5 ч. Выделены из горячего источника в Йеллоустонском национальном парке, США.
Типовой вид; Thermomicrobium roseum.
Признаки вида: Как для рода.
Род Thermus
Прямые палочки, 0,5–0,8 x 5,0–10,0 мкм. В некоторых условиях культивирования могут образовывать нити длиной от 20 до 200 нм и более. Большинство штаммов образуют округлые тела – крупные сферические структуры диаметром 10–20 мкм, возникающие в результате соединения отдельных клеток; эти тела обычно присутствуют в старых культурах. Неподвижные; жгутиков не имеют. Эндоспор не образуют. Грамотрицательные. Колонии у большинства штаммов желтые, оранжевые или красноватые за счет каротиноидных пигментов. Аэробы; метаболизм чисто дыхательного типа с использованием кислорода в качестве конечного акцептора электронов. Оксидазо- и каталазоположительные. Желатину обычно гидролизуют. Крахмал обычно разлагают слабо. Как правило, восстанавливают нитрат до нитрита. Термофильные, оптимальная температура для роста 70–75°С. Оптимальное значение рН близко к нейтральному. Обнаружены в горячих источниках с рН от щелочного до нейтрального, а также в водонагревательных установках и в природных водоемах, подвергающихся термальному загрязнению.
Типовой вид; Thermus aquatic us.
Дифференциация видов рода Thermus: См. табл. 4.48.
Таблица 4.48. Дифференцирующие признаки видов рода Thermus 1,2 
Род Variovorax
От редакции: Род Variovorax не приведен в «Bergey's Manual of Systematic Bacteriology», Этот род описан в 1991 г. (Willems et al., Int. J. Syst, Bacteriol. 41: 445–450) и включает единственный вид V paradoxus (прежнее название – Alcaligenes paradoxus).
Палочки, от прямых до слегка изогнутых, 0,5-0,6 x 1,2-3,0 мкм, одиночные или в парах. Подвижные за счет дегенеративных перитрихиальных жгутиков. Грамотрицательные. Колонии желтые за счет каротиноидных пигментов. Оксидазо- и каталазоположительные. Аэробы и хемоорганотрофы. Некоторые штаммы способны к литоавтотрофному росту, при котором источником энергии служит водород. Окисление углеводов осуществляют с использованием кислорода в качестве конечного акцептора электронов. Хорошо растут на средах с углеводами, органическими кислотами (включая аминокислоты) или пептоном.
Типовой (и единственный) вид: Variovorax paradoxus.
Признаки вида: Как для рода: См. также табл. 4.14.
Род Volcaniella
От редакции: Род Volcaniella не приведен в «Вегgey's Manual of Systematic Bacteriology». Этот род описан в 1990 г, (Quesada et al., Int. J. Syst. Bacteriol. 40: 261–267) и включает единственный вид Кeurihalina.
Клетки в форме коротких прямых палочек, 0,8-1,0 x 2,0-2,5 мкм, окружены капсулой, одиночные или в парах. Неподвижные. Грамотрицательные. Накапливают поли-β-гидроксибутират в качестве запасного вещества. Эндоспор и чехлов не образуют. Метаболизм дыхательного типа с использованием кислорода в качестве конечного акцептора электронов. Умеренные галофbлы; способны расти при концентрациях соли 5-20% (масса/объем). Оптимальная концентрация соли 7,5% (масса/объем). Хемоорганотрофы. Все штаммы хорошо растут при рН 5-10 и температуре 15-45°С. Оксидаэоотрицательные и каталазоположительные. Выделены из сильно засоленных мест обитания (почва, соленые водоемы) и морской воды.
Типовой (и единственный) вид: V. eurihalina.
Признаки вида: Как для рода.
Род Weeksella
От редакции: Род Weeksella не приведен в «Вегgey's Manual of Systematic Bacteriology». Этот род описан в 1986 г. (Holmes et al., Syst. Appl. Microbiol. 8: 185-190, Int. J. Syst. Bacteriol. 37: 179-180, 1987) и исходно включал единственный вид W. virosa (известный ранее как С DC Group IIF). В 1986 г. в него добавлен второй вид W. zoohelcum (ранее известный как CDC Group Ilj) (Holmes et al., Syst. Appl. Microbiol. 8: 191-196; Int. J. Syst. Bacteriol. 37: 179, 1987).
Палочки с параллельными сторонами н закругленными концами, обычно 0,6 x 2-3 мкм. Не содержат внутриклеточных гранул поли-β-гидроксибутирата. Эндоспор не образуют. Грамотрицательные. Неподвижные. Не скользят и не распространяются. Аэробы; метаболизм чисто дыхательного типа. Растут при температурах 18-42°С. При росте на плотной среде пигментов не образуют. Колонии округлые (диаметром 0,5-2 мм), слегка выпуклые, гладкие, блестящие, с цельным краем. Каталазо- и оксндазоположительные. Агар не разлагают. Хемоорганотрофы; сахара не разлагают. Иyдолположнтельные. В обычной природной среде не обнаружены; по-видимому, это внутренние паразиты, сапрофиты или комменсалы человека и пойкилотермных животных.
Типовой вид: Weeksella virosa.
Дифференциация видов рода Weeksella: См. табл. 4.49.
Таблица 4.49. Дифференцирующие признаки видов рода Weeksella 1,2 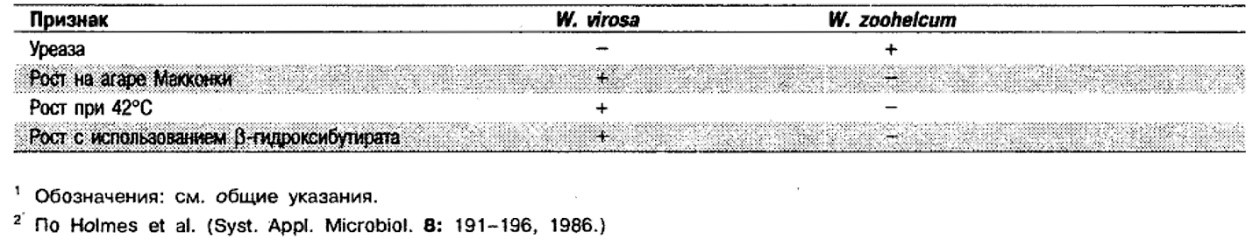
Род Xanthobacter
Палочки, 0,4-1,0 x 0,8-6,0 мкм. Некоторые виды образуют плеоморфные клетки при росте на средах с сукцннатом или кокковндные клетки на средах, содержащих в качестве единственного источника углерода спирт. Содержат равномерно распределенные в цитоплазме светопреломляющие (полифосфаты) и липидные (поли-β-гидроксибутират) гранулы. Покоящиеся стадии неизвестны. Грамположительные или грамвариабельные, однако клеточная стенка, по-видимому, грамотрицательного типа, т. е. содержит пептидогликан, в котором поперечные сшивки образованы мезо-диаминопимелиновой кислотой, и липополисахариды. Неподвижные или подвижные за счет перитрихнальных жгутиков. Аэробы; метаболизм чисто дыхательного типа с использованием кислорода в качестве конечного акцептора электронов. Оптимальная температура 25-30°С. Оптимальный диапазон рН 5,8-9,0. Колонии обычно непрозрачные и слизистые (хотя существуют и виды, не образующие слизи), окрашены в желтый цвет за счет водонерастворимого каротиноидного пигмента (дирамно-зидзеаксантин). Каталазоположительные. Все штаммы способны расти как хемолнтоавтотрофы в минеральной среде в атмосфере Н2, О2 и СО2 (7:2:1, объем/объем), а также как хемогетеротрофы, используя в качестве единственного источника углерода метанол, этанол, н-пропанол, н-бутанол или различные органические кислоты. Использование углеводов ограничено; из некоторых углеводов ни летучих или нелетучих кислот, ни газа не образуют. Ряд штаммов нуждается в витаминах. При росте в среде без азота фиксируют атмосферный азот, однако большинство штаммов осуществляют этот процесс только при пониженном давлении кислорода. Эффективность азотфиксации >10, обычно ~20 мг фиксированного азота на 1 г потребленной сахарозы. Образование клубеньков на корнях растений не индуцируют. Встречаются как свободноживущие во влажной почве, содержащей разлагающийся органический материал, а также в воде.
Типовой вид: Xanthobacter autotrophicus.
Дифференциация видов рода Xanthobacter: См. табл. 4.50.
Таблица 4.50. Дифференцирующие признаки видов рода Xanthobacter 1,2 
Род Xanthomonas
Прямые палочки, обычно 0,4-0,7 x 0,7-1,8 мкм, преимущественно одиночные. Включений поли-β-гидроксибутирата не образуют. Простек и чехлов не имеют. Покоящиеся стадии неизвестны. Грамотрицательные. Подвижные за счет одного полярного жгутика (за исключением вида X. maltophilia, имеющего перитрихиальное жгутикование). Аэробы; метаболизм чисто дыхательного типа с использованием кислорода в качестве конечного акцептора электронов. Нитрат не восстанавливают (за исключением X. maltophilia, восстанавливающего нитрат до нитрита). Оптимальная температура 25-30°С. Колонии обычно желтого цвета, гладкие, маслянистые или клейкие. Образуют очень специфические пигменты: бромзамещенные арильные полнены, или «ксантомонадины» (за исключением X. maltophilia, который не образует ксантомонадины). Оксидазоотрицательные или слабо положительные. Каталазоположительные. Хемоорганотрофы; способны использовать в качестве единственных источников углерода разнообразные углеводы и соли органических кислот. Из многих углеводов образуют в небольших количествах кислоту. В молочке с бромкрезоловым пурпурным или лакмусовом молочке кислоту не образуют, Растут в среде с лактатом кальция, но не глутамнном. Аспарагин в качестве единственного источника углерода и азота не используют. Трифенилтетразолийхлорид в концентрации 0,1% (обычно уже 0,02%) ингибирует рост. Как правило, нуждаются в факторах роста, в том числе метиоиине, глутамнновой кислоте и никотиновой кислоте или их сочетаний. Патогенны для растений; встречаются в ассоциации с растениями (за исключением X. maltophilia, вызывающего оппортунистическую инфекцию у человека).
Типовой вид: Xanthomonas campestris.
Дифференциация видов рода Xanthomonas: См. табл. 4.51.
Таблица 4.51. Дифференцирующие признаки видов рода Xanthomonas 1 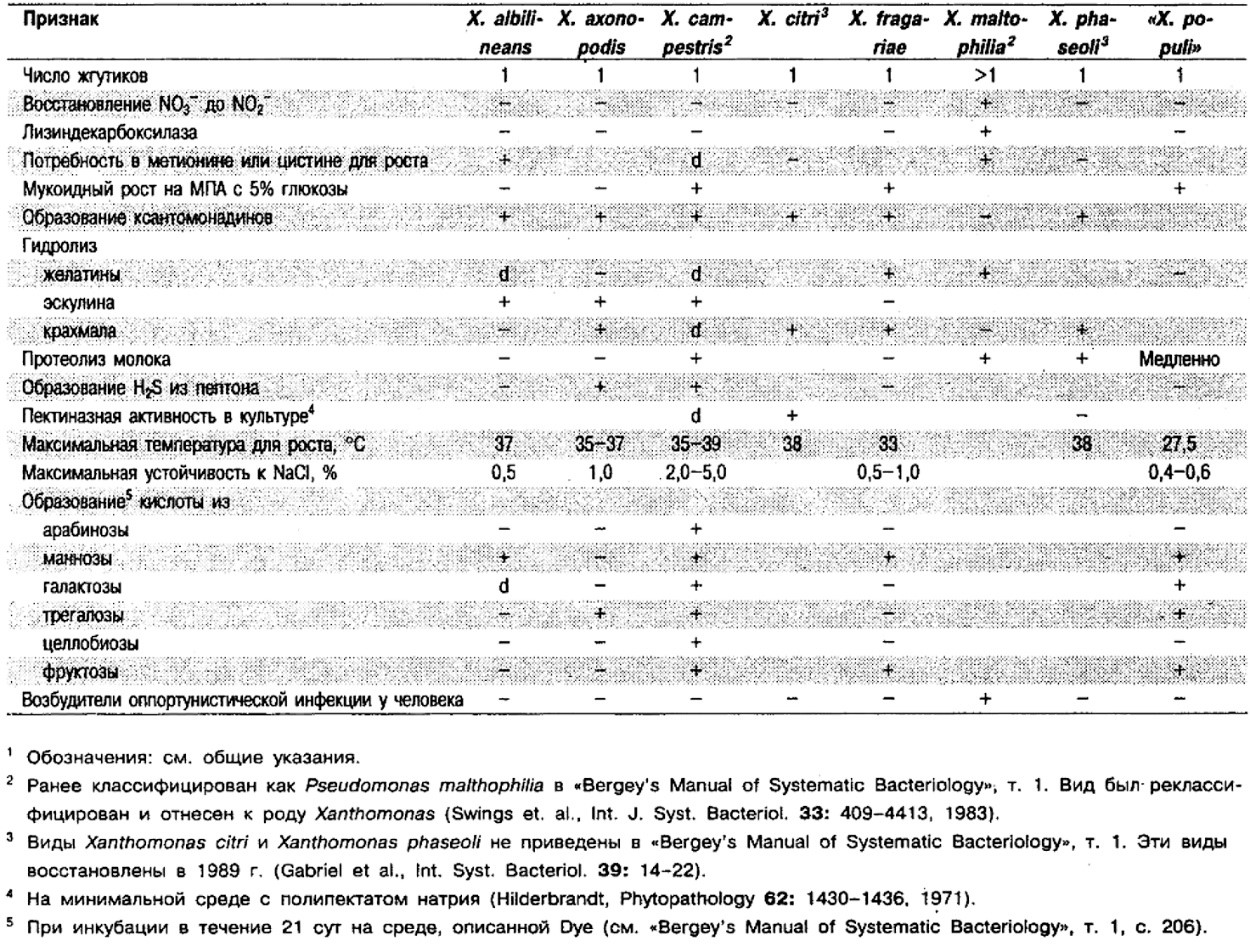
Род Xylella
От редакции: Род Xylella не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1987 г. (Wells et al., Int. J. Syst. Bacteriol 37: 13-143) и включает единственный вид X. fastidiosa.
Одиночные прямые палочки, 0,25-0,35 х 0,9-3,5 мкм, в некоторых условиях культивирования с длинными нитевидными тяжами. Колонии двух типов: от выпуклых до подушечкообразных, гладкие, опалесцирующие, с цельным краем и (другой тип) с центральным возвышением (умбоном) и мелковолнистым краем. Грамотрицательные. Неподвижные. Оксидазоотрицательные и каталазоположительные. Облигатные аэробы; брожение не осуществляют; не галофилы; пигментов не образуют. Высокотребовательны в отношении источников питания; нуждаются для роста в специализированной сложной среде, такой как BCYE с активированным углем или глутамин-пептонная среда (PW) с сывороточным альбумином. Оптимальная температура для роста 26–28°С. Оптимальное значение рН 6,5–6,9. Местообитание – ксилема растений.
Типовой (и единственный) вид: Xylella fastidiosa.
Признаки вида. Гидролизуют желатину, используют гиппурат. Большинство штаммов образует β-лактамазу. Глюкозу не сбраживают. Отрицательные по следующим признакам: индол, H2S, β-галактозидаза, липаза, амилаза, коагулаза и фосфатаза. Вид выделен как фитопатогенный из тканей ряда растений-хозяев. Типовой штамм выделен из лозы винограда, пораженного болезнью. Пирса.
Род Xylophilus
От редакции: Род Xylophilus не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1987 г. (Willems et at., Int J. Syst. Bacteriol. 37: 422-430) и включает единственный вид X. ampelinus (прежнее название – Xanthomonas ampelina).
Палочки, от прямых до слегка изогнутых, 0,4-0,8 х 0,6-3,3 мкм. В старых культурах могут встречаться нитевидные клетки (длиной 30 мкм и более). Клетки располагаются одиночно, в парах или в коротких цепочках. Подвижные за счет одного полярного жгутика. Грамотрицательные. Оксидазоотрицательные, каталазоположительные. Аэробы; хемоорганотрофы. Мета бол изируют углеводы по окислительному пути. Даже при оптимальной температуре 24° С рост обычно очень медленный и слабый. Растут на среде с L-глутамином, но не лактатом кальция (в отличие от штаммов Xanthomonas). Патогенны для растений как возбудители бактериального некроза и рака.
Типовой (и единственный) вид: Xylophilus ampelinus.
Признаки вида: На питательном агаре колонии округлые, полупрозрачные, слегка приподнятые, блестящие, бледно-желтого цвета, с цельным краем. Через 6 сут роста диаметр колоний 0,2-0,3 мм, через 15 сут – 0,6-0,8 мм. Хорошо растут на среде GYAC (Willems et al., 1987) и лучше всего на среде, содержащей 1% дрожжевого экстракта, 2% D-галактозы, 2% СаСО3, и 2% агара, при 24°С. На такой среде колонии окрашены в желтый цвет и выделяют коричневый диффундирующий пигмент. Некоторые штаммы могут образовывать на среде GYCA колонии двух типов. Через 15 сут роста при 24°С средний диаметр колоний быстро- и медленнорастущего типов составляет соответственно 1,4 ±0,6 и 0,7 ± 0,3 мм. Минимальная и максимальная температуры для роста 6 и 30°С соответственно. Максимальная концентрация NaCl, к которой клетки устойчивы, – 1%; очень медленно растут в присутствии 0,01-0,02% 2,3,5- трифенилтетразолийхлорида. Нуждаются для роста в L-глутамате (0,1%); в L-метионине не нуждаются. Образуют кислоту из L-арабинозы и D-галактозы, но не D-ксилозы, D-рибозы, L-рамно- зы, D-глюкозы, D-маннозы, D-фруктозы, сахарозы, трегалозы, целлобиозы, лактозы, мальтозы, D- мелибиозы, рафинозы, гликогена, инулина, декстрина, адонитола, дульцитола, D-маннитола, D-сорбитола, L-сорбозы, салицина, α-метилгликозида, арбутина и мезо-инозитола. Растут за счет использования ацетата (0,2, но не 0,5%), цитрата, DL-малата, сукцината, DL-тартрата и фумарата, но не формиата, пропионата, малоната, малеата, оксалата, бензоата или глюконата кальция. Растут на среде Dye с аспарагином. Желатину, эскулин, крахмал, казеин, арбутин и гиппурат натрия, а также Твин-80 не гидролизуют. Результат теста на развитие мокрой гнили картофеля отрицательный. Образуют H2S из цисте и на и слабо из тиосульфата. Обладают активностью уреазы. По следующим признакам все исследованные штаммы отрицательные: восстановление нитрата, образование индола и аммония, реакция Фогеса-Проскауэра, аргининдишдролаза, лизин- и орнитиндекарбоксилаза, лецитиназа и образование кислоты из глюкозы. Выделены из Vitis vinifera, у которого вызывают бактериальный некроз и рак, главным образом древесных частей, в районах Средиземноморья и в Южной Африке. Имеются сообщения о сходных симптомах заболеваний растений в Швейцарии, Австрии, Болгарии, на Канарских о-вах и в Аргентине.
Род Zoogloea
Толстые палочки, от прямых до слегка изогнутых, 1,0-1,3 x 2,1-3,6 мкм, с закругленными концами; иногда тупоугольнозаостренные на одном или обоих полюсах. Спор и цист не образуют. В старых культурах окружены хорошо заметной капсулой. Грамотрицательные. Активно подвижные, особенно в молодых культурах, за счет одного полярного жгутика. На средах с солями органических кислот образуют внутриклеточные гранулы поли-β-гидроксибутирата. На поздних стадиях роста в жидкой среде культуры имеют вид хлопьев и пленок, клетки погружены в гелеоб разный матрикс и образуют зооглен – структуры с характерной «древовидной» или «пальцевидной» морфологией. Молодые колонии на плотной среде в атмосфере воздуха прозрачные и точечные, однако могут достигать 1-2 мм в диаметре и иметь непрозрачный центр. Клетки не пигментированы. Аэробы; метаболизм чисто дыхательного типа с использованием кислорода в качестве конечного акцептора электронов; возможен также рост в анаэробных условиях в присутствии нитрата (за счет нитратного дыхания). Осуществляют денитрнфикацию с образованием N2. Оптимальная температура для роста 28–37°С, оптимальный диапазон рН 7,0-7,5. Окснщазоположительные. Слабо каталазоположительные. Хемоорганотрофы. Не образуют кислоту из углеводов, за исключением ксилозы, глицерола и этанола, используемых отдельными штаммами. Осуществляют протеолнз желатины. Большинство штаммов уреазоположитсльные. Цвет лакмусового молочка не изменяют. H2S из цистеина обычно не образуют. Основными источниками углерода служат соли некоторых органических кислот (например, лактат, пируват и фумарат), дикарбоновые аминокислоты (например, аспартат, глутамат и аспарагин), спирты и соли ряда ароматических кислот (например, бензоат и м-толуат). Разлагают производные бензола по механизму лдета-расщеплення кольца. Источниками азота служат его органические соединения (дикарбоновые аминокислоты) и аммиак, но не нитрат. Специфические потребности в факторах роста, если они имеются, неизвестны. Встречаются как свободноживущне в загрязненных органическими веществами пресных водах и в сточных водах на всех стадиях очистки.
Типовой (и единственный) вид: Zoogloea ramigera.
Признаки вида: На среде Козера с цитратом не растут. H2S из цистеина или на агаре Клиглера с железом не образуют. Индол не образуют. Образуют аммиак из аспарагина. Обладают липолитической активностью. Растут за счет использования бензоата, м-толуата и о-крезола, осуществляя мета-расщепление ароматического кольца. Метанол, формиат и формальдегид в качестве единственных источников углерода не используют.
Подгруппа 4 В
Дифференциация родов и видов в Подгруппе 4 В: Основные дифференцирующие признаки представлены в табл. 4.52.
Таблица 4.52. Дифференцирующие признаки Taytorella equigenitalis,
Wolinella recta, Bacteroides ureoiyticu? и Bacteroides gracilis1 
Род Taylorella
От редакции: Род Taylorella не приведен в «Веrgey's Manual of Systematic Bacteriology». Этот род описан в 1983 г, (Sugimoto et al., Curr, Microbiol. 9: 155-162; Int. J. Syst. BacterioL 34: 503-504; 1984) и включает единственный вид Т. equigenitalis. Ранее данный вид был приведен в «Bergey's Manual of Systematic Bacteriology» как species incer- tae sedis в составе рода Haemophilus.
Клетки палочковидные, часто приближающиеся по форме к сферическим, 0,7 x 0,7-1,8 мкм, иногда с нитями длиной 5-6 мкм. Грамотрицательные. Неподвижные. Микроаэрофилы: в атмосфере воздуха или в анаэробных условиях растут очень слабо. Наилучший рост наблюдается в присутствии 5-10% СО2. Метаболизм чисто дыхательного типа с использованием кислорода в качестве конечного акцептора электронов. Оптимальная температура 35-37°С; диапазон температуры 30-42°С. Каталазо- и оксндазололожительные. Нитрат и нитрит не восстанавливают. Хемоорганотрофы. Лучше всего растут на шоколадном агаре, однако Х- и V-факторы не стимулируют рост. На обычных средах не растут; на кровяном агаре растут очень слабо или не растут. Кислоту из углеводов не образуют. Обладают фосфатазной и фосфоамидазной активностями. По следующим признакам отрицательные: образование индола и H2S, активности лизин- и орнитиндекарбоксилазы, аргигощдигидролазы, уреазы, желатин азы, липазы и ДНКазы. Паразиты лошадей; встречаются как комменсалы на наружных половых органах здоровых жеребцов, но могут также в течение длительного времени сохранять жизнеспособность в области клитора и вульвы кобыл, вызывая впоследствии инфекционный процесс. Патогенны для кобыл: вызывают эндометрит и цервицит как результат случки с жеребцом-носителем.
Типовой (и единственный) вид: Taylorella equigeni talis.
Признаки вида: Как для рода. См, также табл. 4.52.
Род Wolinella: Wolinella recta
Преимущественно прямые палочки, 0,5-1,0 x 2-6 мкм, с закругленными концами. Спор не образуют. Подвижные за счет одного полярного иоугака. Хотя представители Wolinella recta описаны в «Веrgey's Manual of Systematic Bacteriology», т. 1, как анаэробы, на самом деле это микроаэрофилы, нуждающиеся в Н2 и формиате и способные к кислородному дыханию. В отсутствие альтернативных конечных акцепторов электронов, таких как фумарат, способны к кислород-зависимому росту при низких уровнях кислорода, но не растут ни в анаэробных, ни в аэробных условиях, (Ohta, Gott- schal, FEMS Microbiol. Ecol, 53: 79-86, 1988; Ohta, Gottschal, FEMC Microbiol Lett. 50: 163-168, 1988). Используют фумарат и нитрат как акцепторы электронов для анаэробного дыхания (см. также разд. «Группа 6» в данном определителе). Оксндазоположительные; каталазоотрицательные. Содержат цитохромы b и с. Углеводы не используют. Выделены из воспаленного периодонта у человека.
Отличия Wolinella recta от других грамотрицательных микроаэрофильных палочковидных бактерий: См. табл. 4.51.
Род Bacteroides: Bacteroides ureolyticus и Bacteroides gracilis
Палочки, 0,4-0,5 х 1,4–6,0 мкм. Неподвижные. Хотя представители этих двух видов описаны в «Bergey's Manual of Systematic Bacteriology», т. 1, как анаэробы, на самом деле это микроаэрофилы, нуждающиеся в Н2 и формиате и способные к кислородному дыханию. В отсутствие альтернативных акцепторов электронов, таких как фумарат, растут за счет использования О2 при низких уровнях кислорода, но не растут ни в анаэробных, ни в аэробных условиях (Han et al., Int. Syst. Bacteriol., 41: 218-222, 1991). Используют фумарат и нитрат как акцепторы электронов для анаэробного дыхания (см. также разд. «Группа 6» в данном определителе). Содержат цитохромы b и с. Растут при 25 и 37°С, но не при 42°С. Катала зоотрицате л ьные. Один вид (В, ureolyticus) оксидазоположительный. Углеводы не используют. Образуют H2S в тестовой среде SIM (сульфидиндол–подвижность), а также в конденсационной воде у основания свежеприготовленного скошенкого агара с тремя сахарами и железом. Бактерии В. ureolyticus выделены при инфекциях дыхательных путей и кишечного тракта, из полости рта, кишечника, мочеполовых путей и крови в местах удаления зубов. Бактерии В. gracilis выделены у человека из десневой борозды, а также при инфекциях внутренних органов, головы или шеи и при анаэробных плевро-легочных инфекциях.
Отличия Bacteroides ureolyticus и Bacteroides gracilis от других грамотрицателъных микроаэрофилъ- ных палочковидных бактерий: Си. табл. 4.52.
Необходимые указания для пользования данным определителем
Если в сносках под таблицей мет других пояснений, то используемые обозначения имеют следующий смысл:
«+» – 90% и более штаммов положительные
«–» – 90% и более штаммов отрицательные
«d» – 11-89% штаммов положительные
«v» – нестабильность на уровне штамма (не равнозначно «d»)
«D» – различные реакции у разных таксонов (видов одного рода или родов одного семейства).
Для других обозначений пояснении приведены в сносках под таблицами.
