Клетки от кокковидных до прямых палочек, обычно 0,2–0,4 х 0,4–2,0 мкм. Возможен плеоморфизм с образованием раздутых и нитевидных клеток.
Грамотрицательные. Содержат деметилменахиноны; убихиноны могут синтезироваться или отсутствуют. Неподвижные. Аэробы с более или менее выраженной микроаэрофильностыо, факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура 37°С. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты, однако у наиболее требовательных к составу среды видов соответствующие тесты для определения брожения могут не выявить кислых продуктов брожения. Как правило, не образующие газ, однако встречаются и газообразующие виды. В типе положительные по оксидазе, каталазе и щелочной фосфатазе, однако некоторые виды отрицательные. Восстанавливают нитрат до нитрита. Для первичного выделения рекомендуются сложные среды с добавлением дрожжевого экстракта и сыворотки крови или лизата цельной крови. Нуждаются в органических источниках азота. Разнообразные потребности в питательных веществах могут включать потребность в нескольких аминокислотах, витаминах группы В, β-никотинамиде, адениннуклеотидах и гематине или протопорфнрине. Паразиты позвоночных, в основном млекопитающих и/или птиц.
Типовой род: Pasteurella.
Отличия PasteureUaceae от других семейств и родов, входящих в Группу 5: Признаки, отличающие PasteureUaceae от других семейств и родов, представлены в табл. 5.1.
Дифференциация родов в семействе PasteureUaceae:
От редакции: В «Bergey's Manual of Systematic Bacteriology» указано на гетерогенность каждого рода в семействе PasteureUaceae и обусловленную этим необходимость переопределения всех родов на основе генетических и биохимических критериев. Новые исследования, опубликованные уже после выхода в свет «Bergey's Manual of Systematic Bacteriology», подтвердили эту гетерогенность и привели к перераспределению видов между существующими родами и добавлению новых видов в каждый из существующих родов (Mutters et al., Int. J. Syst. Bacteriol. 35: 309–322, 1985; Schlater et al., J. Clin. Microbiol. 27: 2169-2174, 1989; De Ley et al., Int. J. Syst. Bacteriol. 40: 126–137, 1990; Sneath, Stevens, Int. J. Syst. Bacteriol. 40: 148–153, 1990). Судя по данным, полученным в этих исследованиях, следует, возможно, ввести по крайней мере 4 дополнительных рода, но при этом трудно составить фенотипические диагнозы как новых, так и установленных родов.
Учитывая, что в дальнейшем будут проведены исследования по установлению и определению родов в семействе PasteureUaceae, мы не делаем попытки дифференцировать роды. В табл. 5.53 приведены биохимические свойства каждого вида этого семейства. Для видов, которые предложено перенести в тот или другой род, использовано в каждом случае новое родовое название (это не вызвано предвзятым мнением), а старое дано в скобках. Следует отметить, что оба названия узаконены.
Таблица 5.53. Биохимические признаки видов семейства Pasteurellaceae 1,2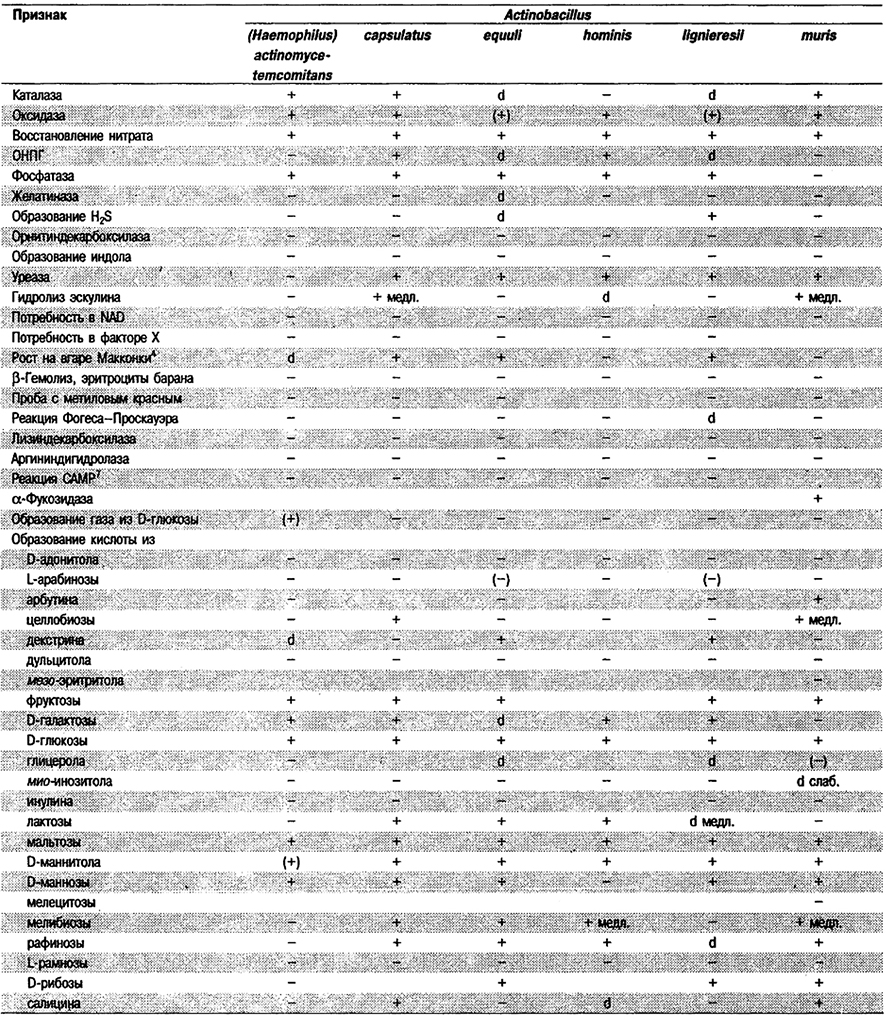
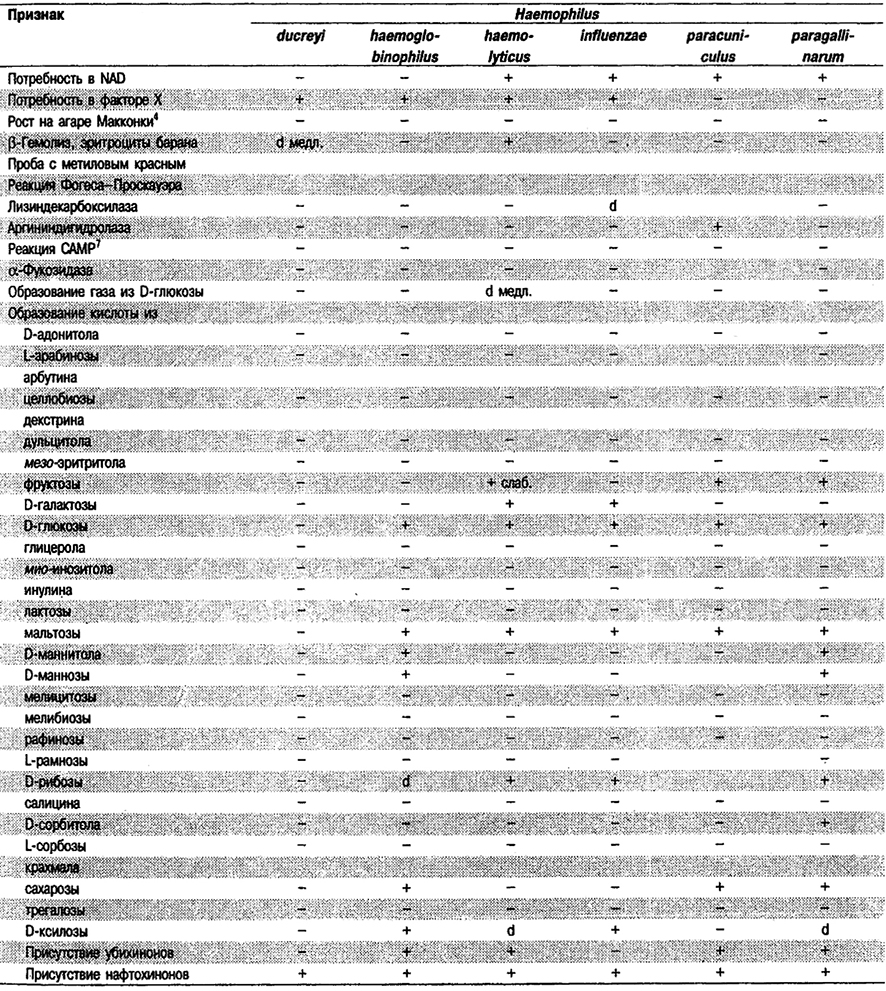
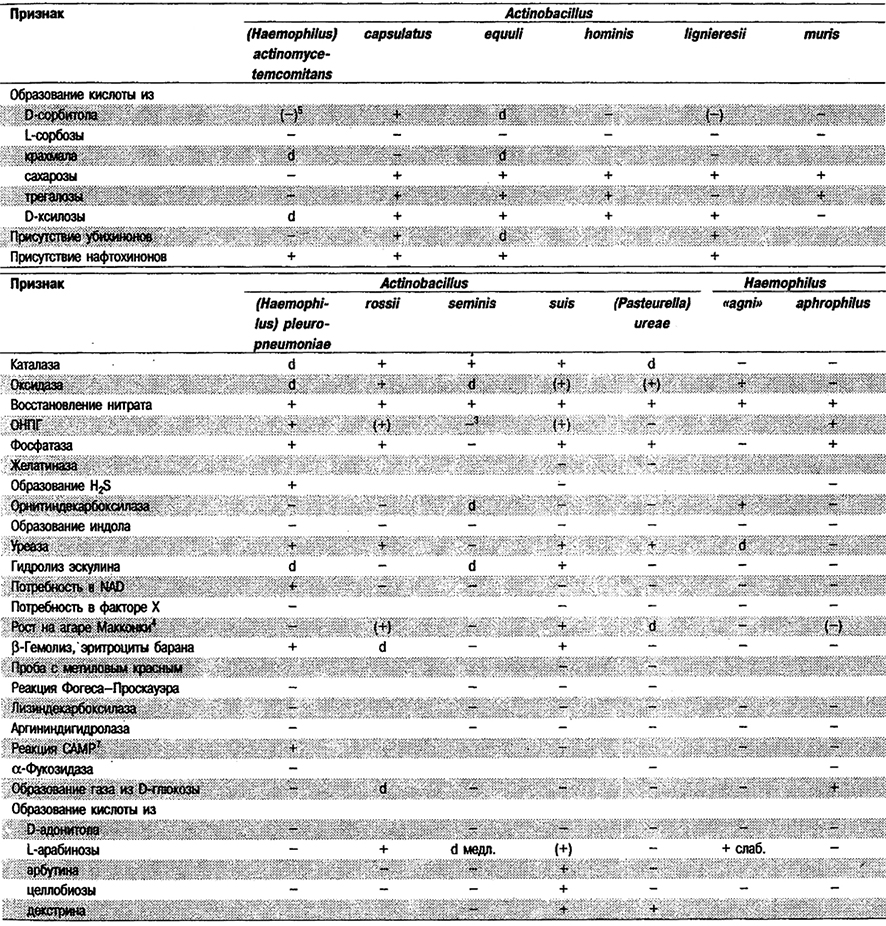
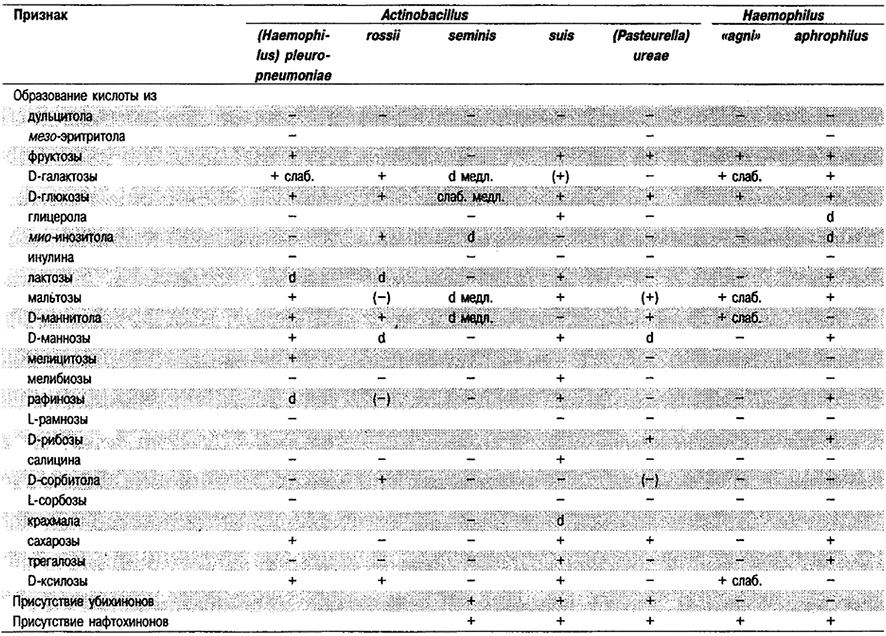
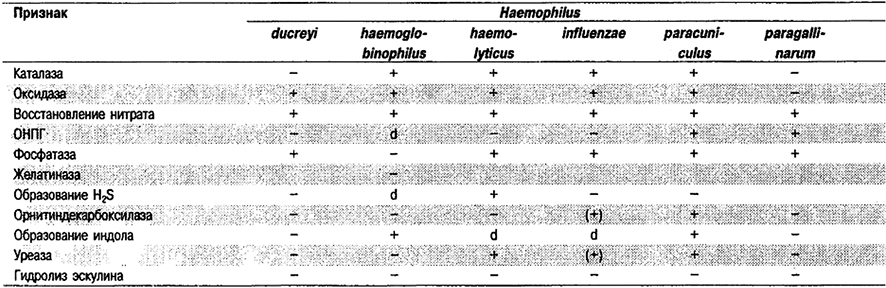
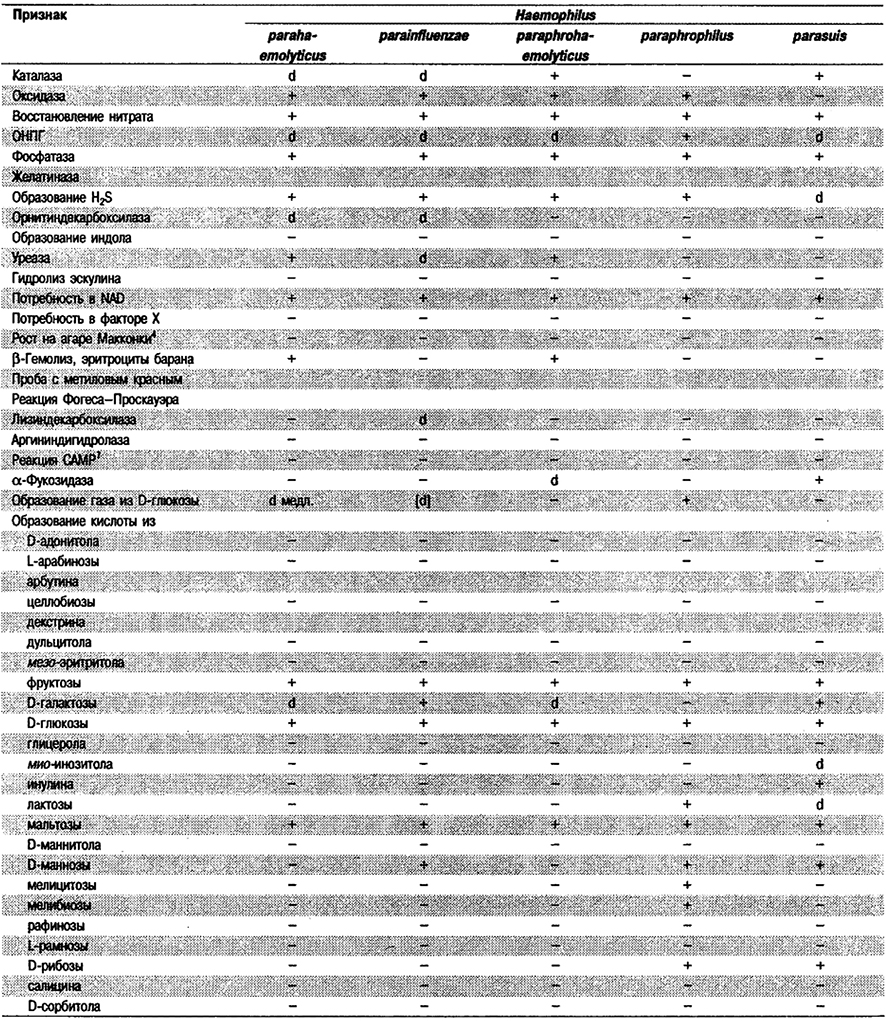
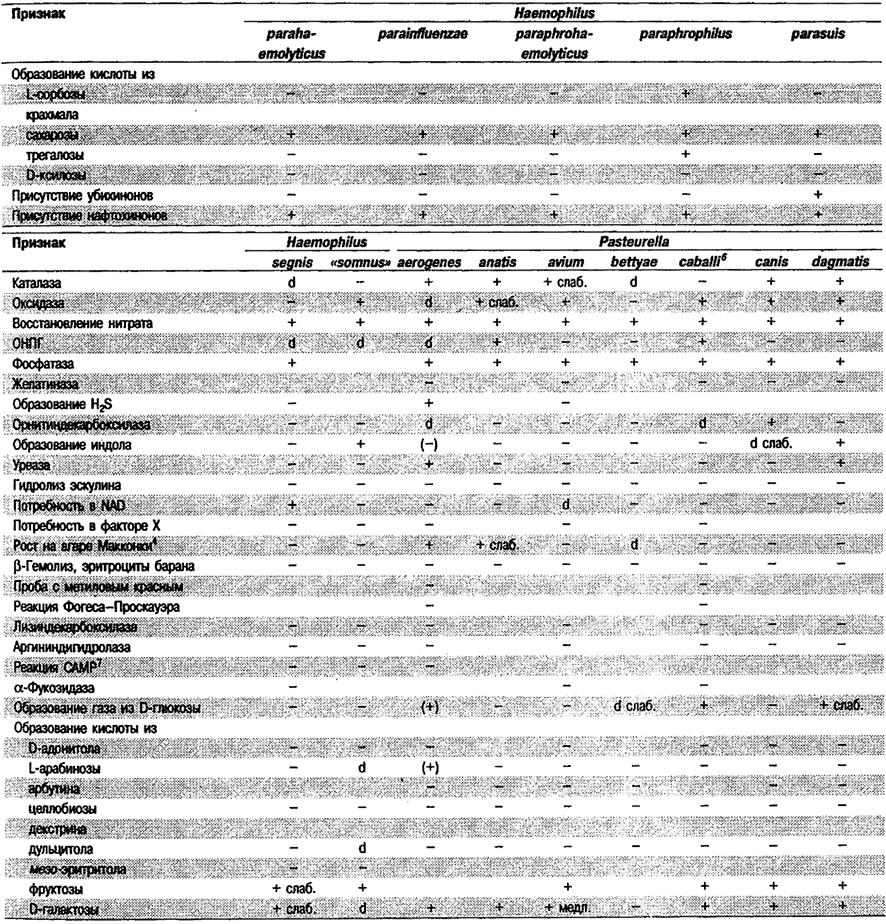
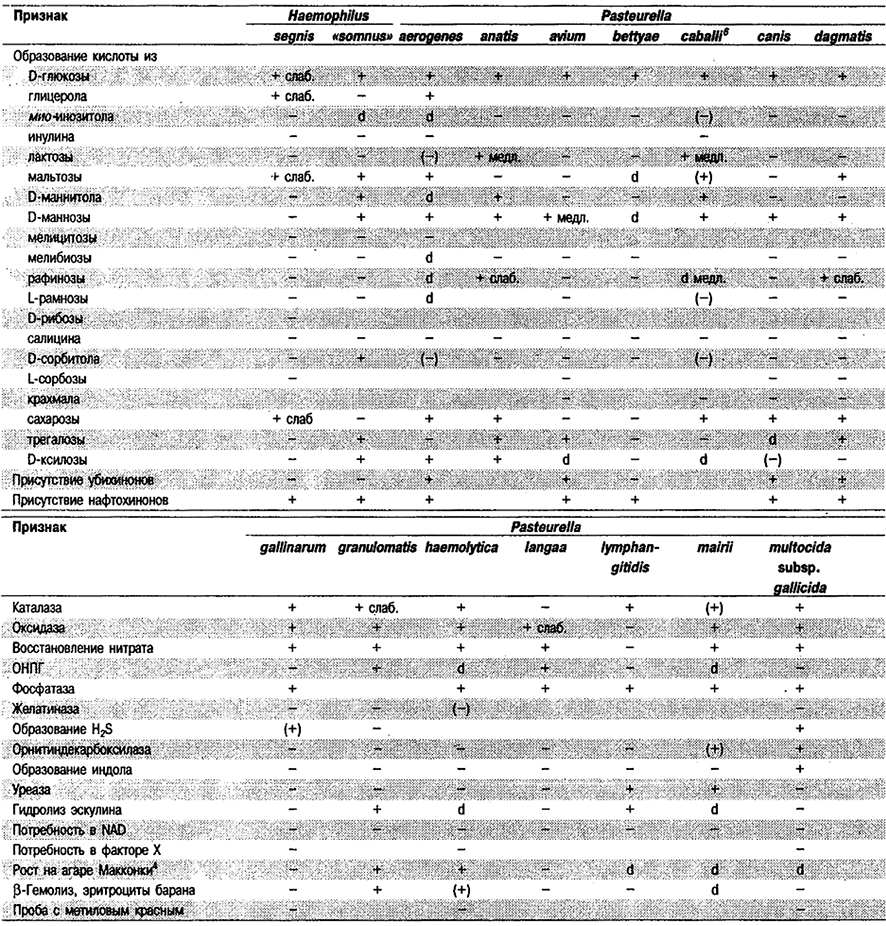
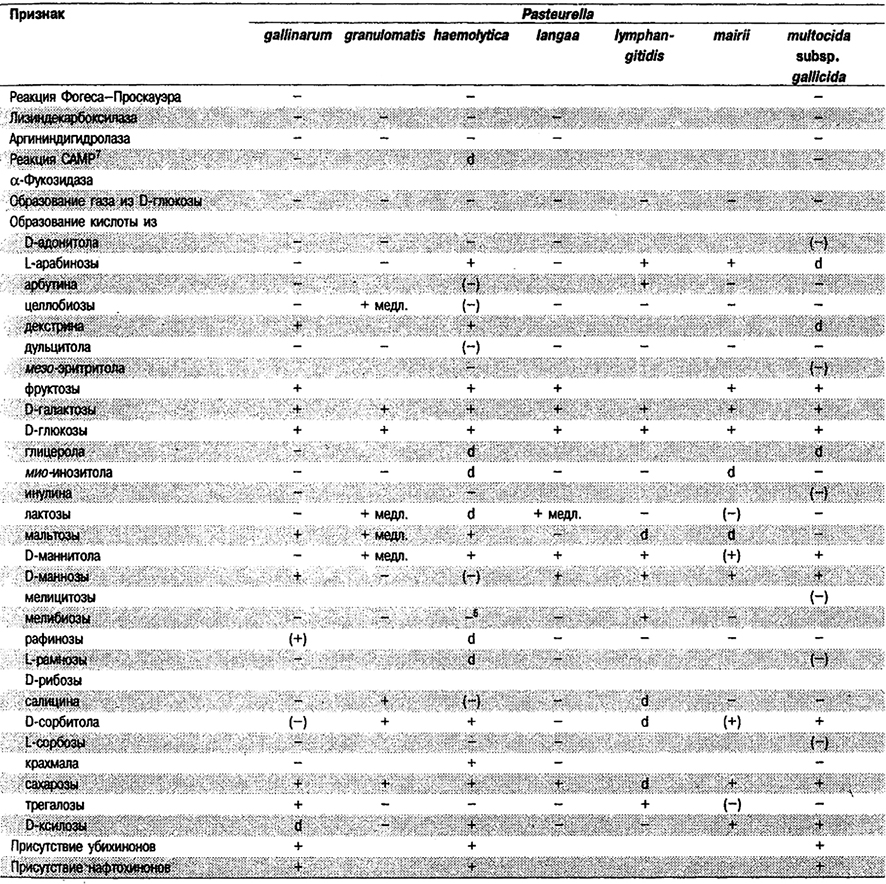
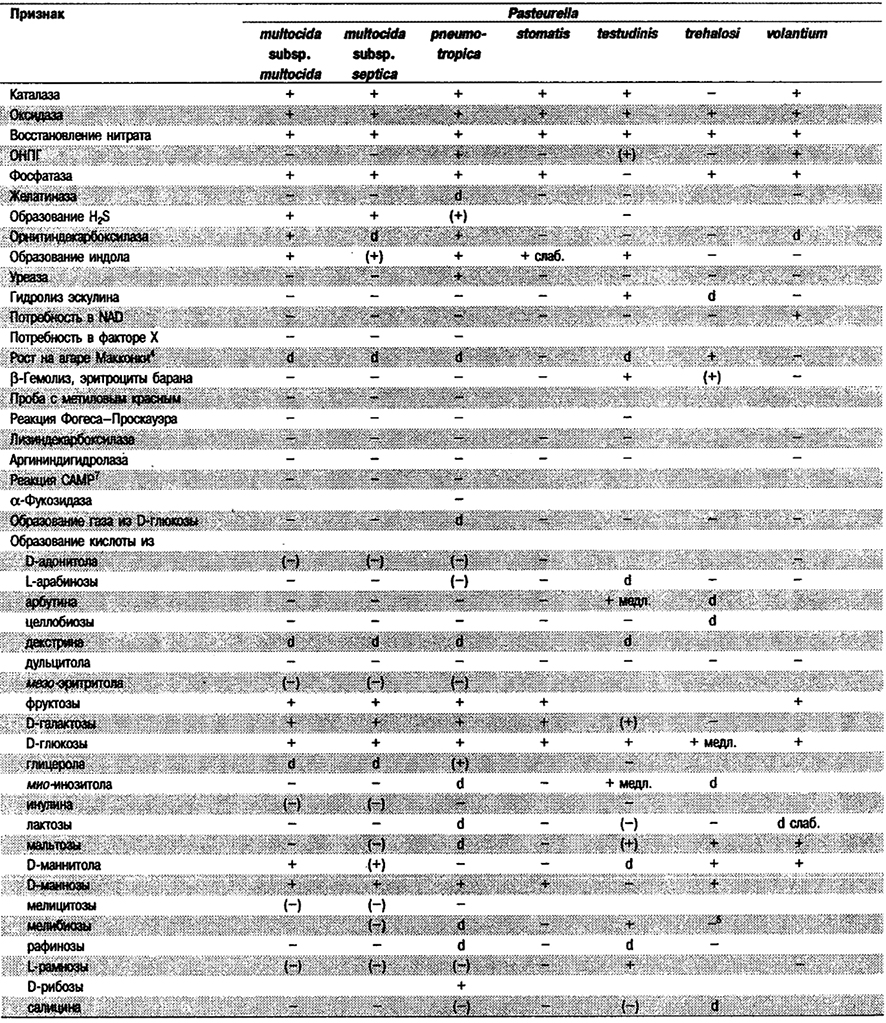
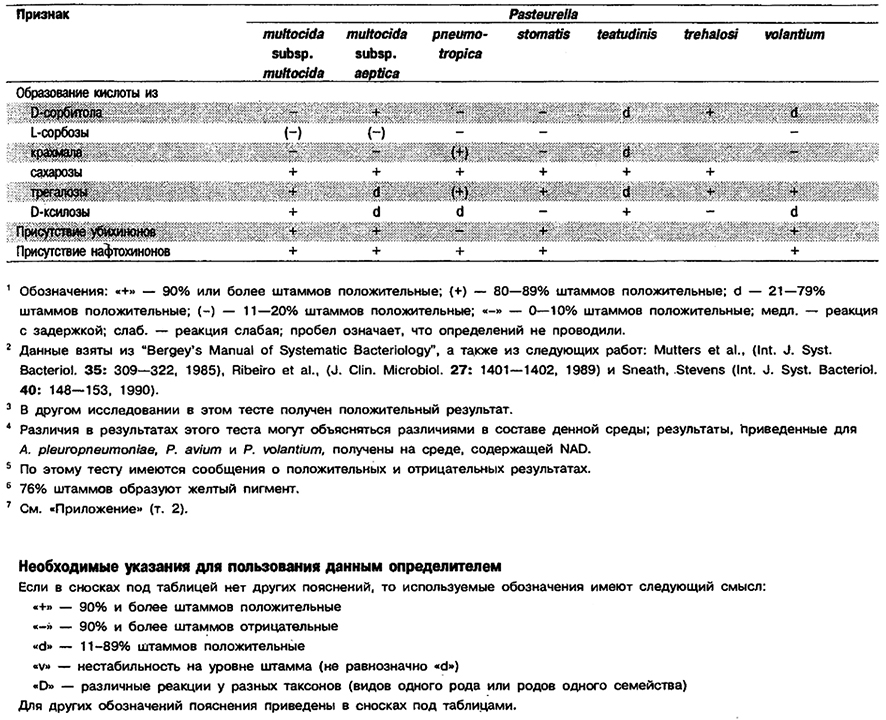
Род Actinobacillus
Клетки сферические, овальные или палочковидные, 0,4 х 1,0 мкм, в большинстве бациллярные, но с примесью кокковых элементов, которые часто расположены у полюса палочек, что дает характерную картину знаков азбуки Морзе. Иногда встречаются удлиненные формы (до 6 мкм), особенно при росте на средах, содержащих глюкозу или мальтозу. Клетки одиночные, в парах или, реже, в цепочках. Грамотрицательные. Окрашиваются неравномерно. Во влажных препаратах, контрастированных тушью, может быть выявлено небольшое количество внеклеточной слизи. Неподвижные. Культуры в процессе первичного выделения очень клейкие; колонии бывает трудно снять полностью с поверхности агара. Культуры, растущие на поверхности субстрата, маложизнеспособны и погибают за 5–7 сут. Факультативные анаэробы и хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура 37°С. Катаболизируют D-глюкозу и фруктозу с образованием кислоты, но не газа. Положительные по β-галактозидазе. Пробы с метиловым красным и на образование индола отрицательные. Паразиты или комменсалы человека, овец, крупного рогатого скота, лошадей, свиней, других млекопитающих и птиц.
Типовой вид: Actinobacillus lignieresii.
Дифференциация видов рода ActinobaciUus. См. табл. 5.54. Другие признаки видов приведены в табл. 5.53.
Определенная трудность состоит в различении актинобацилл и представителей родов Haemophilus и Pasteurella из-за их близкого сходства. Обычно требуется провести сравнение биохимических признаков на видовом уровне (табл. 5.53).
Таблица 5.54. Дифференциация видов Actinobacillus (Haemophilus/Pasteurella) 1,2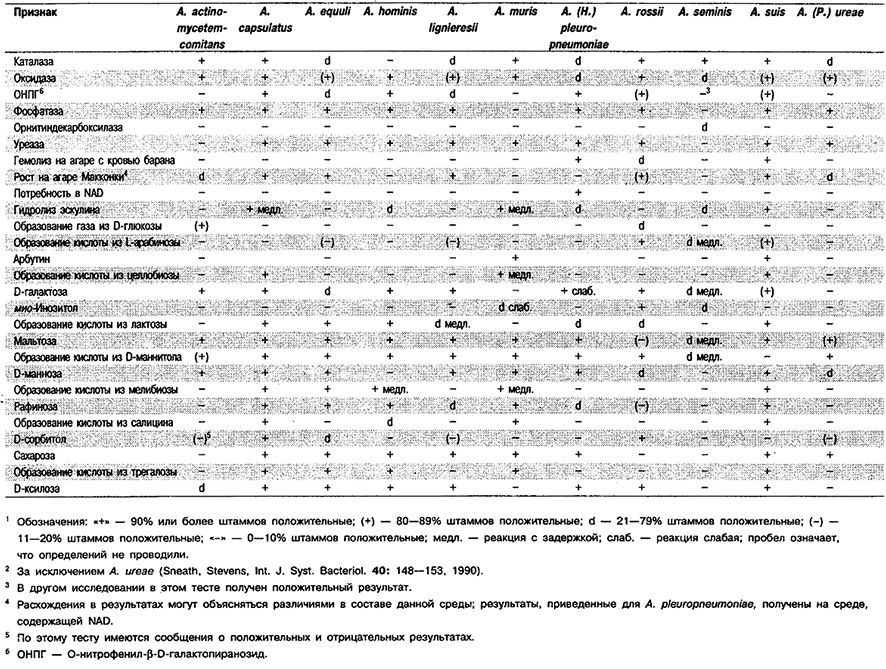
От редакции: Виды Actinobacillus ho minis, Actino- bacillus muris и Actinobacillus rossii не приведены в «Bergey's Manual of Systematic Bacteriology»; вид Actinobacillus (Haemophilus) pleuropneumoniae внесен в род Haemophilus; Actinobacillus seminis упомянут как вид insertae sedis, название которого не узаконено; вид Actinobacillus (Pasteurella) ureae внесен в род Pasteurella. Вищ Actinobacillus hominis описан в 1981 г. (Friis–Moller, In: Kilian, Frederiksen, Biberstein (Ed.), Haemophilus, Pasteurella and Actinobacillus, Academic Press, London, 151–157; Int. J. Syst. Bacteriol. 35: 375–376, 1985); вид Actinobacillus muris описан в 1986 г, (Bisgaard, Acta Pathol. Microbiol. Immunol. Scand. Sect. B. 94: 1– 18; Int. J. Syst. Bacteriol. 38: 220-222, 1988); вид Actinobacillus pleuropneumoniae перенесен в род Actinobacillus из рода Haemophilus в 1983 г. (Pohl et al., Int. J. Syst. Bacteriol 33: 510–514); виды Actinobacillus rossii и Actinobacillus seminis описаны в 1990 г. (Sneath, Stevens, Int. J. Syst. Bacteriol. 40: 148–153); вид Actinobacillus ureae перенесен в род Actinobacillus из рода Pasteurella в 1986 г. (Mutters et al., Int. J. Syst. Bacteriol. 36: 343–344).
Род Haemophilus
Клетки сферические, овальные или палочковидные, по размерам от точечных до средних, обычно менее 1 мкм в ширину и различные по длине, иногда образуют нити и обнаруживают заметный плеоморфизм. Грамотрицательные. Неподвижные. Факультативные анаэробы. Почти все виды нуждаются в готовых факторах роста, присутствующих в крови, особенно в факторе X (протопорфирин IX, или протогем) и/или факторе V [никотинамидадениндинуклеотид (NAD) или NAD-фосфат (NADP)]. Даже при наличии факторов роста лучше растут на средах сложного состава. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура 35–37°С. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты; небольшое число – газообразующие. Восстанавливают нитрат до нитрита или далее. По результатам тестов на оксидазу и каталазу виды различаются. Встречаются как облигатные паразиты или комменсалы на слизистых оболочках у человека и разнообразных животных. Н. influenzae – основной возбудитель менингита у детей, а также вызывает септицемию, воспаление среднего уха, синусит и хронические бронхиты. Н. influenzae биовар aegyptius главным образом ответствен за конъюнктивит, а некоторые его штаммы – за недавно описанное заболевание красная бразильская лихорадка (Brenner et al., J. Clin. Microbiol. 26: 1524-1534, 1988). H ducreyi – возбудитель венерического заболевания мягкий шанкр, или шанкроид.
Типовой вид: Haemophilus influenzae.
Потребность в факторах роста не служит определяющим признаком Haemophilus, поскольку не все виды Haemophilus нуждаются в факторах X и/или V, а некоторые виды Actinobacillus и Pasteurella также нуждаются в NAD. Чтобы отличить Haemophilus от других родов семейства Pasteurellaceae, как правило, необходимо сравнить биохимические признаки на видовом уровне (табл. 5.53).
Дифференциация видов рода Haemophilus. См. табл. 5.53. Дифференциация биоваров Н. influenzae и биоваров Н. parainfluenzae представлена в табл. 5.55.
Таблица 5.55. Дифференциация биоваров Haemophilus influenzae и Н. Parainfluenzae1
От редакции: В ряде видов, приведенных в «Вегgey's Manual of Systematic Bacteriology» в составе рода Haemophilus, произошли изменения номенклатуры. Установлено, что Haemophilus aegyptius – это тот же вид, что Haemophilus influenzae (Casin et al, Ann. Microbiol. (Paris) 137B: 155–163, 1986). В данном определителе мы рассматриваем его как биовар H influenzae. Вид Haemophilus equigenitalis перенесен в новый род Taylorella (Sugimoto et al., Curr. Microbiol. 9: J55–162, 1983; Int. J. Syst. Bacteriol 34: 503–504, 1984). Вид Haemophilus pleuropheumoniae перенесен в род Actinobacillus (Pohl et al., Int. J. Syst. Bacteriol. 33: 510–514, 1983). Виды «Haemophilusagni» и «Haemophilus somnus» не узаконены; оба они приведены в «Bergey's Manual of Systematic Bacteriology» как виды incertae sedis. Установлено (De Ley et al., Int. J. Syst. Bacteriol. 40: 126–137, 1990), что «Histophilus ovis», другое неузаконенное название, – это старший синоним «Н. agni», и высказано предположение, что как «H. agni», так и «Н, somnus» следует перенести в род «Histophilus».
Род Pasteurella
Клетки сферические, овальные или палочковидные, 0,3–1,0 х 1,0–2,0 мкм, одиночные, реже в парах или коротких цепочках. Обычно окрашиваются биполярно, особенно в препаратах, приготовленных из инфицированных тканей животных. Грамотрицательные. Неподвижные. Факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура 37°С. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты, но обычно не газа. Большинство видов оксидазо- и каталазоположительные. Восстанавливают нитрат до нитрита (за исключением P. lymphangitidis). Пробы с метиловым красным и на образование ацетоина (Фогеса-Проскауэра) отрицательные. По лизиндекарбоксилазе, аргининдигидролазе и желатиназе отрицательные. Паразиты или комменсалы на слизистых оболочках верхних дыхательных путей и пищеварительного тракта у млекопитающих (редко человека) и птиц. P. multocida вызывает геморрагическую септицемию крупного рогатого скота, холеру птиц и пневмонию у сельскохозяйственных животных. P. haemolytica вызывает пневмонию и «транспортную лихорадку» у крупного рогатого скота, овец и коз.
Типовой вид: Pasteurella multocida.
Из-за частичных совпадений и исключений из общих правил в фенотипических описаниях Pasteurella, Haemophilus и Actinobacillus дифференциация этих родов существенно затруднена. Лучше всего проводить определение непосредственно на видовом уровне (табл. 5.53).
Дифференциация видов рода Pasteurella: См. табл. 5.53.
От редакции: В «Bergey's Manual of Systematic Bacteriology» вид Actinobacillus (Pasteurella) ureae включен в род Pasteurella, a Pasteurella (Haemophilus) avium – в род Haemophilus. После выхода в свет этого издания были предложены новые комбинации для Pasteurella avium (Mutters et al., Int. J. Syst. Bacteriol. 35: 5–9, 1985) и Actinobacillus ureae (Mutters et al., Int. J. Syst. Bacteriol. 36: 343–344, 1986). Виды и подвиды Pasteurella anatis, Pasteurella bettyae, Pasteurella caballi, Pasteurella canis, Pasteurella dagmatis, Pasteurella grqnulomatis, Pasteurella langaa, Pasteurella lymphangitidis, Pasteurella mairiit Pasteurella multocida subsp. gallicida, Pasteurella multocida subsp. multocida, Pasteurella multocida subsp. septica, Pasteurella stomatis, Pasteurella testudinis, Pasteurella tregalosi и Pasteurella volantium не приведены в «Bergey's Manual of Systematic Bacteriology». Pasteurella anatis, Pasteurella canis, Pasteurella dagmatis, Pasteurella langaa, Pasteurella multocida subsp. gallicida, Pasteurella multocida subsp. multocida, Pasteurella multocida subsp. septica и Pasteurella stomatis описаны в 1985 г. (Mutters et al., Int. J. Syst. Bacteriol. 35: 309–22). Pasteurella bettyae, Pasteurella lymphangitidis, Pasteurella mairit и Pasteurella trehalosi описаны в 1990 г. (Sneath, Stevens, Int. J. Syst. Bacteriol. 40: 148–153; см. Sneath, Int. J. Syst. Bacteriol. 42: 658–659, 1992 относительно поправки орфографии эпитетов bettyae и mairii). Вид Pasteurella caballi описан в 1989 г, (Schlater et. al., J. Clin. Microbiol. 27: 2169–2174; Int. J. Syst. Bacteriol. 40: 320-321, 1990), Pasteurella gmnulomatis – в 1989 г. (Ribeiro et al., J. Clin. Microbiol. 27: 1401–1402; Int. J. Syst. Bacteriol. 32: 105–106, 1982), Pasteurella testudinis – в 1982 г. (Snipes, Bi- berstein, Int. J. Syst. Bacteriol. 32: 201–210). Pasteurella volantium – в 1985 г. (Mutters et al., Int. J. Syst. Bacteriol. 35: 5–9).
