Прямые или изогнутые палочки, 0,3–1,3 х 1,4–5,0 мкм. Грамотрицательные. Подвижные за счет полярных жгутиков или неподвижные (род Enhydrobacter). При росте на плотных средах возможно образование дополнительных латеральных жгутиков. Факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Большинство видов хорошо растет при 37°С, но фотобактерии и некоторые морские вибрионы для роста нуждаются в температуре 25°С или ниже. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты и (некоторые виды) – газа. Большинство оксидазоположительные (исключение составляют Vibrio metschnikovii, V. gasogenes, Photobacterium phosphoreum, большинство P. angustum и некоторые P. leiognathi). Фотобактерии и большинство видов Vibrio для оптимального роста нуждаются в присутствии в среде 2–3% NaCl. Распространены повсеместно; преимущественно водные организмы, обнаруживаемые в морской и пресной воде, а также в ассоциациях с водными животными. Несколько видов патогенны для человека, рыб, угреобразных и лягушек, а также для других позвоночных и беспозвоночных. Vibrio cholerae – возбудитель холеры. Ряд видов Vibrio и Aeromonas, а также Plesiomonas вызывают желудочно-кишечные и раневые инфекции, а также септицемию у человека.
Типовой род: Vibrio.
От редакции: Уже после выхода в свет «Bergey's Manual of Systematic Bacteiology» было предложено изъять Aeromonas и Plesiomonas из семейства Vibrionaceae и перенести Aeromonas в новое семейство Aeromonadaceae, a Plesiomonas – в семейство Enterobacteriaceae (Colwell et al., Int. J. Syst. Bacteriol. 36: 473-477, 1986; MacDonnell, Colwell, Syst. Appl. Microbiol. 6: 171–182, 1985). В данном определителе в соответствии с его назначением оба рода оставлены в семействе Vibrionaceae, что, однако, не связано с оценкой правомерности указанных предложений.
Отличия Vibrionaceae от других семейств и родов, входящих в Группу 5: Признаки, отличающие Vibrionaceae от других семейств и родов, представлены в табл. 5.1. Для родов семейства Enterobacteriaceae характерно отсутствие в клетках окси- дазной активности и то, что подвижность, если она имеется, обусловлена перитрихиальным жгу- тикованием, – эти признаки отличают их от Vibrionaceae. Однако у Tatumella ptyseos (сем. Enterobacteriaceae) жгутики латеральные; кроме того, по крайней мере у двух видов Vibrio (V. metschnikovii и V. gasogenes) отсутствует оксид аза, и некоторые штаммы других видов могут быть оксидазоотрицательными или слабо положительными.
Дифференциация родов семейства Vibrionaceae: См. табл. 5.46.
Таблица 5.46. Дифференциация родов семейства Vibrionaceae 1
От редакции: Для видов, ранее входивших в род Vibrio, предложены два новых рода – Listonella (MacDonell, Colwell, Syst. Appl. Microbiol. 6: 171– 182, 1985; Int. J. Syst. Bacteriol. 36: 354, 1986) и Colwellia (Deming et al. Syst Appl. Microbiol. 10: 152–160, 1988; Int. J. Syst. Microbiol. 38: 328, 1988). В данном определителе, в соответствии с его назначением, все эти виды оставлены в роде Vibrio, что не означает оценку указанных предложений. Новые родовые названия приведены в скобках во избежание путаницы.
Род Aeromonas
Клетки от прямых палочек с закругленными концами до сферических, 0,3–1,0 х 1,0–3,5 мкм, одиночные, в парах или коротких цепочках. Грамотрицательные. Как правило, подвижные за счет единственного полярного жгутика (в молодых культурах на плотной среде могут образовываться перитрихиальные жгутики). Факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальный диапазон температуры 22–28°С; большинство видов хорошо растет при 37°С, за исключением некоторых штаммов. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты и часто газа. Оксидазоположительные и каталазоположительные. Обычно положительны по артининдигидролазе и отрицательны по орнитиндекарбоксилазе. Результаты тестов на уреазу и фенилаланиндезаминазу отрицательные. По ДНКазе и желатин азе положительные. Восстанавливают нитрат. Почти все штаммы сбраживают углеводы, включая мальтозу, D-галактозу и трегалозу. Устойчивы к внбриостатическому агенту 2,4-диамино-6,7-диизопропилптеридину (О/129). Встречаются в пресной воде и сточных водах. Некоторые виды патогенны для лягушек, рыб и человека; вызываемые у человека заболевания – как правило, диарея или бактериемия.
Типовой вид: Aeromonas hydrophila.
Признаки, отличающие Aeromonas от других родов, приведены в табл. 5,46 и 5.47.
Таблица 5.47. Отличия Aeromonas от Plesiomonas1
Дифференциация видов рода Aeromonas: Новые виды Aeromonas трудно отличать от хорошо известных A. caviae, A. hydrophila, A. salmonicida и A. sobria. В связи с этим большинство клинических лабораторий будет сообщать о выделенных штаммах как о представителях группы A. caviae, которая включает A. caviae, A. eucrenophila и A. media; группы A. hydrophila, которая включает A. hydrophila и биогруппу подвижных организмов в составе A. salmonicida; группы A. sobria, которая включает А. sobria и A. veronii; или A. schubertii. Биохимические признаки видов Aeromonas перечислены в табл. 5.48. Следует отметить также, что практически все клинические изоляты, идентифицированные как A. sobria, в действительности представляют собой A. veronii (Hickman-Brenner et al., J. Clin. Microbiol. 25: 900-906, 1987). Строго говоря, A. sobria почти никогда не обнаруживается в клиническом материале.
Таблица 5.48. Дифференциация видов рода Aeromonas1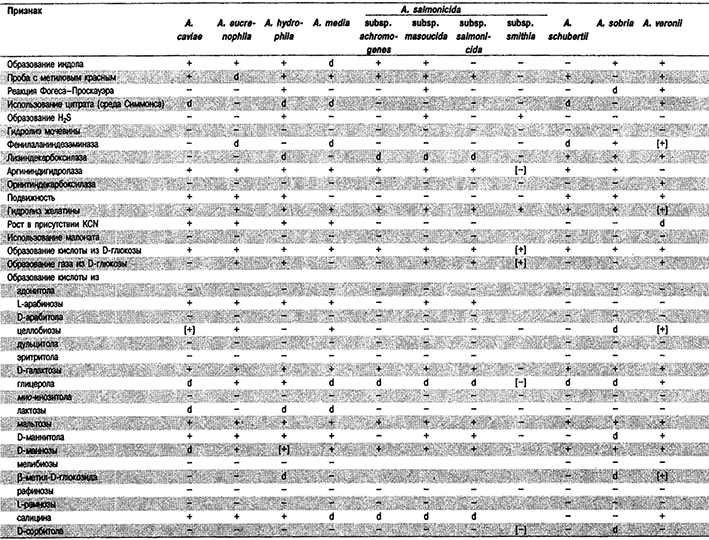
От редакции: Виды Aeromonas eucrenophilat Aero- monas media, Aeromonas salmonicida subsp. smithia, Aeromonas schubertii и Aeromonas vervnii не приведены в «Bergey's Manual of Systematic Bacteriology». Вид Aeromonas eucrenophila описан в 1988 г. (Schubert, Hehazi, Zentralbl. Bakteriol. Parasitenkd, Infektionskr. Hyg. Abt. 1 Orig. Reihe A 268: 34–39; Int. J. Syst. Bacteriol. 38: 449, 1988). Вид Aeromonas media описан в 1983 г. (Allen et al, Int. J. Syst. Bacteriol. 33: 599–604). Подвид Aeromonas salmonicida subsp. smithia описан в 1989 r. (Austin et al., Syst, AppL Microbiol. 11: 277–290; Int. J. Syst. Bacteriol. 39: 495–497, 1989). Вид Aeromonas schubertii описан в 1988 г. (Hickman–Brenner et al., J. Clin. Microbiol. 26: 1561–1564; Int. J. Syst. Bacteriol. 39: 205–206, 1989). Вид Aeromonas veronii описан в 1987 г. (Hickman–Brenner et al., J. Clin. Microbiol. 25: 900–906, Int. J. Syst. Bacteriol. 38: 220-222, 1988).
Род Enhydrobacter
От редакции: Род Enhydrobacter не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1987 г. (Staley et al., Int. J. Syst. Bacteriol. 37: 289–291) и включает один вид Е. aerosaccus.
Очень короткие прямые палочки, 0,5–0,7 х 1,0–5,0 мкм, достигающие сферической формы. Содержат газовые вакуоли. Клетки одиночные, в ларах и коротких цепочках. Грамотрицательные. Неподвижные; факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура для роста 37–39°С. Характерен очень медленный рост; результаты биохимических тестов проявляются в период до 60 сут. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты (о выделении газа сообщений нет). Оксидазо- и каталазоположительные, индолотрицательные. Не растут в среде Фогеса-Проскауэра с метиловым красным. По лизин- и орнитиндекарбоксилазе и аргининдигидролазе положительные. Восстанавливают нитрат. Устойчивы к вибриостатическому агенту О/129. Выделены из бедной кислородом зоны эвтрофного озера.
Типовой (и единственный) вид: Enhydrobacter aerosaccus.
От редакции: Организм высокотребователен к составу питательной среды и растет очень медленно. Для определения его биохимических признаков требуются необычные среды и срок инкубации 30–60 сут. Выделен только однажды; не рекомендуется для изучения студентам.
Колонии на агаризованной среде RM-2 (Staley et al., Int. J. Syst. Bacteriol. 37: 289–291, 1987) бесцветные, выпуклые, с цельным краем, прикрепленные к поверхности агара. Для получения роста на синтетических минеральных средах необходимо вносить фолиевую кислоту и биотин. Наилучший рост наблюдается в микроаэробных условиях. На среде с дезоксирибонуклеазой не растут; не гидролизуют О-нитрофенил-β-О-галактопиранозид. Гидролизуют Твин-80. В качестве источников углерода используют ацетат, L-арабинозу, цитрат, этанол, D-фруктозу, формиат, L-фукозу, фумарат, D-галактозу, D-глюкозу, глицерол, инулин, лактат, мезо-малат, мальтозу, D-маннозу, оксалоацетат, пируват, L-рамнозу, D-рибозу, L-copбозу, сукцинат, сахарозу, мезо-тартрат и D-ксилозу. Не используют следующие источники углерода: адонитол, агар, амигдалин, бензоат, бутират, декстрин, дульцитол, желатину, мезо-инозитол, лактозу, малонат, D-маннитол, мелецитозу, мелибиозу, метанол, пектин, салицин, D-сорбитол, крахмал, рафинозу, трегалозу, фталат, формиат, хитин, целлобиозу, целлюлозу и эритритол. Реакции, по которым отличают Е. aerosaccus от других родов сем, Vibrionaceae, приведены в табл. 5.46.
Род Photobacterium
Толстые прямые палочки, 0,8–1,3 х 1,8–2,4 мкм. В старых культурах или при неблагоприятных условиях культивирования, как правило, присутствуют инволюционные формы. Грамотрицательные, Подвижные за счет 1–3 полярных неочехленных жгутиков; некоторые неподвижные. Факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура, по-видимому, 18–25°С, Для роста нуждаются в ионах натрия. Катаболизируют D-глюкозу и D-маннозу с образованием кислоты и в случае P. phosphoreum газа. Оксидазная реакция вариабельная. Большинство штаммов положительные по лизиндекарбоксилазе и аргининдигидролазе. Отрицательные по орнитиндекарбоксилазе. В некоторых условиях культивирования накапливают поли-β-гидроксибутират; экзогенный мономерный β-гидроксибутират не используют. Большинство штаммов растет в минеральной среде на основе морской воды, в присутствии D-глюкозы и NH4C1; другие штаммы нуждаются, кроме того, в L-метионине. Используют, кроме D-глюкозы, D-фруктозу, глицерол и D-маннозу. Два вида – светящиеся бактерии. Источники выделения – морская среда, а также поверхность и содержимое кишечника морских животных; некоторые представители обнаружены как симбионты в специализированных светящихся органах морских рыб.
Типовой вид: Photobaclerium phosphoreum.
Признаки, отличающие Photobacterium от других родов, приведены в табл. 5.46.
Дифференциация видов рода Photobacterium: См. табл. 5.49А, Другие признаки этих видов приведены в табл. 5.49Б.
Таблица 5.49 А. Дифференциация видов рода Photobacterium 1,2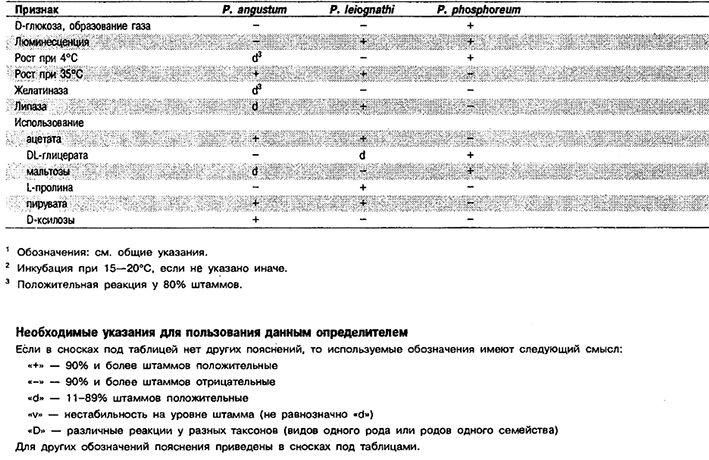
Таблица 5.49 Б. Другие биохимические признаки видов рода Photobacterium1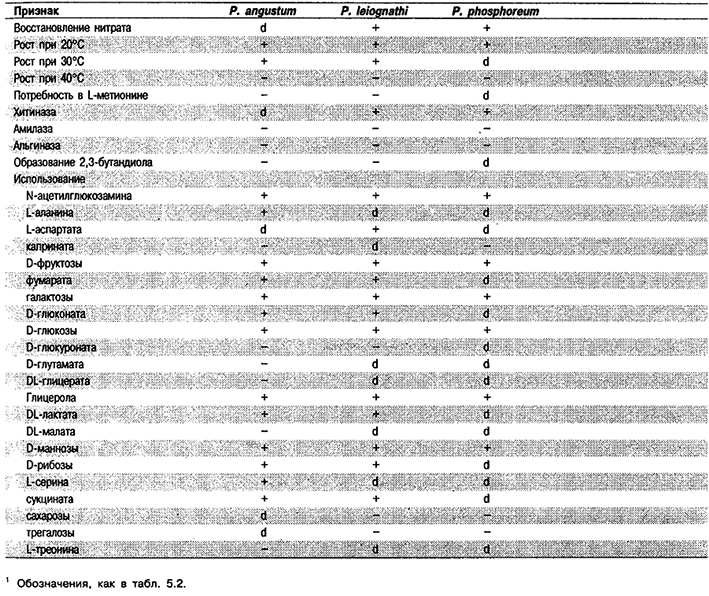
Род Plesiomonas
От редакции: Представителей Plesiomonas рекомендовано перенести в род Proteus (MacDoneil, Colwell, Syst. Appl. Microbiol. 6: 171-182, 1985) сем, Enterobacteriaceae, поскольку их 5S-pPHK близкородственна таковой Proteus mirabilis. Подобное изменение создало бы проблемы в фенотипическом определении рода Proteus.
Прямые палочки с закругленными концами, 0,8–1,0 х 3,0 мкм. Грамотрицательные. Подвижные за счет полярных жгутиков, обычно лофотрихиальных. Факультативные анаэробы; хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура 37°С. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты, но не газа, Оксидазо- и каталазоположительные. Образуют индол. Реакция Фогеса-Проскауэра отрицательная. По лизин- и орнитиндекарбоксилазе и арганиндигидролазе положительные. По липазе отрицательные. Восстанавливают нитрат. Большинство штаммов чувствительны к вибриостатическому агенту О/129. Содержат общий для эктеробактерий антиген. Встречаются у рыб и других водных животных и у разнообразных млекопитающих. Ассоциированы с диареей и редкими оппортунистическими инфекциями у человека.
Типовой (и единственный) вид: Plesiomonas shigelloides.
Признаки, отличающие Plesiomonas от других родов, приведены в табл. 5.46 и 5.47.
Признаки Plesiomonas shigelloses: См. табл. 5.50.
Таблица 5.50. биохимические реакции Plesiomonas shigelloses1

Колонии после роста в течение 24 ч при 37°С на МПА или кровяном агаре 1,0–1,5 мм в диаметре, сероватые, блестящие, матовые, с гладкой поверхностью и цельным краем. Проба с метиловым красным вариабельная. Результаты тестов на использование цитрата, дезоксирибонуклеазу, гидролиз эскулина, разжижение желатины, образование H2S, рост в присутствии KCN, малонат, О-нитрофемил-β-D-галактопиранозид, фенил аланиндезаминазу и уреазу отрицательные. Образуют кислоту из глицерола, мио-инозитола, мальтозы и трегалозы.
Род Vibrio
Прямые или изогнутые палочки, 0,5–0,8 х 1,4–2,6 мкм. Грамотрицательные. Подвижные за счет одного или более полярных жгутиков, заключенных в чехол – продолжение наружной мембраны клеточной стенки. Факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метоболизма. Оптимальная температура существенно варьирует; все виды растут при 20°С, большинство – при 30°С. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты, но не газа (исключение составляют V. furnissii, К gazogenes и некоторые штаммы V. (Listonella) damsela). Оксидазоположительные (за исключением V. gazogenes и V. metschnikovii). Восстанавливают нитрат (за исключением V. gazogenes, V. metschnikovii и V. ordalii). Углеводы, сбраживаемые большинством видов, включают мальтозу, D-маннозу и трегалозу. Большинство видов чувствительны к вибриостатическому агенту О/129, Ионы натрия стимулируют рост всех видов, а для большинства видов абсолютно необходимы. Обнаружены в водных местообитаниях, существенно варьирующих по степени солености. Широко распространены в морской среде и в устьях рек, на поверхности морских животных и в содержимом их кишечника. Некоторые виды обнаружены также в пресной воде. Около десяти видов патогенны для человека, несколько видов патогенны для морских позвоночных и беспозвоночных. Наиболее известные возбудители заболеваний человека – V. cholerae, возбудитель холеры, V. parahaemolyticus, основной возбудитель пищевых отравлений, вызванных зараженной рыбой или моллюсками, и V. vulnificus, вызывающий септицемию с высоким процентом летальности. Эти и другие виды ассоциированы с раневыми инфекциями, желудочно-кишечными заболеваниями и разнообразными внекишечными инфекциями.
Типовой вид: Vibrio cholerae.
Признаки, отличающие Vibrio от других родов, приведены в табл. 5.46. Хотя число легкоопределяемых диагностических различий между этими родами ограничено, их дифференциация не представляет существенных трудностей благодаря четкому определению (как правило) отдельных видов.
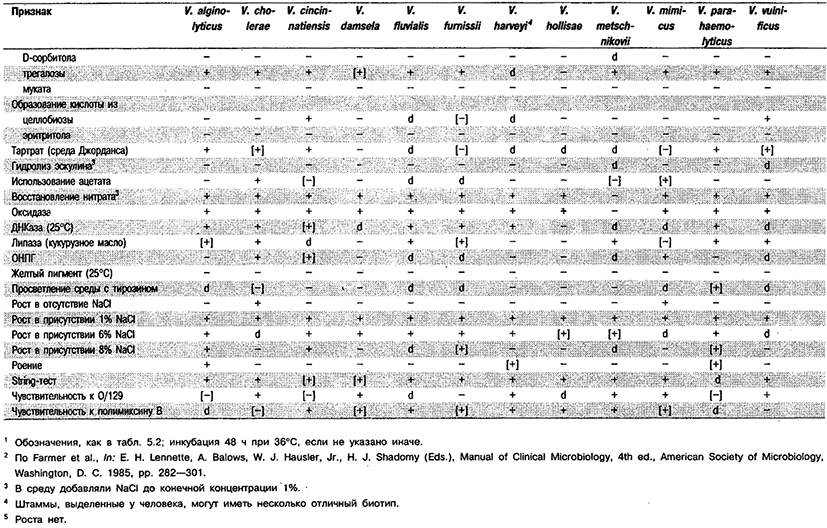
Дифференциация видов рода Vibrio. Признаки, разграничивающие биовары Vibrio cholerae, приведены в табл. 5.51. Признаки, помогающие дифференцировать виды Vibrio, особенно морские вибрионы, представлены в табл. 5.52А. Признаки, устанавливаемые с помощью тестов, которые лучше проводить в клинических лабораториях, перечислены в табл. 5.52Б для тех видов, которые рекомендуется анализировать также в клинических лабораториях.
Таблица 5.51. Дифференциация биоваров вида V. cholerae1 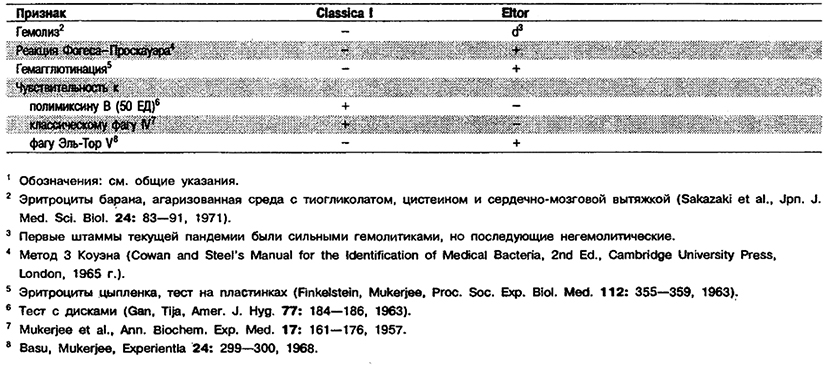
Таблица 5.52 А. Дифференциация видов и биоваров рода Vibrio1
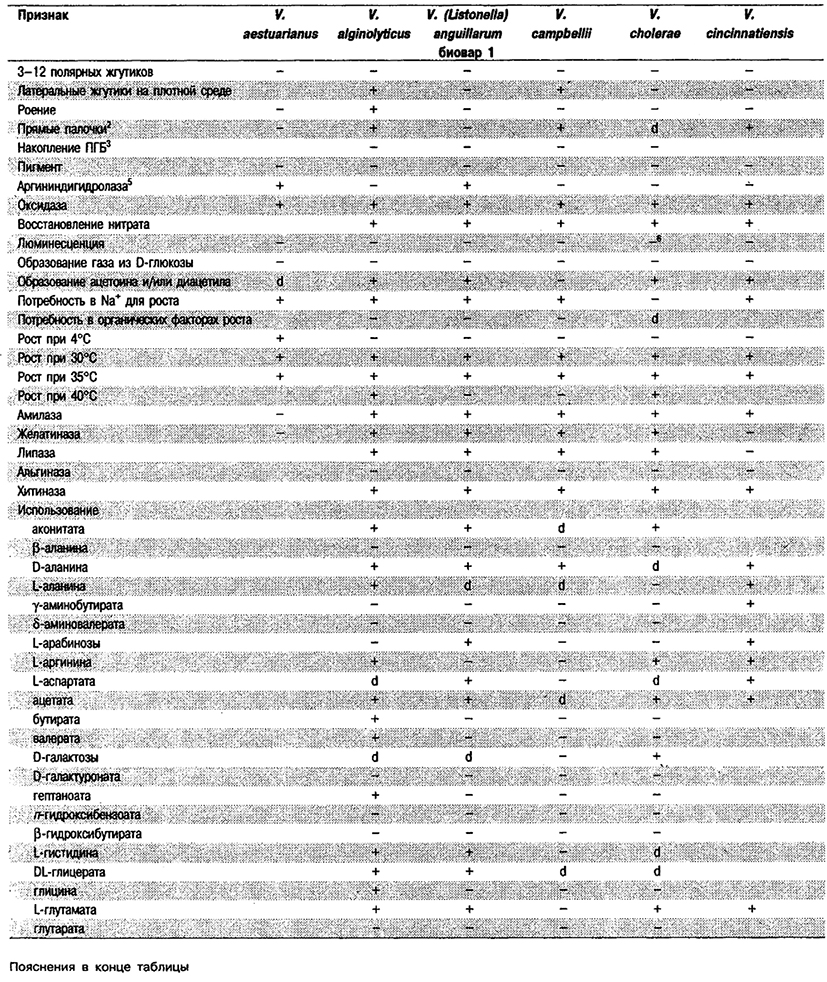
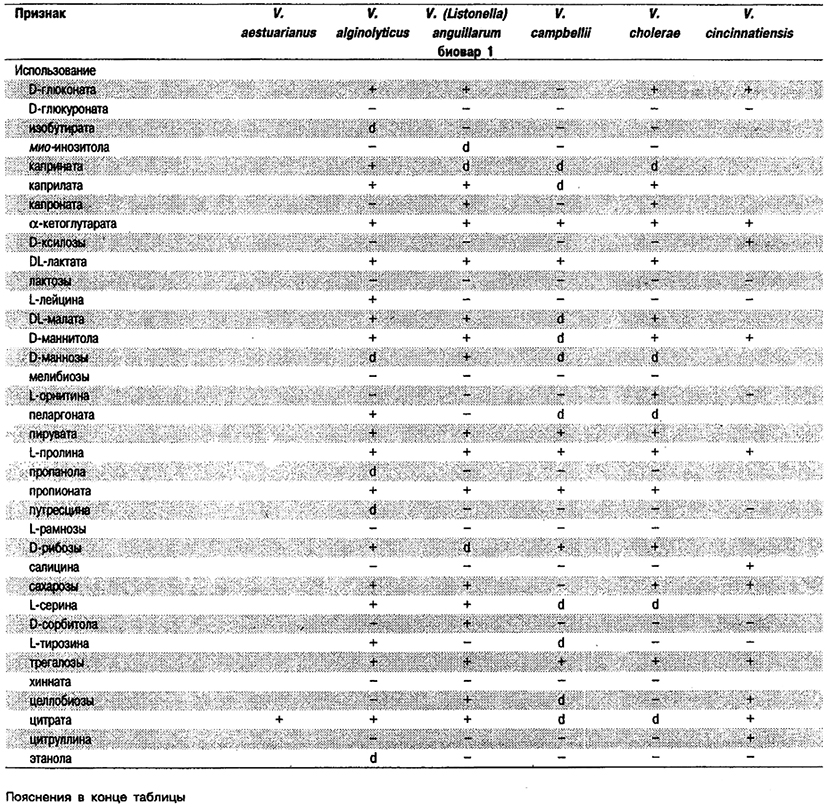
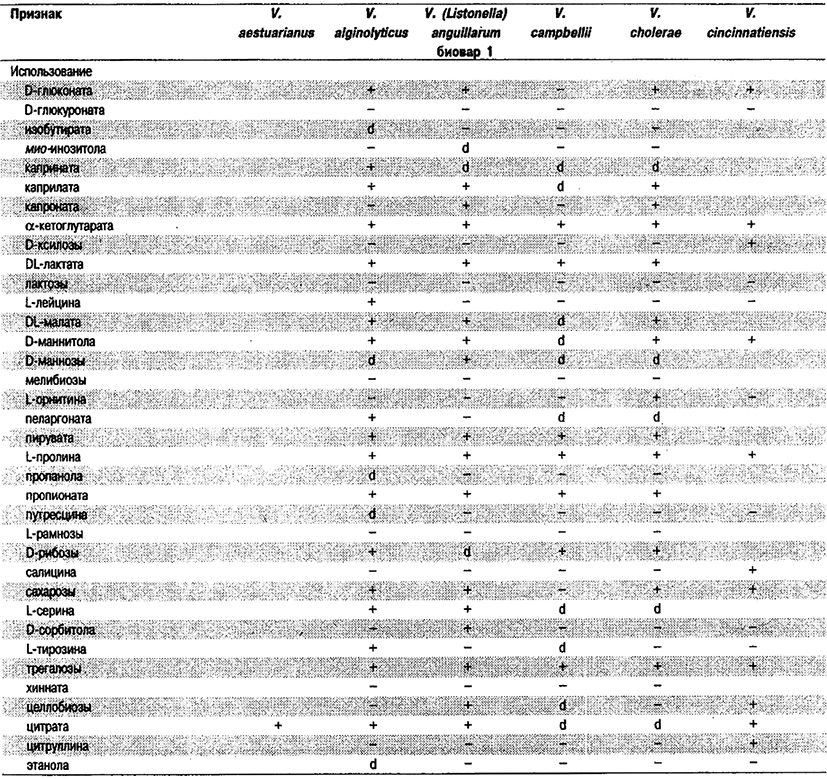
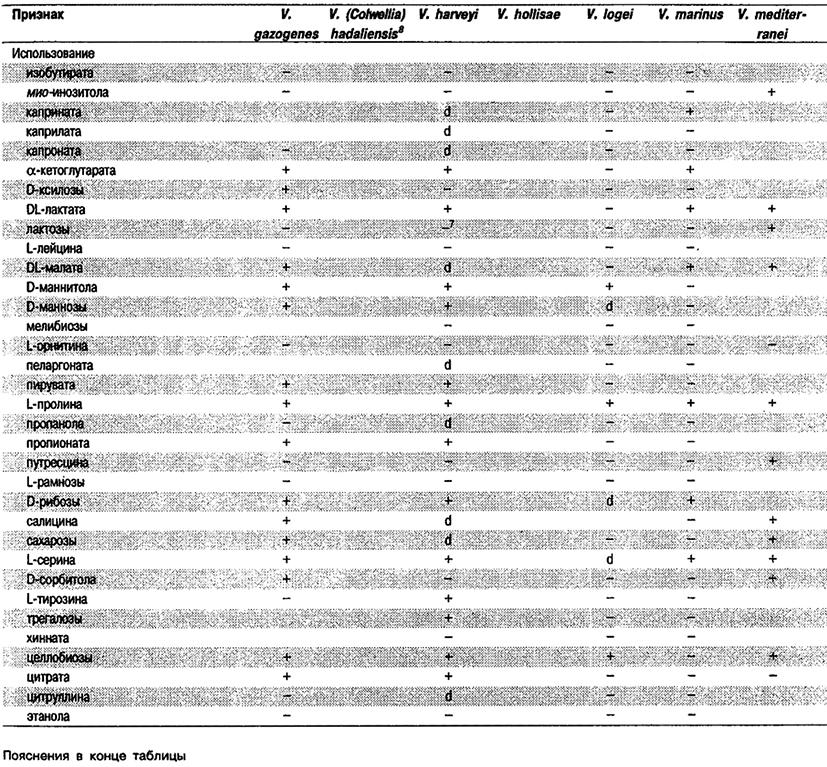
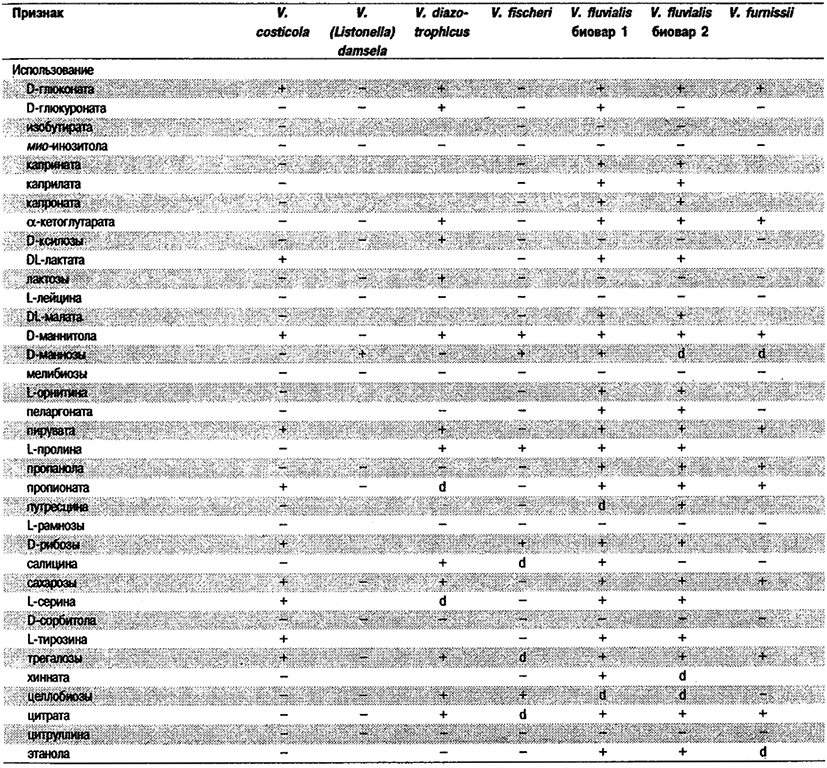
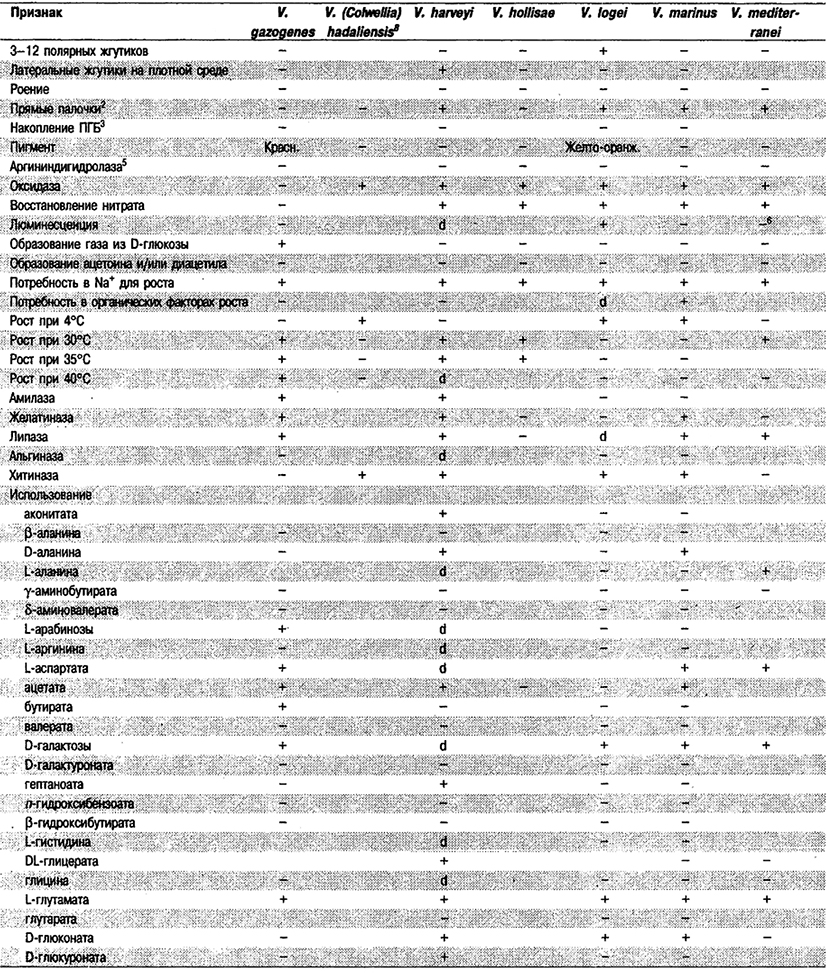
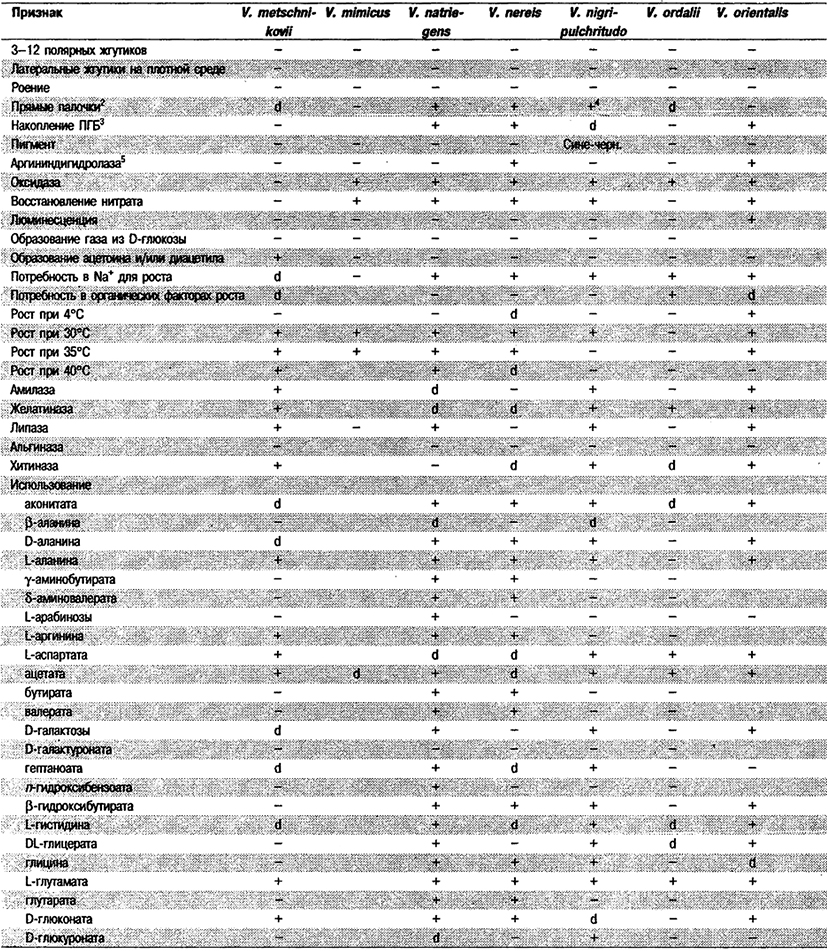
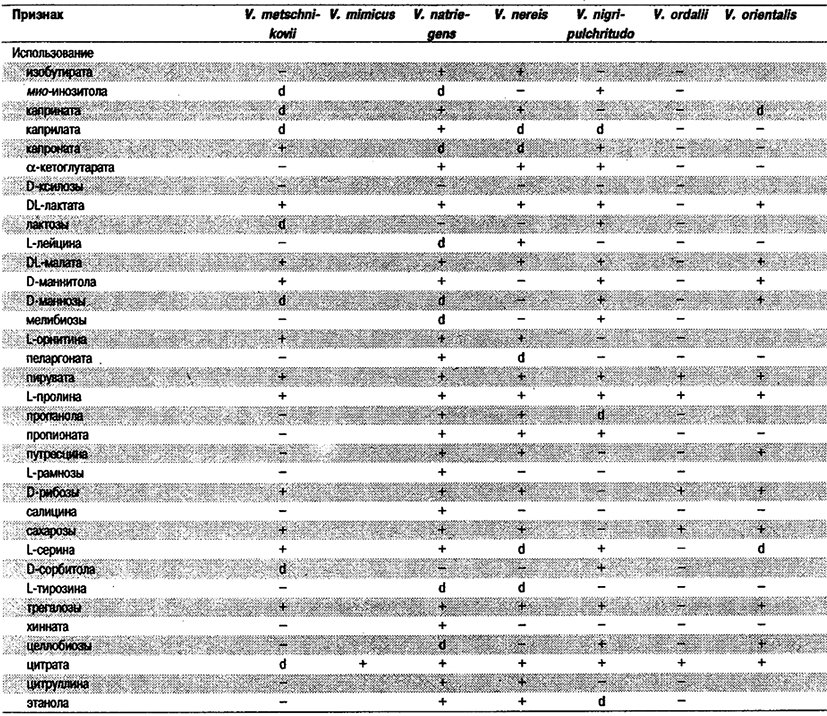
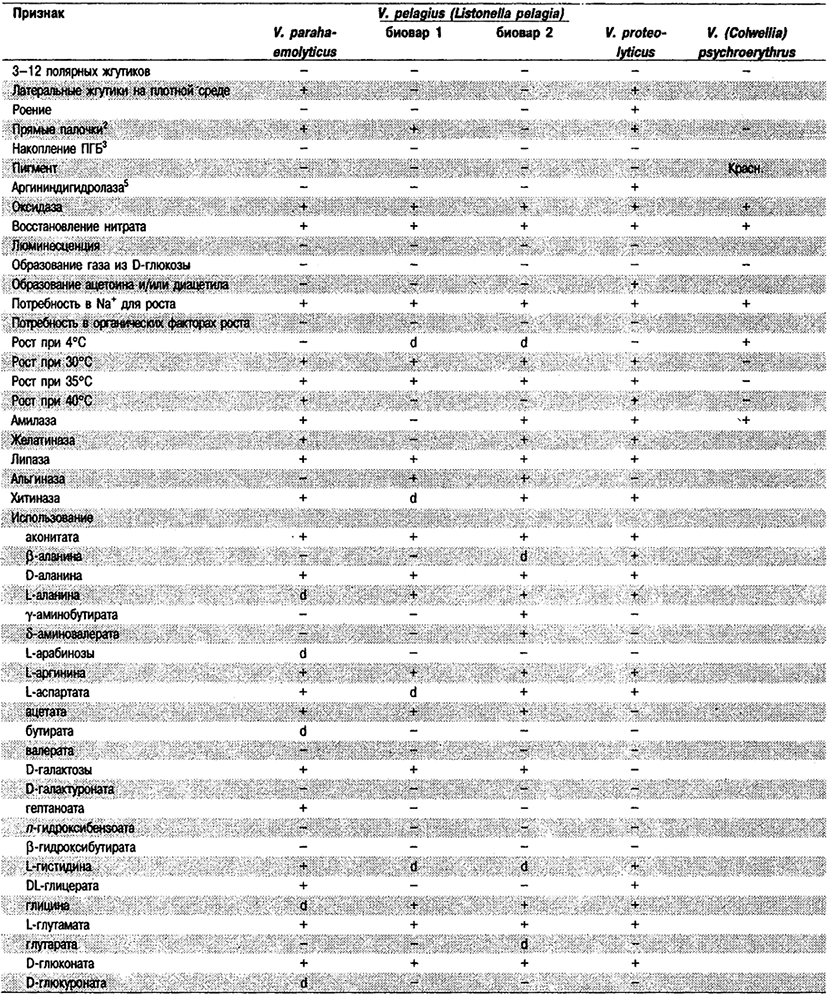
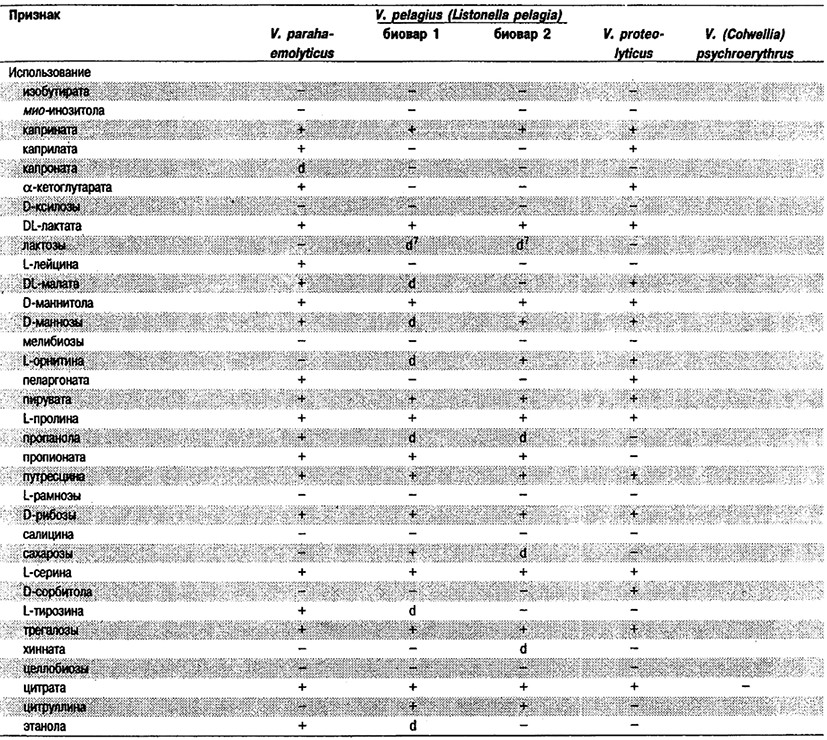
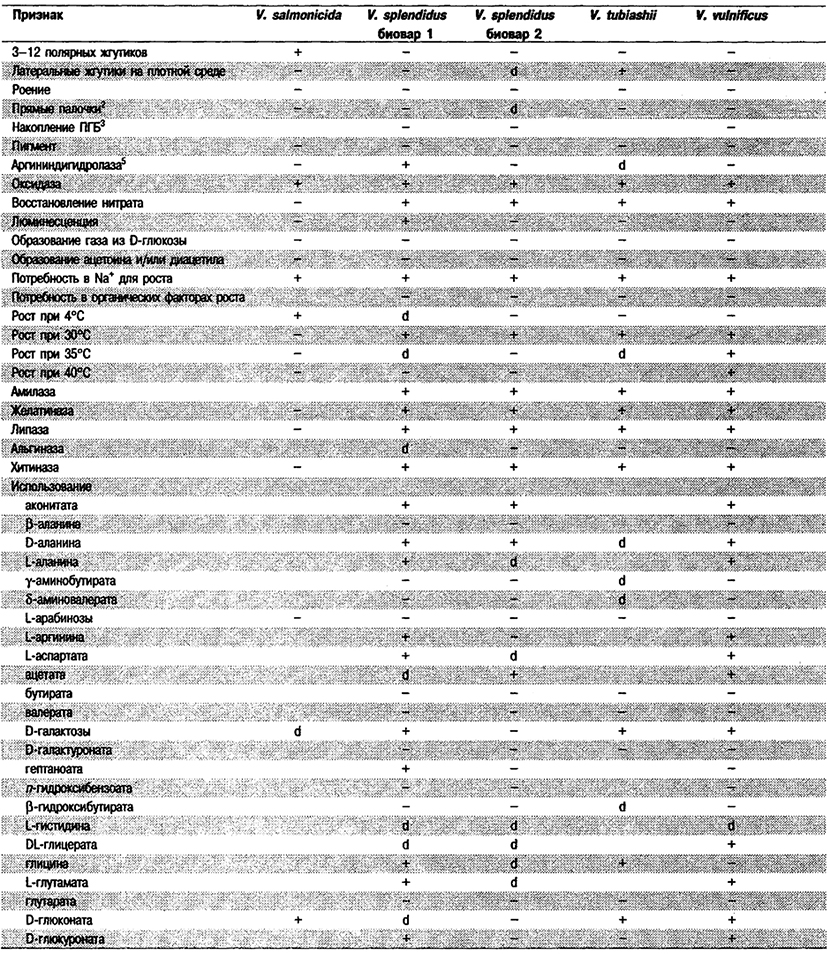
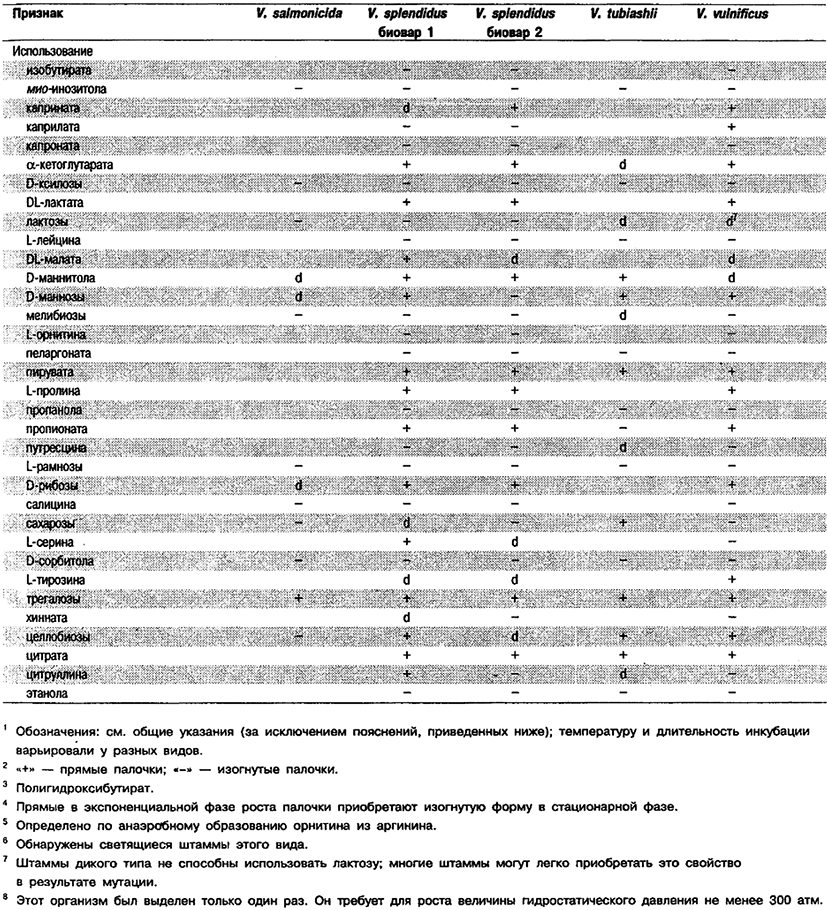
Таблица 5.52 Б. Дифференциация видов рода Vibrio, обнаруженных в клиническом материале 1,2 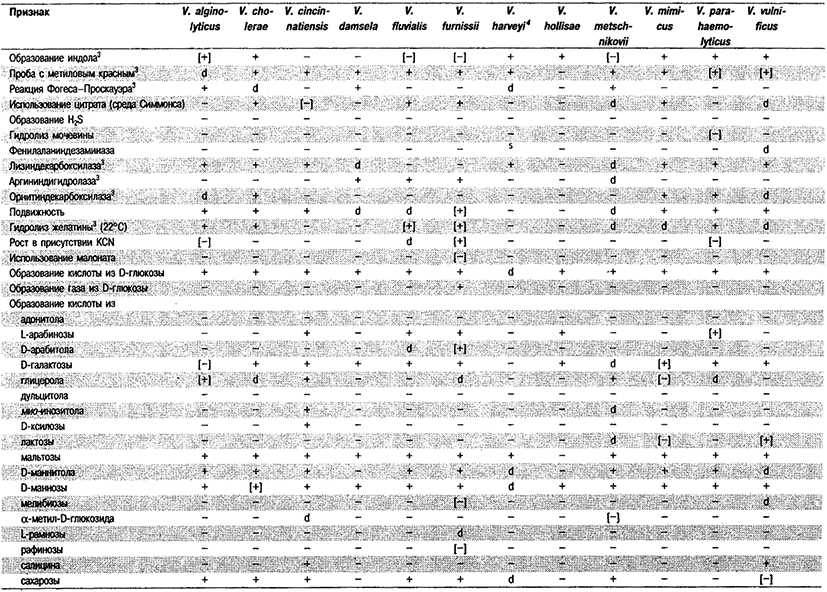
На основе полного комплекса данных по всем представителям Vibrionaceae создана программа для ПЭВМ и отдельно программа для морских вибрионов (тесты проводились при 25°С и наличии катионов из состава морской воды). Для получения информации можно обращаться письменно по адресу: J.J. Farmer III, Centers for Disease Control, Building l, Room B310, Mailstop C03, Atlanta, Georgia 30333, USA.
От редакции; Род Listonella описан в 1985 г, (MacDonell, Colwell, Syst. Appl. Microbiol. 6: 171–182; Int. J. Syst. Bacteriol. 36: 354–356, 1986) для видов Listonella (Vibrio) anguillarum, Listonella (Vibrio) damsel а и Listonella (Vibrio pelagius) pelagia, ранее относившихся к роду Vibrio. Род Colwellia описан в 1988 г. (Deming et al, Syst. Appl. Microbiol. 10: 152–160; Int. J. Syst. Bacteriol 38: 328–329, 1988) для видов Colwellia (Vibrio) psychroerythrus и Colwellia (Vibrio) hadaliensis. Вид С. psychroerythrus был включен в род Vibrio, хотя название никогда не было узаконено. Поскольку очевидно, что род Vibrio требует детальных исследований в связи с его эволюционной гетерогенностью, для целей данного Определителя, не предрешая выводы, решено оставить все эти виды в роде Vibrio. Родовые названия Listonella и Colwellia приведены в скобках во избежание путаницы.
Виды Vibrio aestuarianus, Vibrio (Listonella) damsela, Vibrio diazotrophicus, Vibrio hollisae, Vibrio mimicus, Vibrio ordalii и Vibrio orientalis были указаны, но не описаны под рубрикой «Другие организмы» в разд. «Род Vibrio» в «Bergey's Manual of Systematic Bacteriology». Виды Vibrio cincinnatiensis, Vibrio furnissii, Colwellia (Vibrio) hadaliensis, Vibrio mediterranei, Colwellia (Vibrio) psychroerythrus,
Vibrio salmonicida и Vibrio tubiashil не приведены в «Beigey's Manual of Systematic Bacteriology». Вид Vibrio aestuarianus описан в 1983 г. (Tison, Seidler, Int. J. Syst. Bacteriol. 33: 699–702), Вид Vibrio cincinnatiensis описан в 1986 г. (Brayton et al., J. Clin. Microbiol. 23: 104–108; Int. J. Syst. Bacteriol. 36: 354–356, 1986). Вид Vibrio (Listonella) damsela описан в 1981 г. (Love et al., Science 214: 1139– 1140; Int. J. Syst. Bacteriol. 32: 266-268, 1982). Вид Vibrio diazotrophicus описан в 1982 г. (Guerinot et al., Int. J. Syst. Bacteriol. 32: 350–357). Вид Vibrio furnissii оиисан в 1983 г. (Brenner et al., J. Clin. Microbiol. 18: 816–824; Int. J. Syst. Bacteriol. 34: 91–92, 1984). Вид Colwellia (Vibrio) hadaliensis описан в 1988 г. (Deming et al., Syst. Appl. Microbiol. 10: 152–160; Int. J. Syst. Bacteriol. 38: 328-329,). Bud Vibrio hollisae описан в 1982 г. (Hickman et al., J. Clin. Microbiol. 15: 395–401); Int J. Syst. Bacteriol. 32: 384–385, 1982). Вид Vibrio mediterranei описан в 1986 г. (Pujalte, Garay, Int. J. Syst. Bacteriol. 36: 271–281). Вид Vibrio mimicus описан в 1981 г. (Davis et al., J. Clin. Microbiol. 14: 631–639; Int. J. Syst. Bacteriol. 32: 266–26 8, 1982). Вид Vibrio ordalii описан в 1981 г, (Schiewe et al., Curr. Microbiol. 6: 343–348; Int. J, Syst. Bacteriol. 32: 384–385, 1982). Вид Vibrio orientalis описан в 1983 г. (Yang et al., Curr. Microbiol, 8: 95–100; Int. J. Syst. Bacteriol. 33: 672– 674, 1983). Вид «Vibrio phychroerythrus» описан в 1972 г. (D'Aoust, Kushner, J. Bacteriol. Ill: 340– 342). Последний вид был заново описан в 1988 г. как Colwellia (Vibrio) psychroerythrus (Deming et al. Syst. Appl, Microbiol. 10: 152–160; Int. J. Syst. Bacteriol. 38: 328–329, 1988). Вид Vibrio salmonicida описан в 1986 г. (Wiik, Egidium, Int. J. Syst. Bacteriol, 36: 521–523), Вид Vibrio tubiashi описан в 1984 г. (Hada et al., Int. J. Syst. Bacteriol. 34: 104),
Вид Vibrio carchariae, описанный в 1984 г. (Grimes et al., Microb. Ecol. 10: 271–282; Int. J. Syst. Bacteriol. 35: 223–225, 1985), согласно данным гибридизации ДНК – младший субъективный синоним названия Vibrio harveyi (F. W. Hickman–Brenner, J.J. Farmer III, D.J. Brenner, G. R, Fanning, неопубликованные данные).
