Эта группа грамотрицательных бактерий включает традиционные семейства Enterobacteriaceae, Vibrionaceae и Pasteurettaceae, а также несколько других родов факультативно анаэробных палочек. Диаметр клеток составляет 0,1–1,5 мкм; среди представителей многих родов встречаются удлиненные нитевидные формы. Бактерии имеют вид прямых палочек, кроме представителей рода Vibrioдля которых характерна изогнутая или вибриовдная форма. Организмы данной группы могут встречаться как свободноживущие или в ассоциациях с хозяевами – животными или растениями (Erwinia). Многие виды патогенны для человека и животных, а некоторые – для насекомых и растений.
Дифференциация родов и семейств Группы 5: См. табл. 5.1.
Таблица 5.1. Дифференцирующие признаки родов и семейств, входящих в Группу 5.11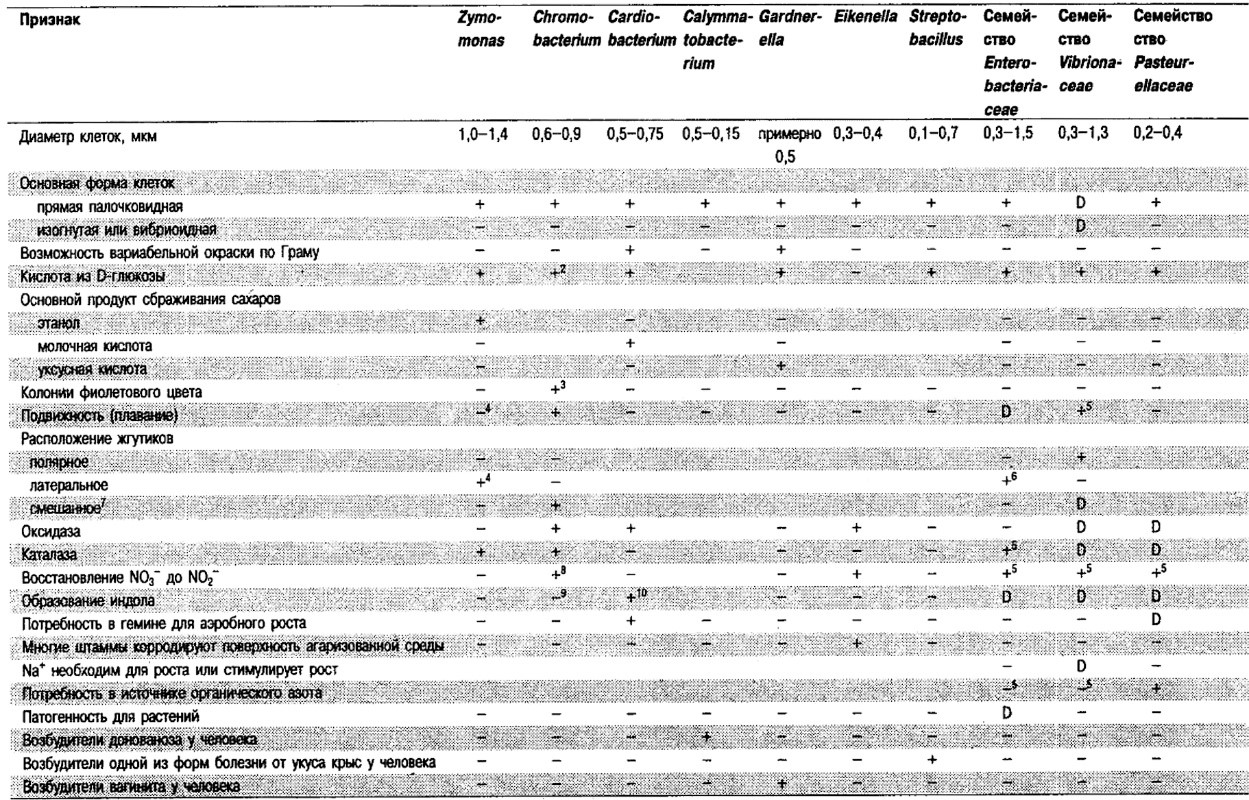
Подгруппа 1: Семейство Enterobacteriaceae
Прямые палочки, обычно 0,3–1,8 мкм. По Граму окрашиваются отрицательно. Подвижные за счет перитрихиальных жгутиков, за исключением Tatumella, или неподвижные. Факультативные анаэробы; хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Большинство видов хорошо растет при 37°С, однако некоторые виды лучше растут при 25–30°С и при этой температуре метаболически более активны. D-глюкозу и другие углеводы катаболизируют с образованием кислоты, а многие виды, кроме того, и газа. Оксидазоотрицательные и каталазоположительные, за исключением Shigella dysenteriae О Группа 1 и видов Xenorhabdus, но не X. luminescens. За исключением Arsenophonus, ряда видов Erwinia, большинства видов Xenorhabdus и некоторых штаммов Klebsiella pneumoniae subsp. ozaenae, Pantoea и Yersinia, все представители восстанавливают нитрат. Распространены повсеместно. Присутствуют в почве, воде, на фруктах, овощах, зерне, цветковых растениях и деревьях, у животных (от червей и насекомых до млекопитающих) и человека. Входящие в эту подгруппу микроорганизмы весьма разнообразны по особенностям экологии, кругу хозяев, а также патогенности для человека, животных, насекомых и растений. Ряд видов вызывает желудочно-кишечные заболевания, включая тиф и бактериальную дизентерию. Многие виды, обычно не связанные с желудочно-кишечными заболеваниями, часто могут служить возбудителями оппортунистических инфекций. Большинство этих видов, как и виды, вызывающие желудочно-кишечные заболевания, могут быть возбудителями разнообразных внекишечных инфекций, таких как бактериемия, менингит, а также инфекции мочевыводящих путей, дыхательных путей и раневые инфекции. Бактерии семейства Enterobacteriaceae служат причиной 50% случаев внутрибольничных инфекций; наиболее часто их вызывают Escherichia coli, представители Klebsiella, Enterobacter, Proteus, Providencia и Serratta marcescens.
Типовой род: Escherichia.
Отличия Enterobacteriaceae от других семейств и родов Группы 5: Признаки, отличающие Enterobacteriaceae от других семейств и родов, приведены в табл. 5.1. Организмы, составляющие ррды семейства Vibrionaceae, оксидазоположительные и при росте в жидких средах имеют, полярные жгутики – эти признаки отличают их от представителей Enterobacteriaceae. Однако по крайней мере два вида Vibrio (V. metschnikovii и V. gazogenes) оксидазоотрицательные; некоторые штаммы других видов также могут быть оксидазоотрицательными или слабо положительными.
Дифференциация родов семейства Enterobacterial сеае: Дать определения родов и составить одну исчерпывающую таблицу биохимических признаков, позволяющую дифференцировать 30 родов семейства Enterobacteriaceae, практически невозможно. Диагностические и другие лаборатории идут по пути идентификации представителей этого семейства непосредственно на уровне видов при помощи одного и того же набора тестов для всех имеющих названия видов и подвидов, число которых в сумме превышает 115. Если тот или иной организм не удается идентифицировать точно, используют имеющиеся дополнительные тесты для дифференциации подвидов, видов внутри данного рода или родов. Биохимические признаки видов семейства Enterobacteriaceae приведены в табл. 5.2.
Таблица 5.2. Биохимическая дифференциация видов семейства Впterobacteriacеае1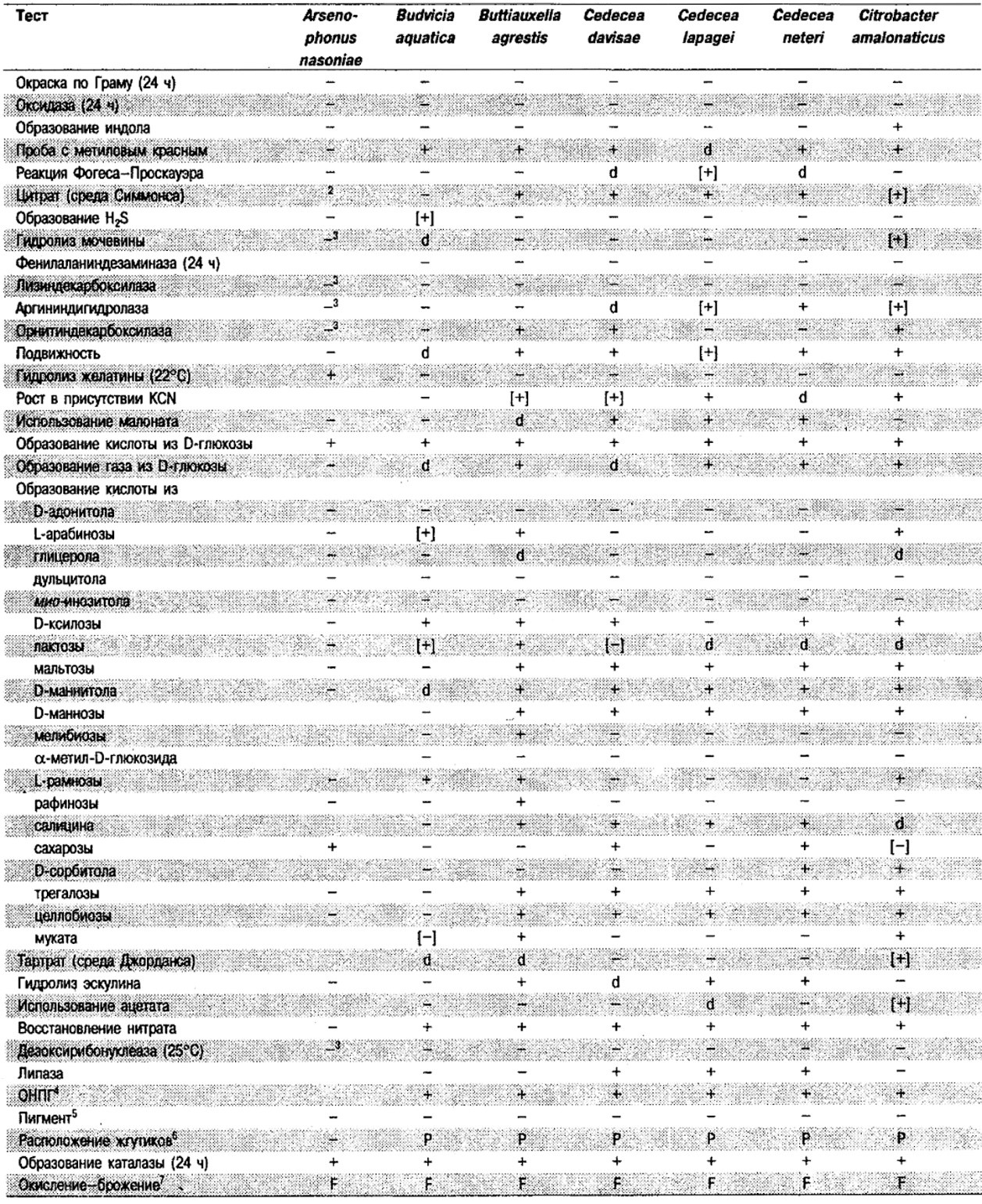
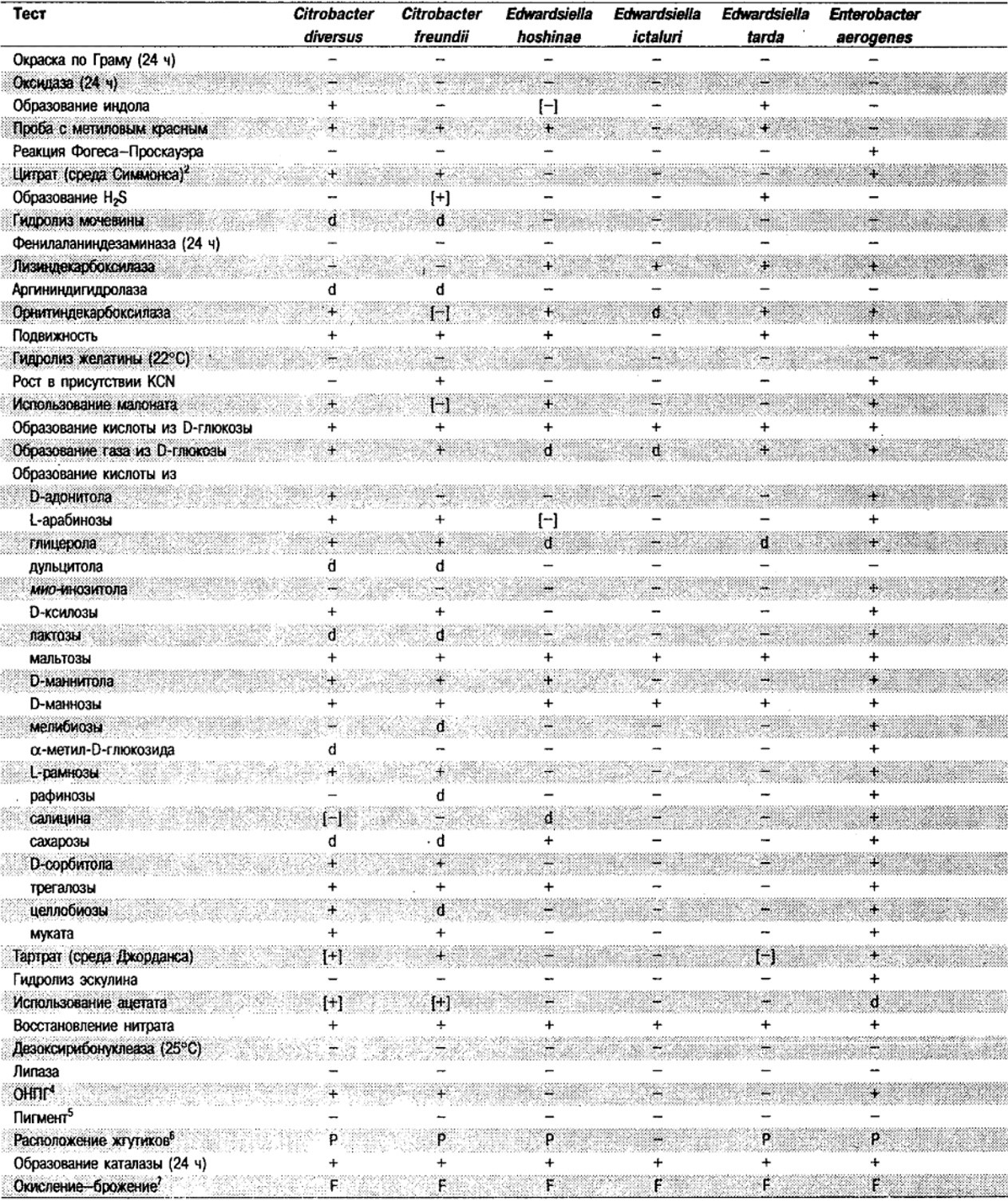
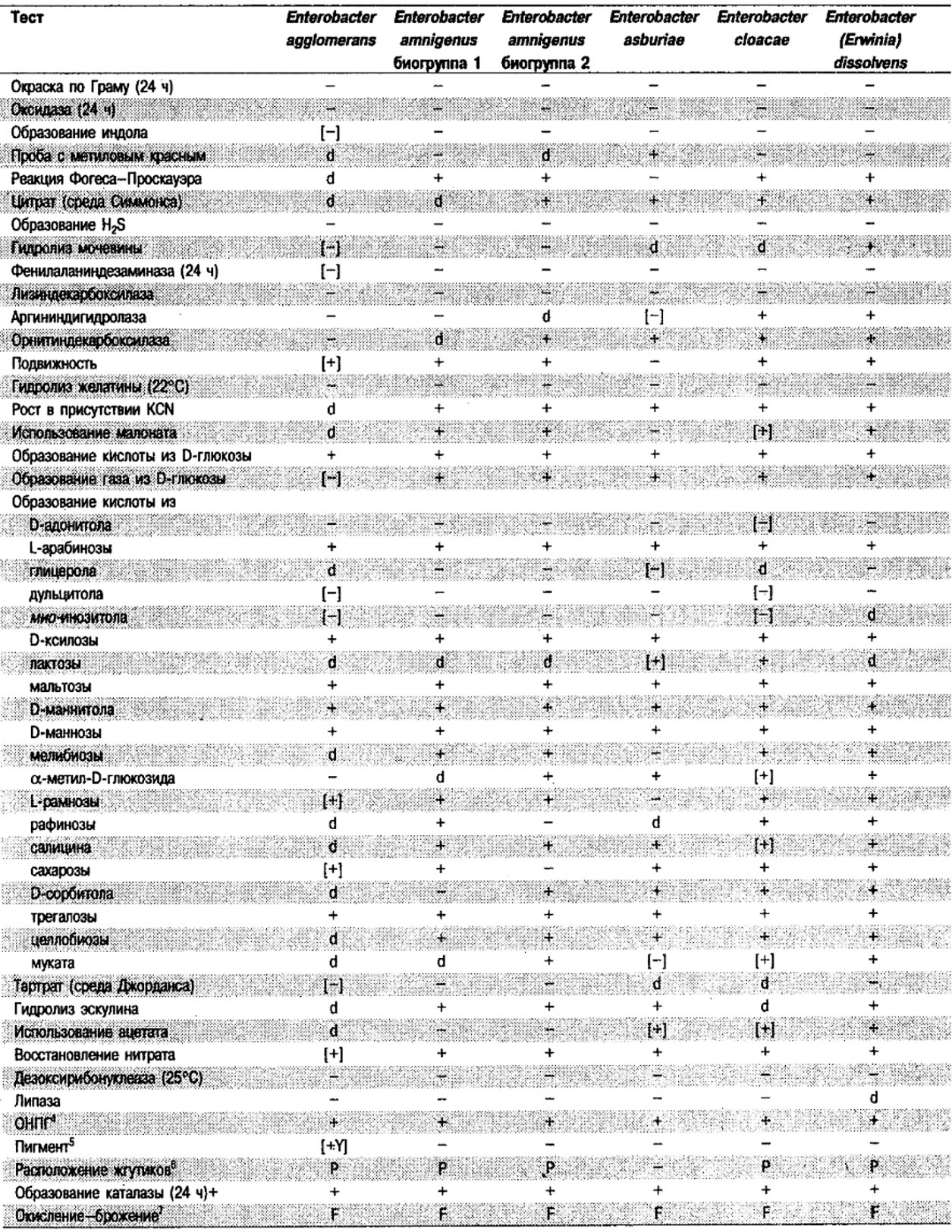
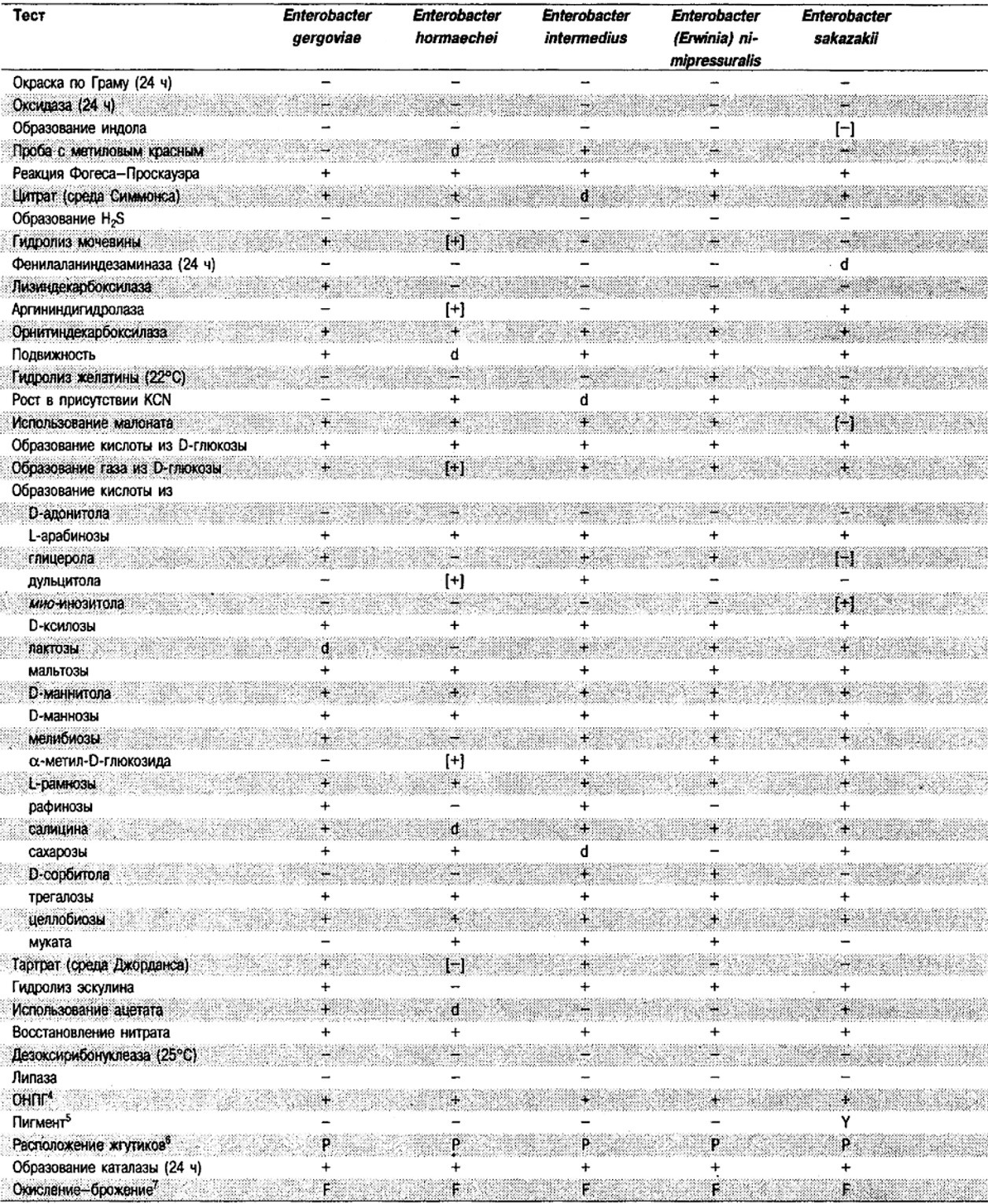
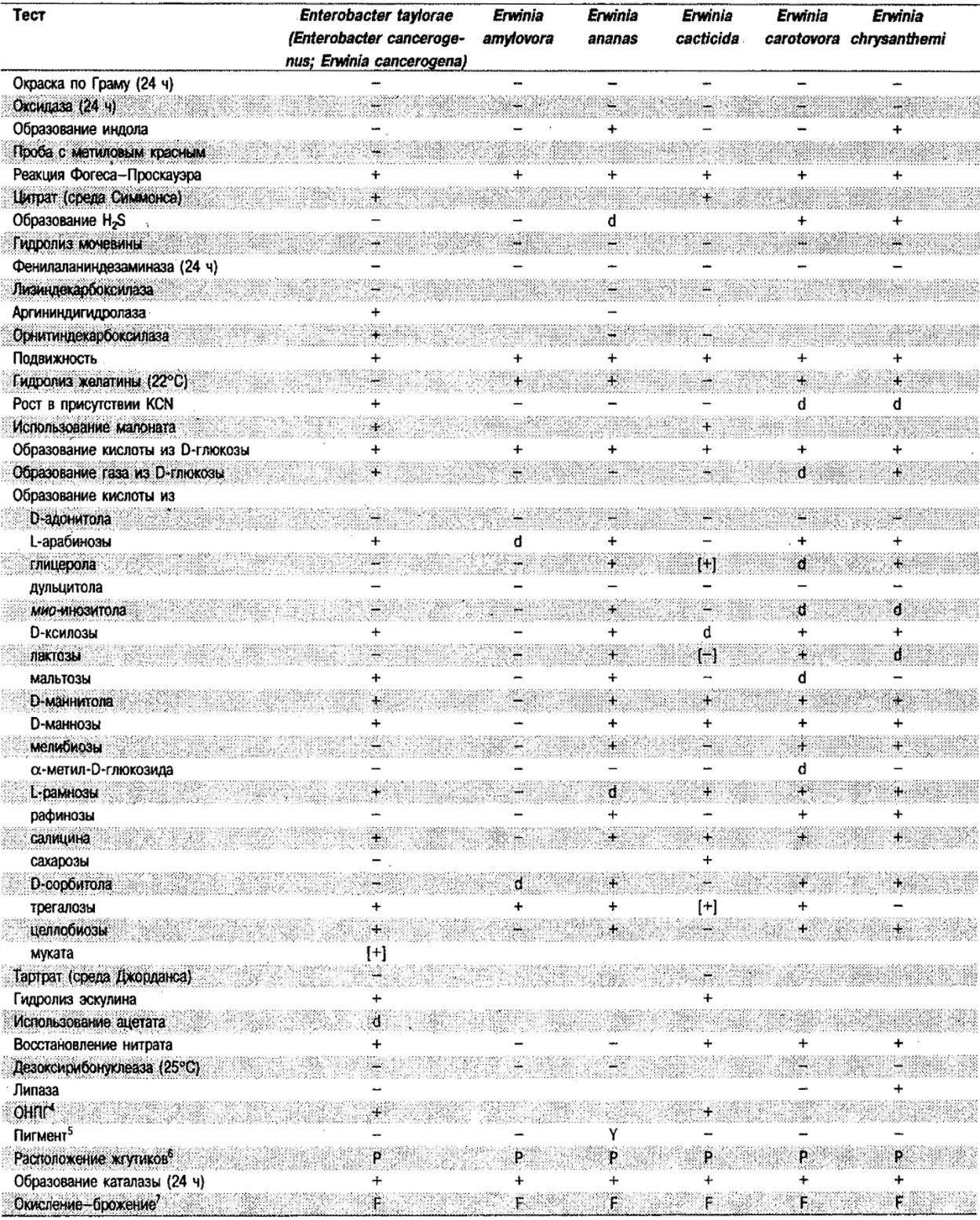
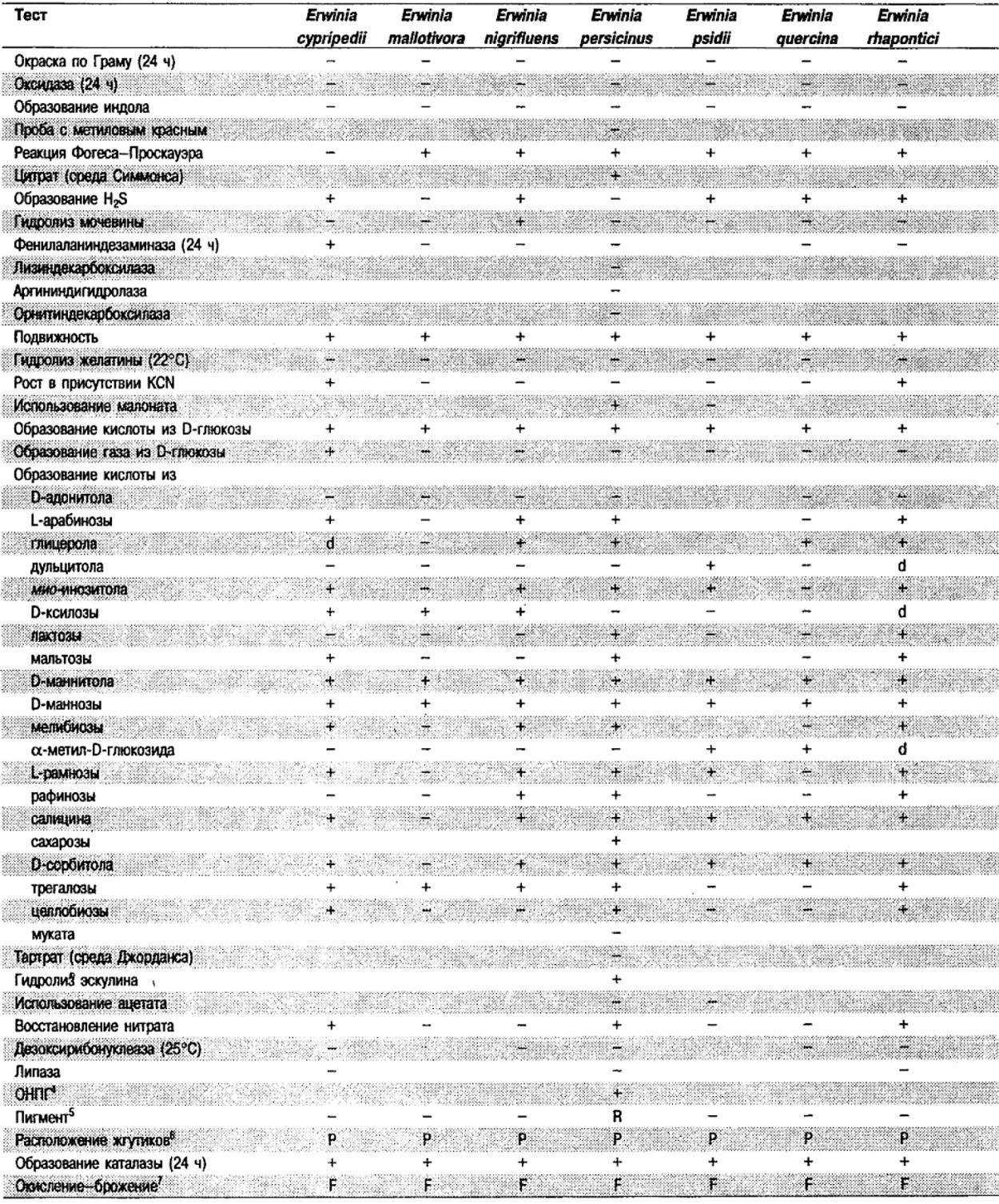
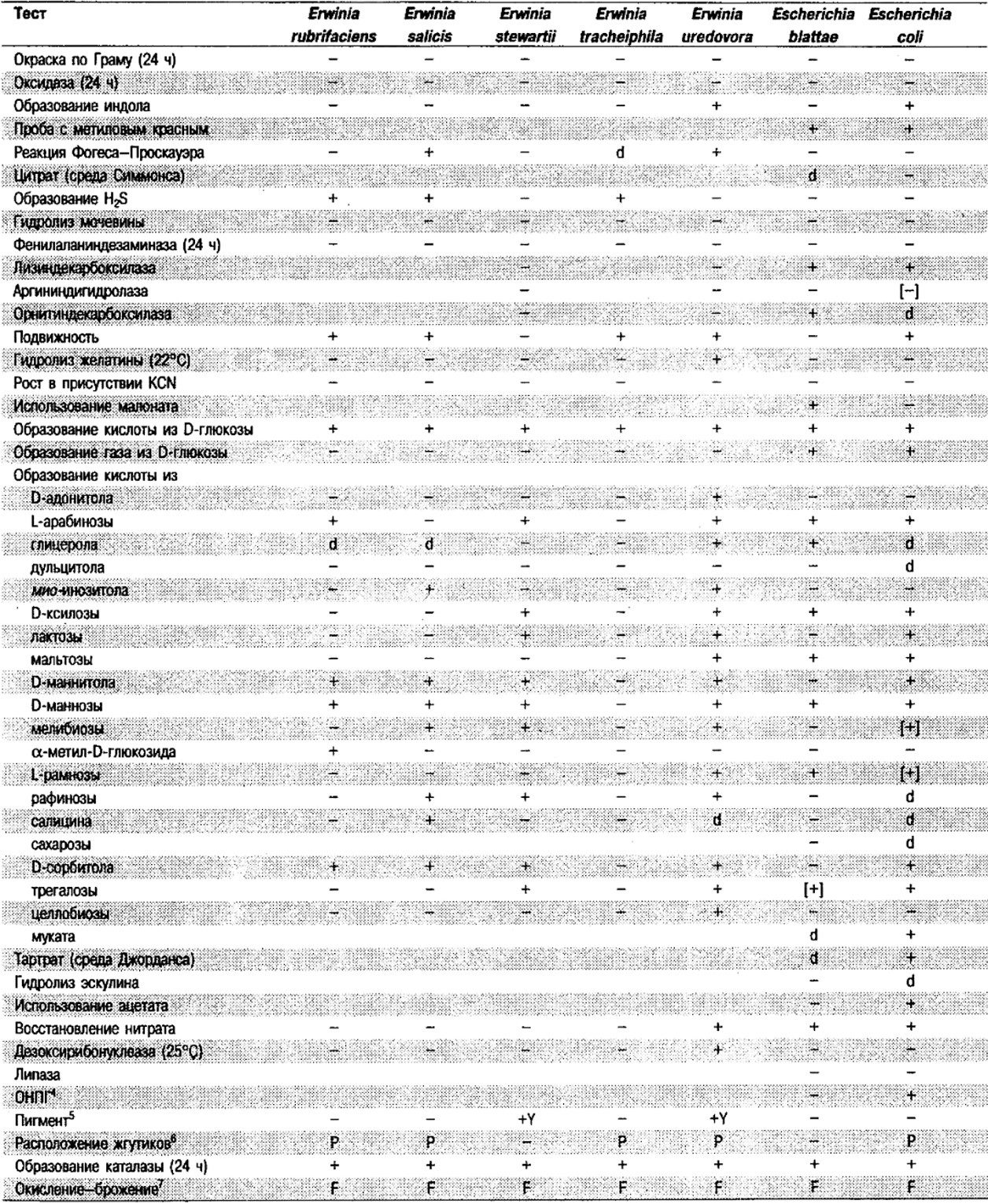
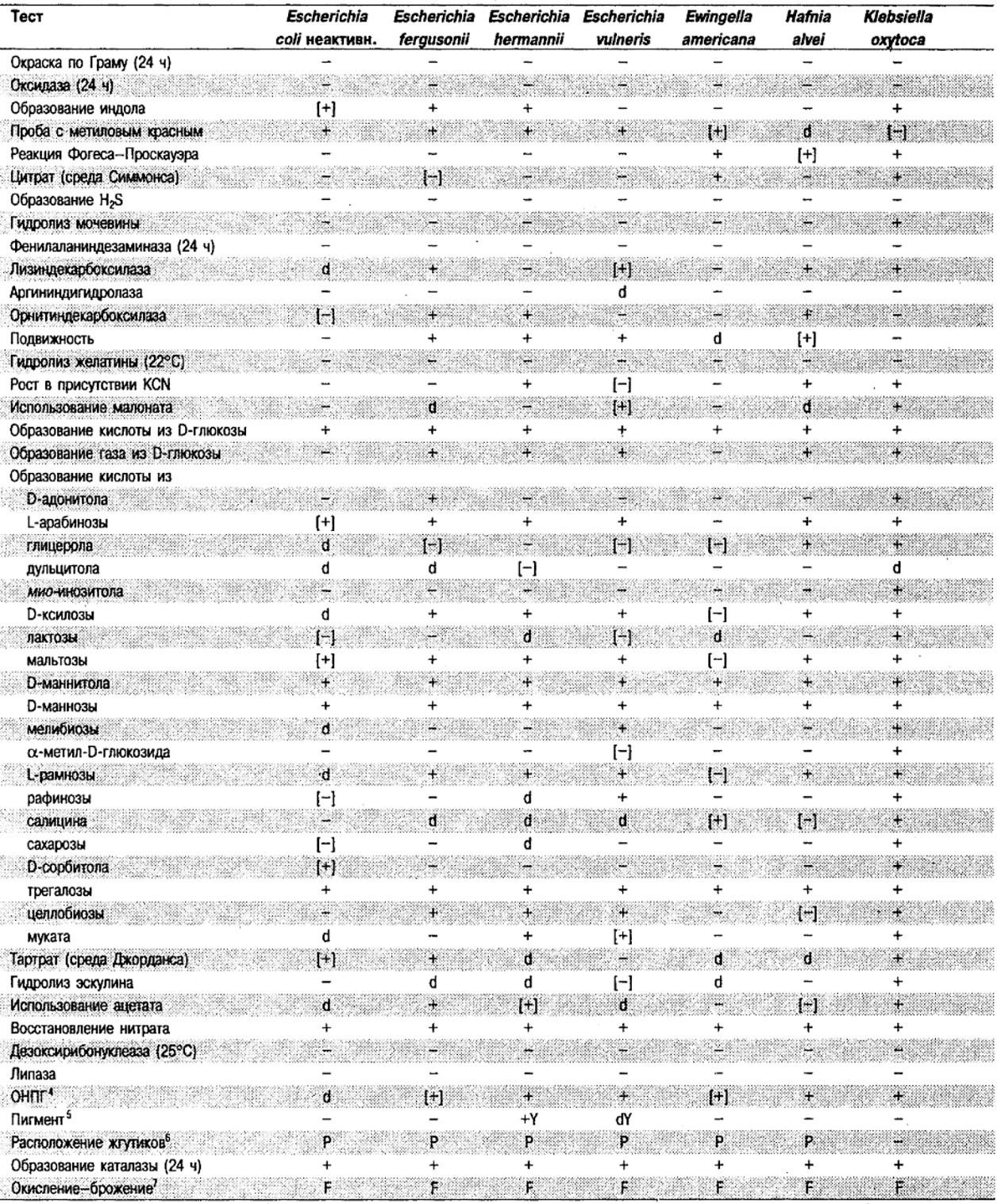
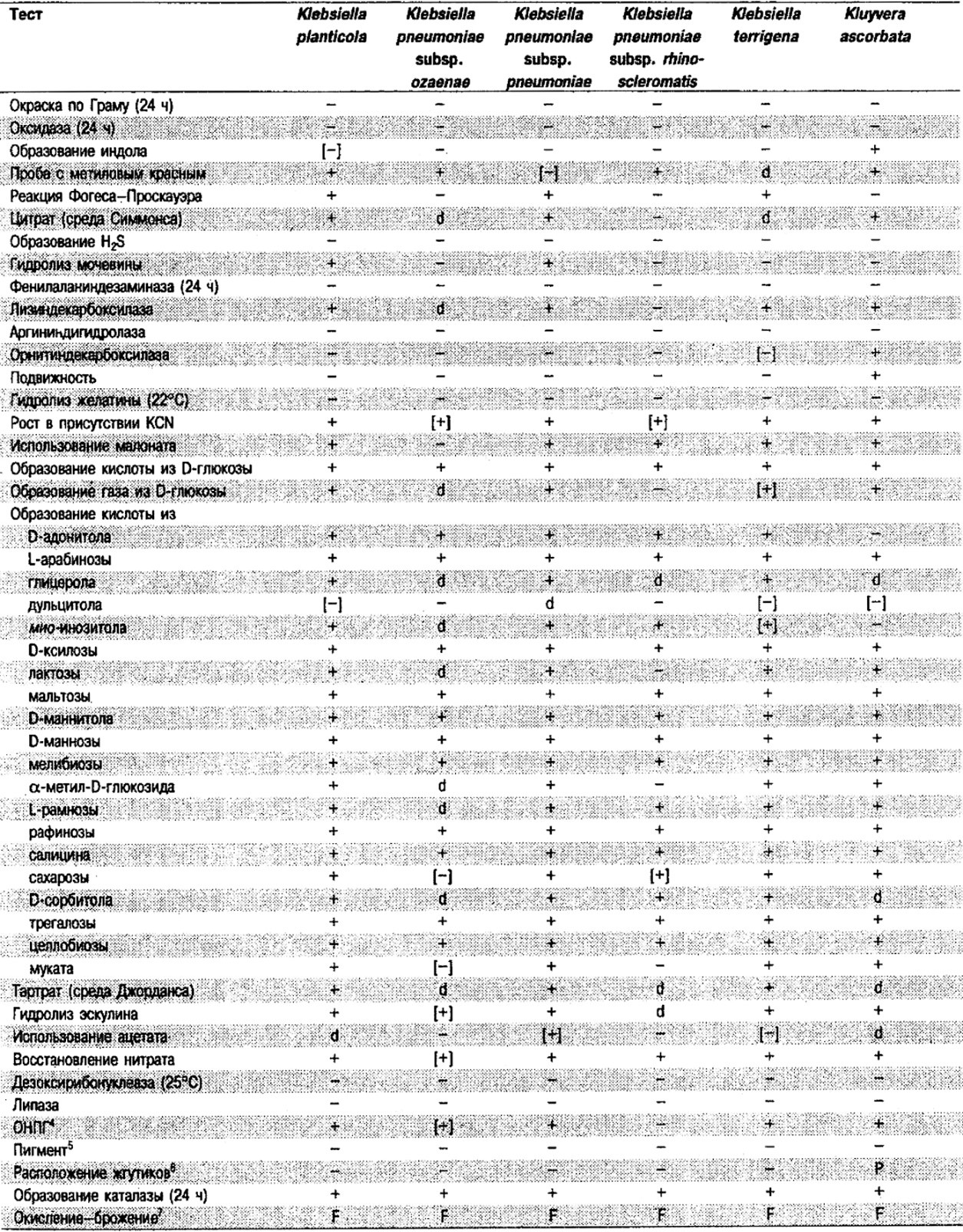
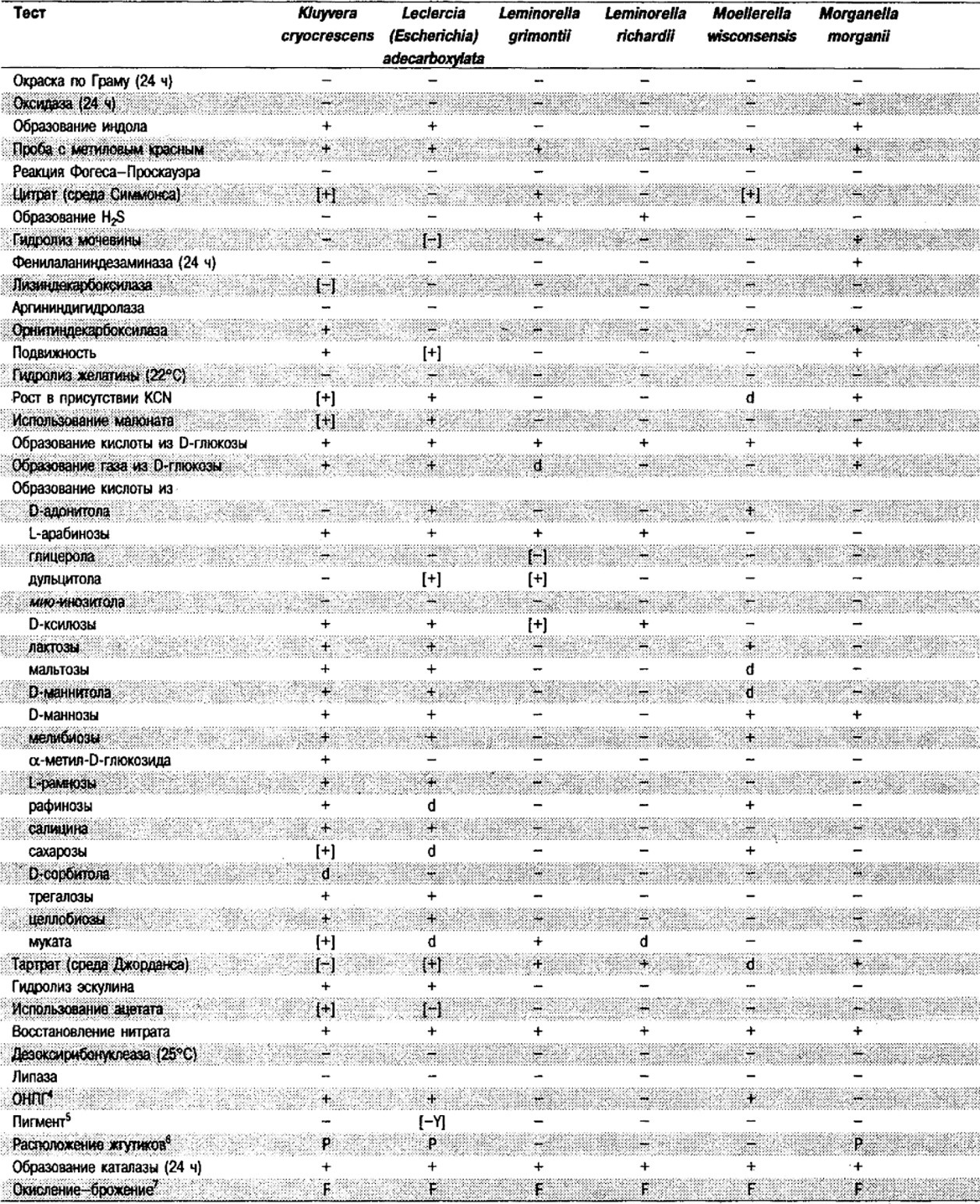
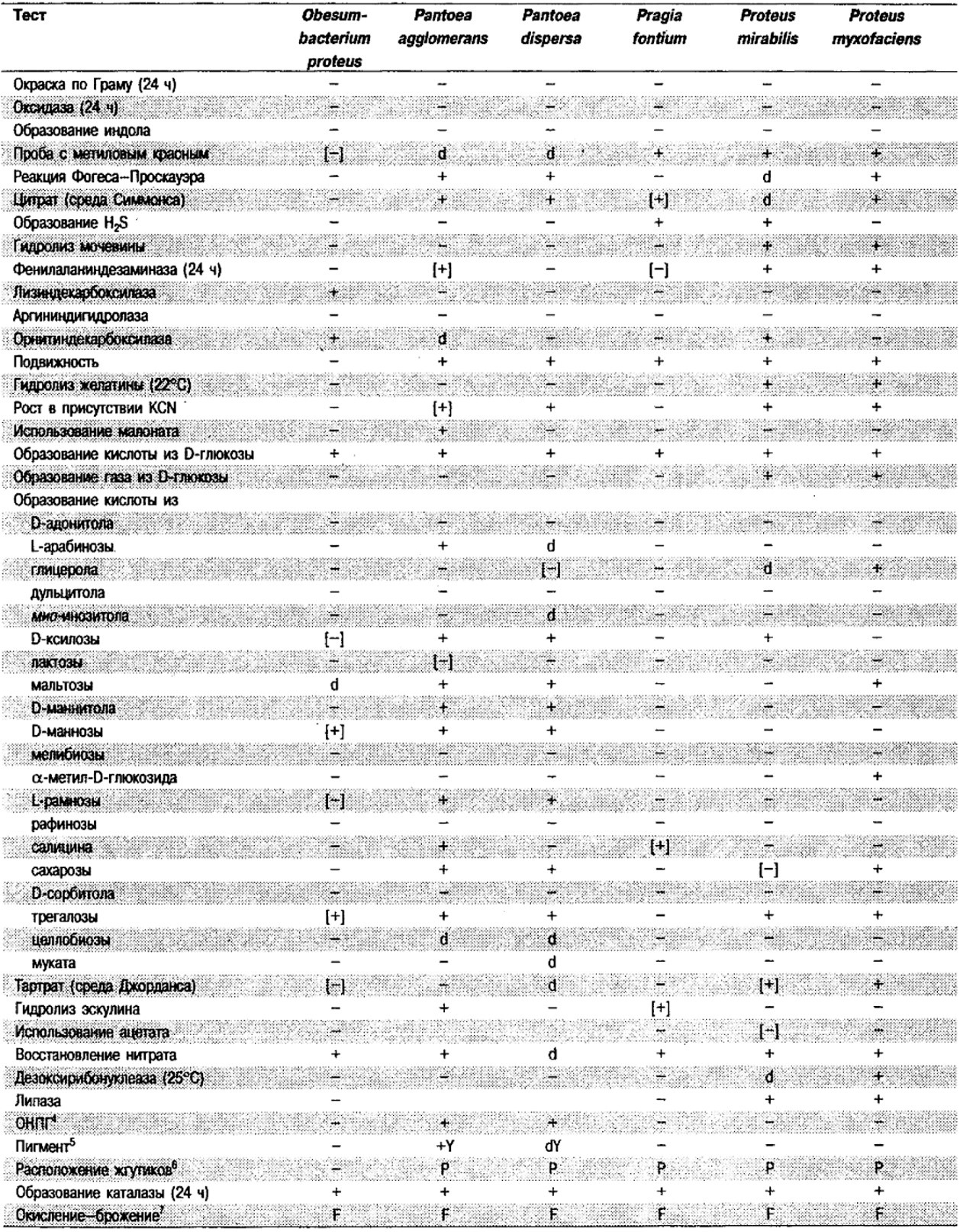
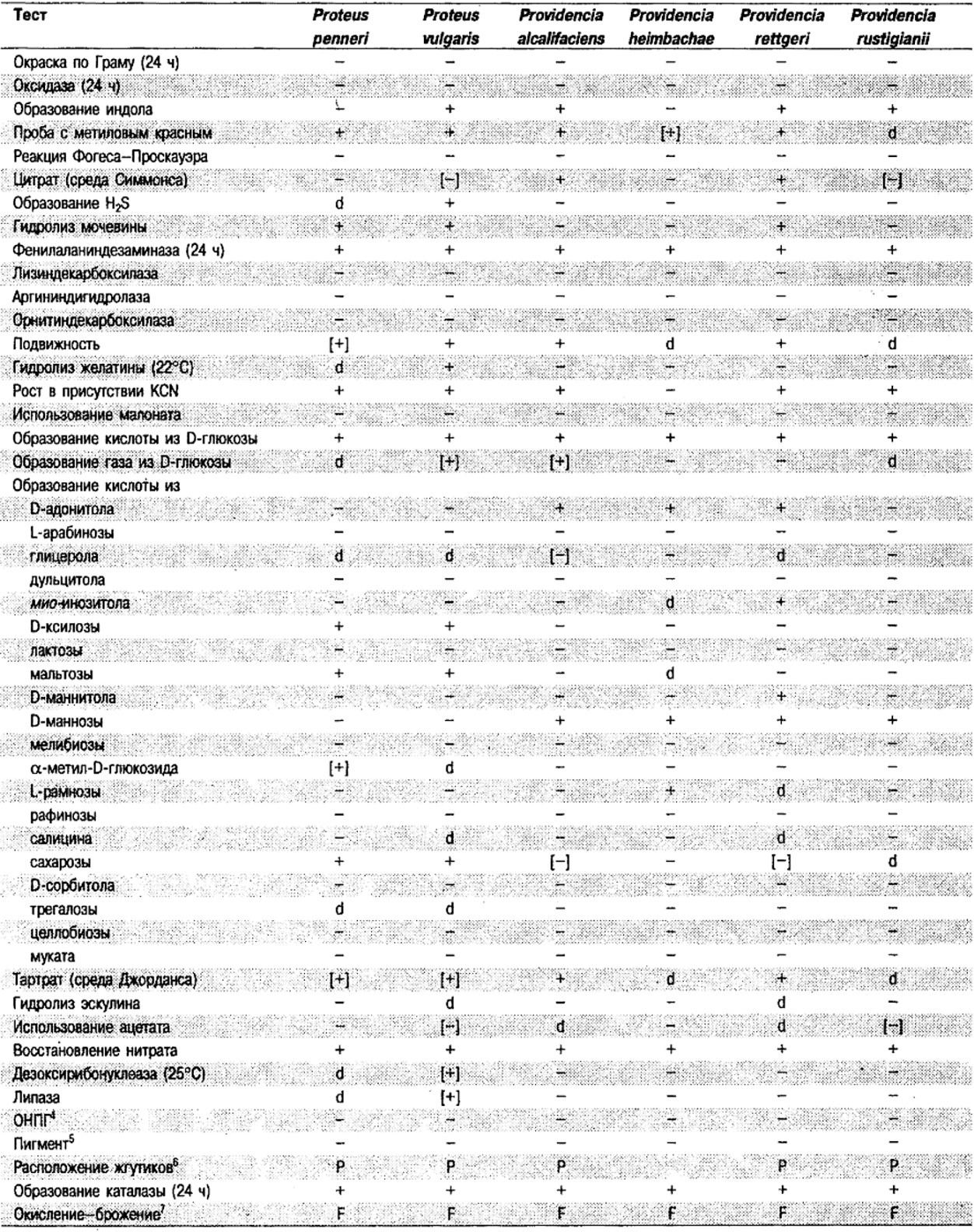
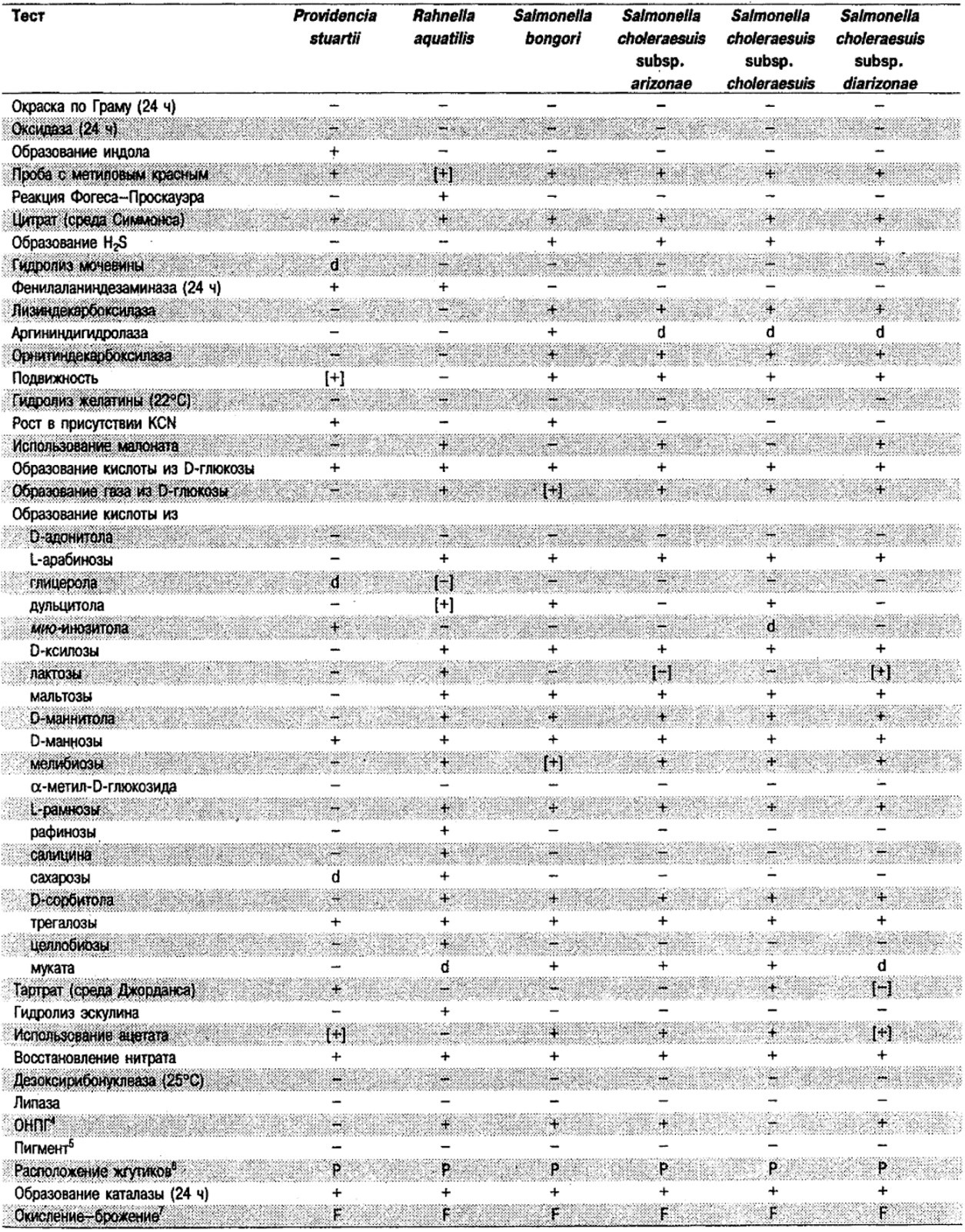
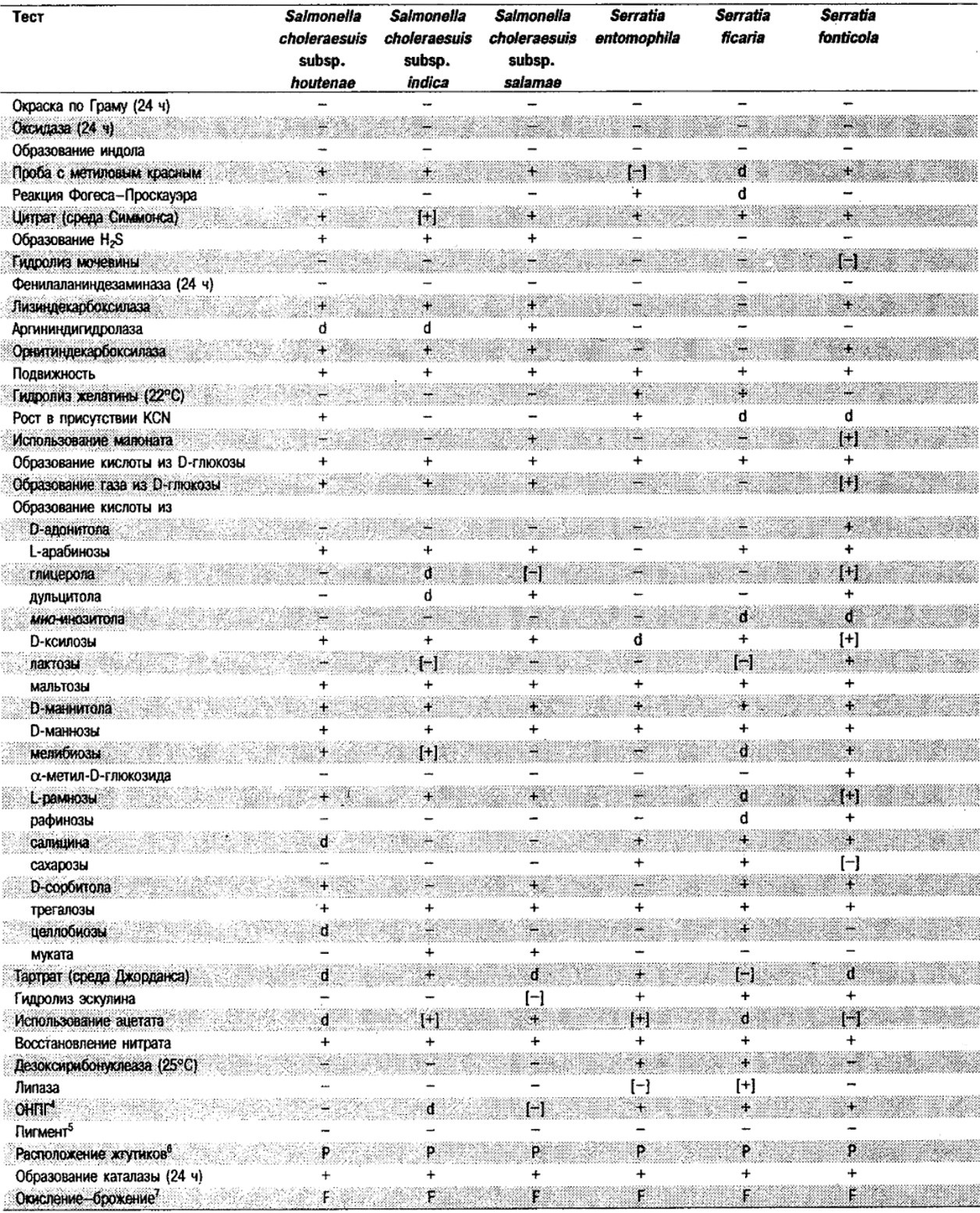
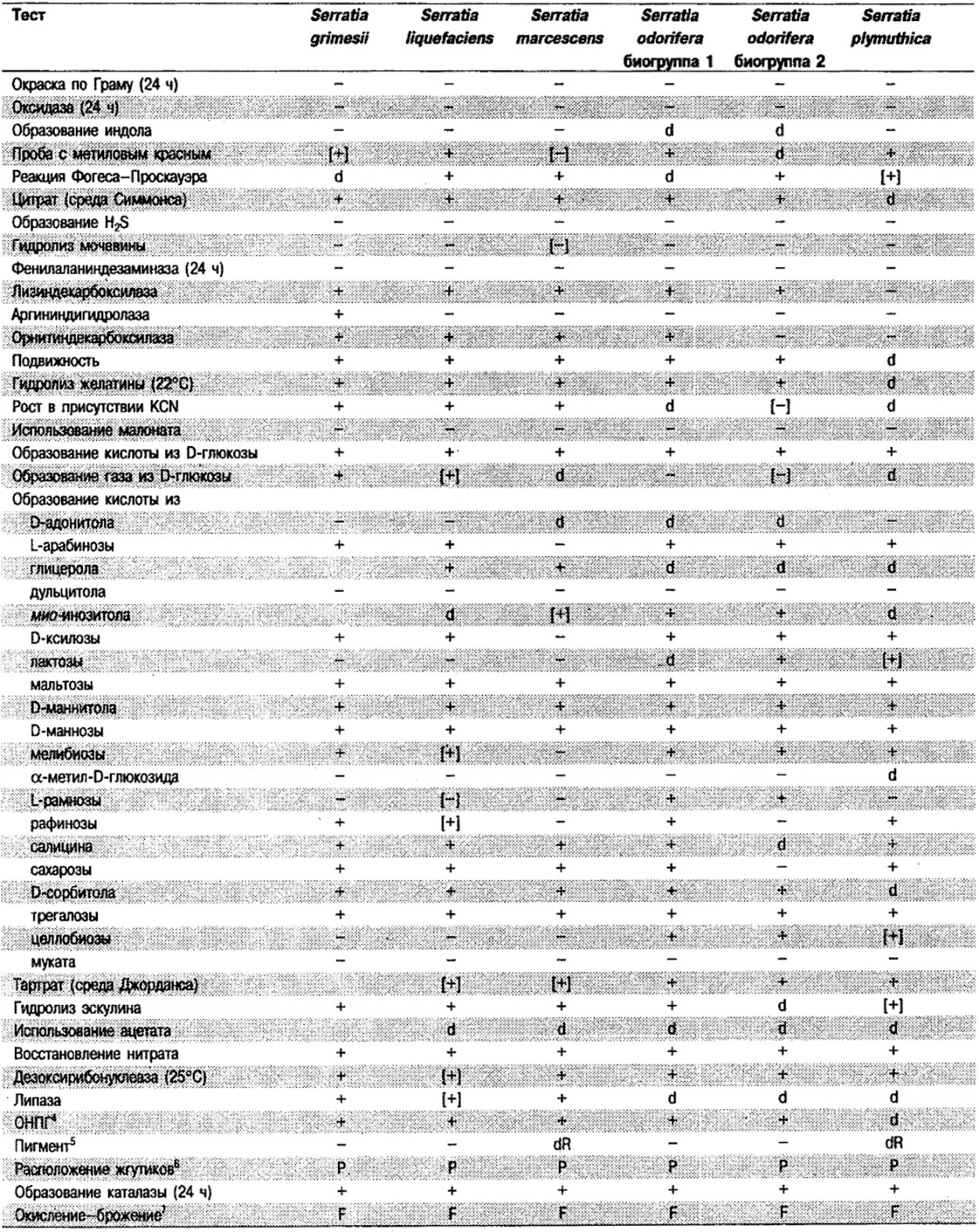
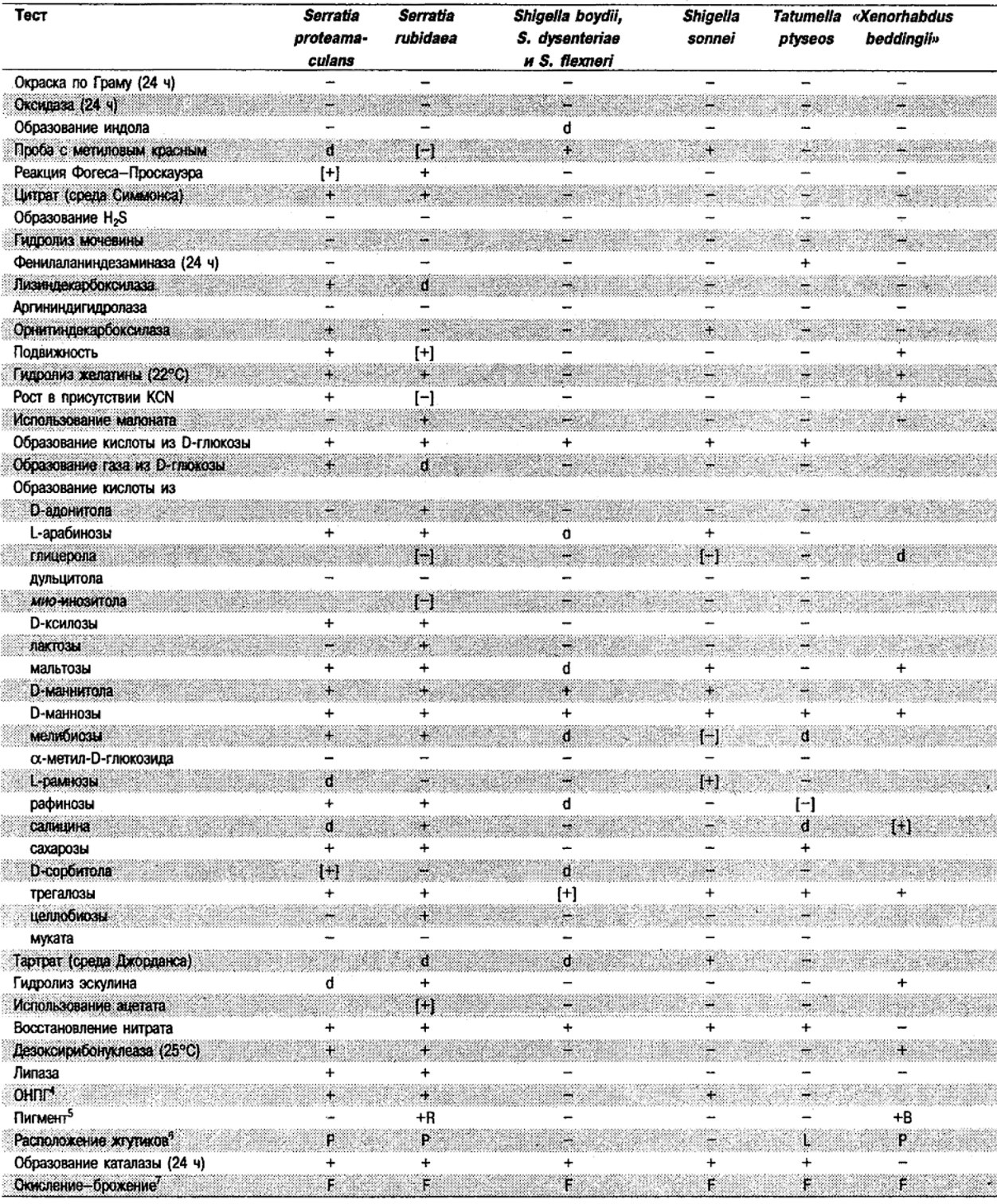
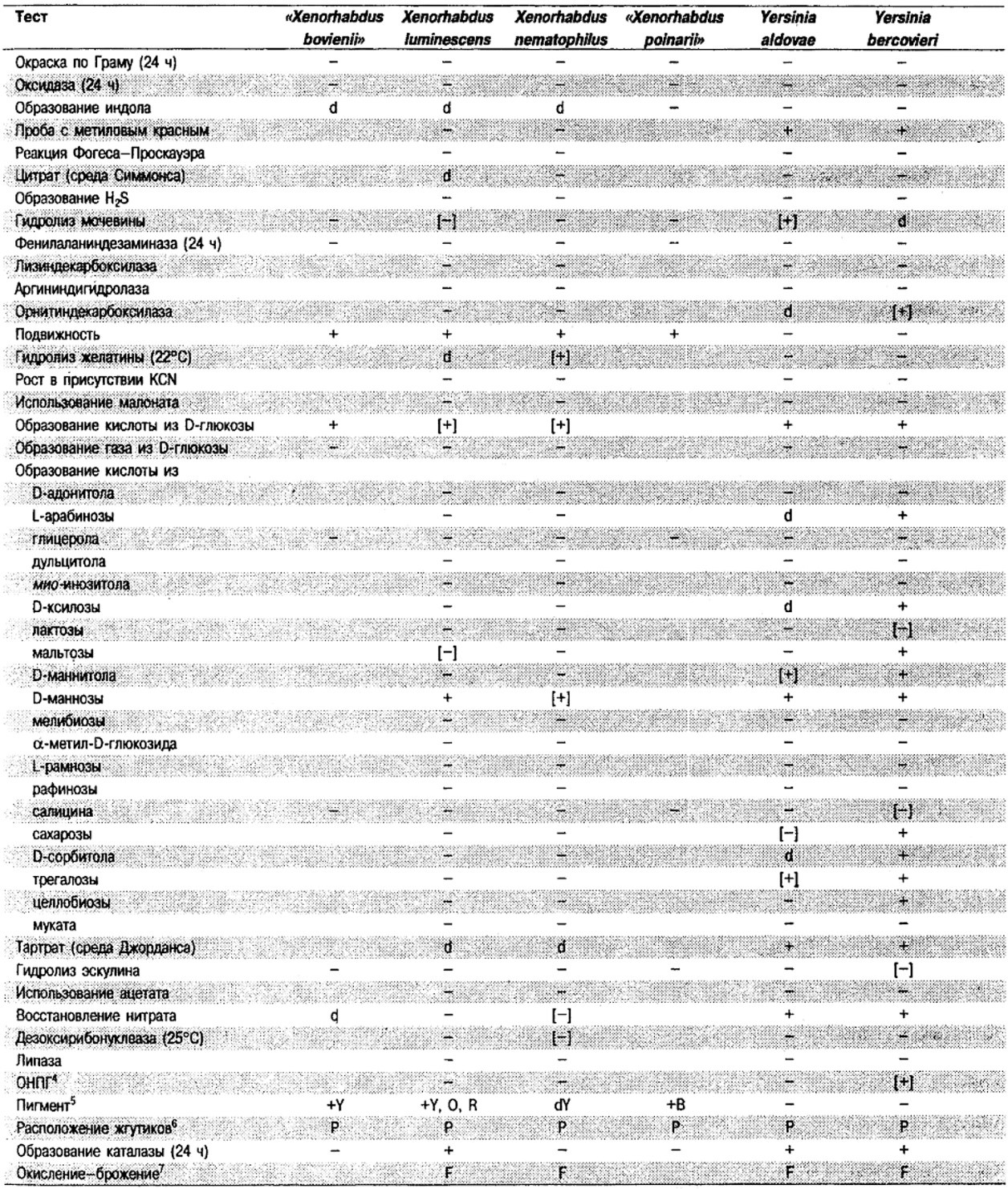
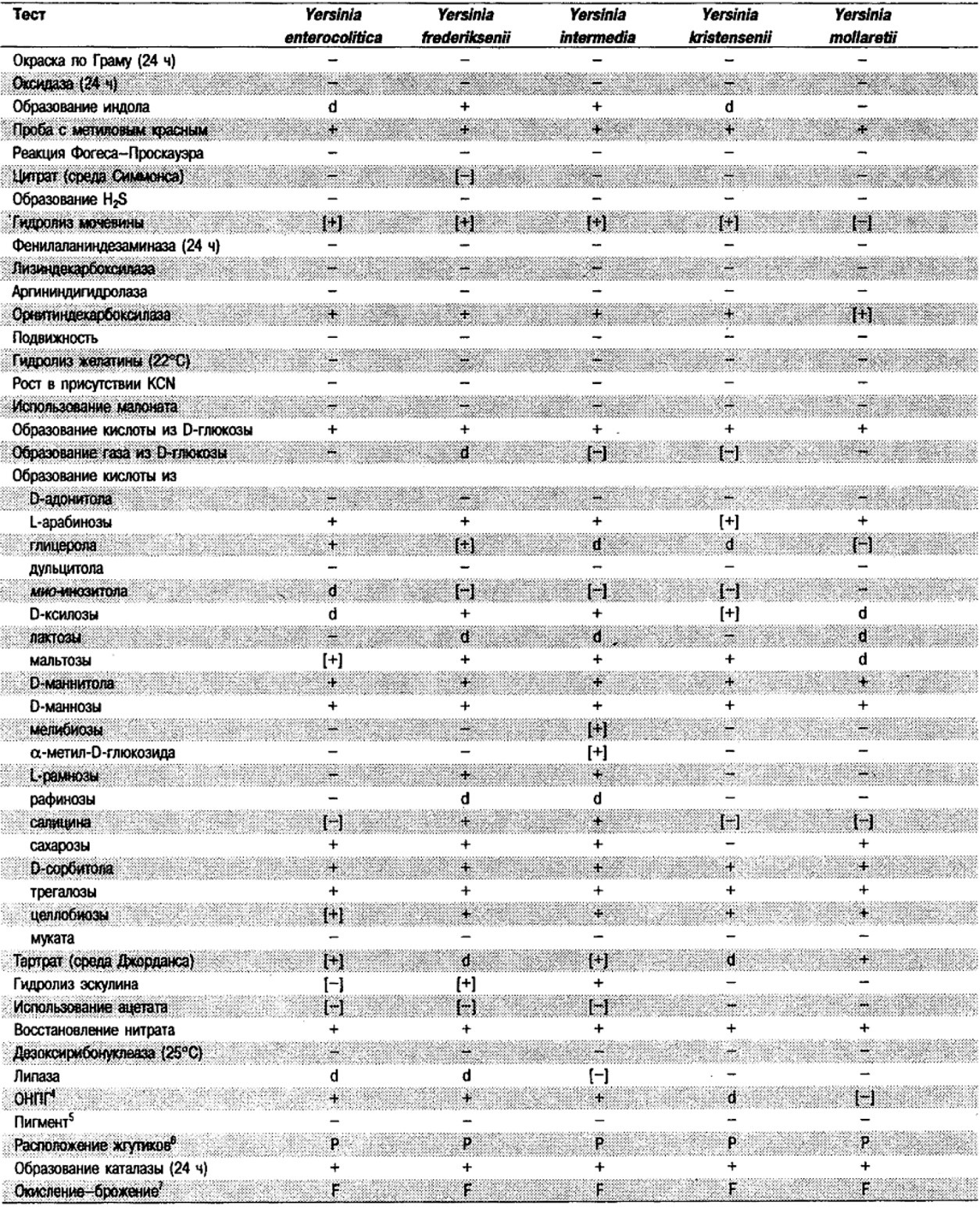
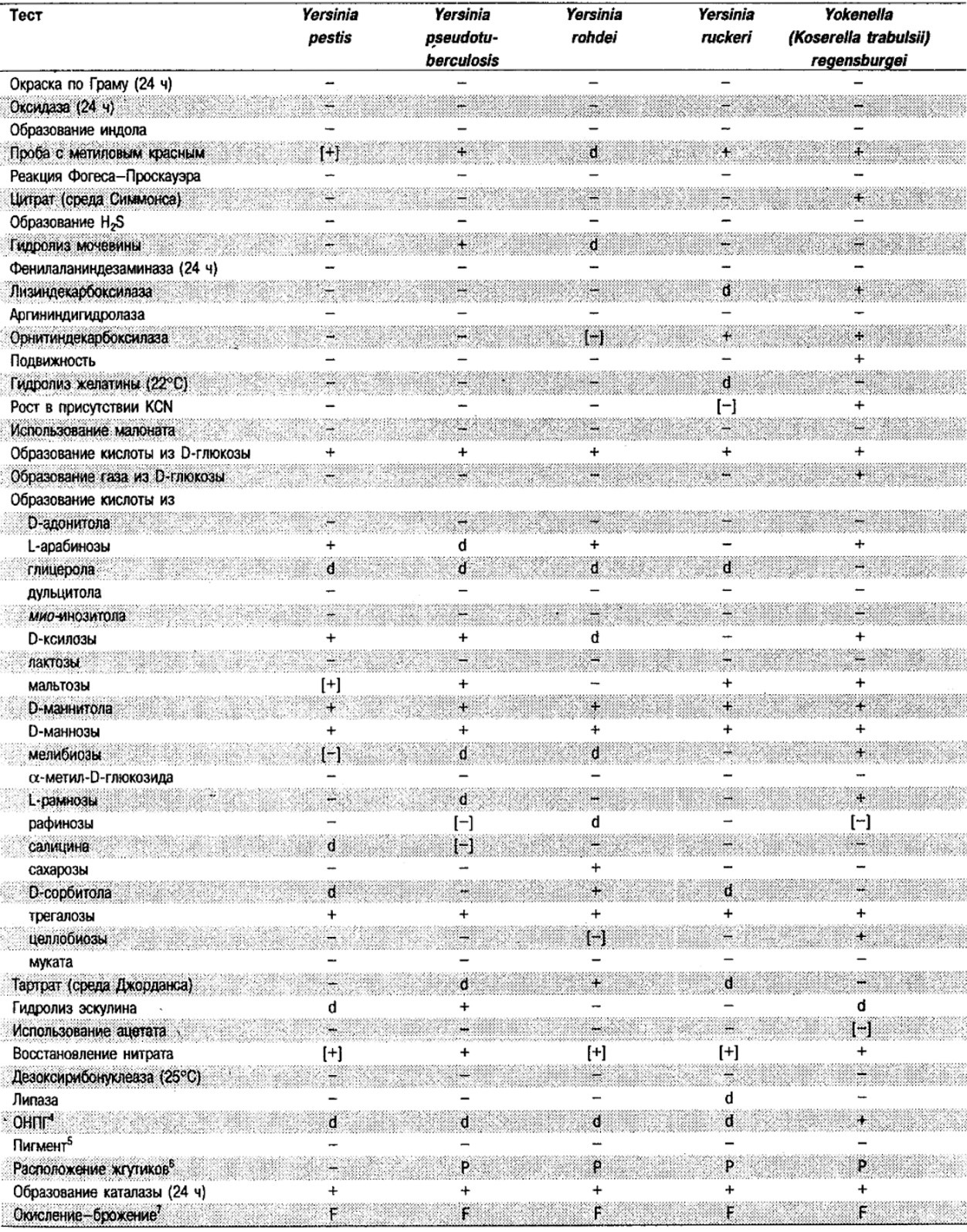
От редакции: Табл. 5.2 составлена на основе соответствующей таблицы в «Bergey's Manual of Systematic Bacteriology)», но с учетом новых данных, В некоторых случаях согласно новым сведениям изменены данные о процентном соотношении результатов реакций, так что оно подпадает под другую категорию (например вместо «d» – [+]). Почти все данные о реакциях приведены для стандартных условий: инкубация 48 ч при 37°С. Ряд видов дает (в одном или нескольких тестах) положительную реакцию с задержкой (на 3 и более суток; хороший пример – разжижение желатины). Представители ряда видов, включая многие йерсинии, эрвинии и бактерии Xenorhabdus, при 25–30°С метаболически более активны, чем при 37°С (пример – подвижность у йерсиний).
Род Arsenophonus
От редакции: Род Arsenophonus не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1991 г. (Werren et al., Int. J. Syst., Bacteriol. 41: 563–565) и включает один вид А. nasoniae.
Палочки, в молодых культурах иногда нитевидные, 0,4–0,6 х 7–10 мкм. Грамотрицательные. Неподвижные; факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура для роста 30°С. D-глюкозу и другие углеводы катаболизируют с образованием кислоты, но не газа. Оксидазоотрииательные; каталазоположительные; индолотрицательные, проба с метиловым красным и реакция Фогеса-Проскауэра отрицательные. Результаты тестов на лизиндекарбоксилазу и аргининдигидролазу отрицательные (роста нет). Нитрат не восстанавливают, H2S не образуют. Не растут на агаре Макконки. Встречаются у самок ос. Патогенны для ос – вызывают гибель яиц, предназначенных для развития мужских особей.
Типовой (и единственный) вид: Arsenophonus nasoniae.
Признаки вида: См. табл. 5.2.
От редакции: Организм высокотребователен в отношении питательных веществ. Для определения биохимических признаков нужны особые (необычные) среды.
Колонии мукоидные, серо-белые, округлые, выпуклые, с цельным краем. Для роста в среде, используемой при определении признаков, необходимо присутствие 1% протеозы-пептона (Werren et al., Int. J. Syst. Bacteriol. 41: 563–565, 1991). Обнаруживают отрицательную реакцию или не дают роста в тестах на: дезоксирибонуклеазу, гидролиз эскулина, рост за счет использования малой ата, гидролиз Твина-20 и -80 и уреазу. Разжижают желатину. Образуют кислоту из фруктозы и сахарозы, но не из адонитола, L-арабинозы, глицерола, дульцитола, D-ксилозы, жио-инозитола, лактозы, мальтозы, D-маннитола, рафинозы, трегалозы и целлобиозы. Представители рода растут на средах, содержащих D-глюкозу, фруктозу и сахарозу и слабо на средах с целлобиозой, мальтозой, трегалозой и D-ксилозой; не растут на средах с адонитолом, L-арабинозой, глицеролом, дульцитолом, мио-инозитолом, лактозой, D-маннитолом и рафинозой.
Род Budvicia
От редакции: Род Budvicia не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1983 г. (Aldova et al., Zentralbl. BakterioL Parasitenkd. [nfektionskr. Hyg. Abt. 1 Orig. Reihe A 254: 95–108) и одобрен в 1985 г, (Bouvetet al. Int. J. Syst. Bacteriol. 35: 60–64). Род включает один вид В. aquatica.
Прямые палочки. Грамотрицательные. Подвижные за счет перитрихнальных жгутиков в условиях роста при 22°С. Встречаются штаммы, неподвижные при 22°С, и менее половины штаммов обнаруживают подвижность при 37°С. Факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура для роста 30–37°С. Катаболизируют D-глюкозу и другие углеводы, включая L-арабинозу (большинство штаммов), D-ксилозу и L-рамнозу с образованием кислоты и небольшого количества газа или не образуют газ. Оксидазоотрицательные и катала зоположителъные. Индол не образуют. Проба с метиловым красным и реакция Фогеса-Проскауэра отрицательные. Цитрат не используют. По лизиндекарбоксилазе, аргининдигидролазе и орнитиндекарбоксилазе отрицательные. Большинство штаммов образует H2S. Восстанавливают нитрат. Выделены в основном из пресной воды, отдельные изоляты – из кала человека и из животных. Медицинское значение неизвестно.
Типовой (и единственный) вид: Budvicia aquatica.
Штаммы Budvicia вначале принимали за нетипичные штаммы Citrobacter, однако их легко различить (табл. 5.6). Биохимические признаки, отличающие Budvicia от H2S-положительных родов Leminorella и Pragia, указаны в табл. 5.22 и 5.26. Отличия от родов с отрицательной реакцией в тестах на использование лизина, аргинина и ор- нитина приведены в табл. 5.2.
Признаки вида: Как для рода, табл. 5.2.
Род Buttiauxella
От редакции: Род Buttiauxella упомянут как «Другой организм» после описания рода Kluyvera, но не включен как отдельный род в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1981 г. (Ferragut el ai Zentralbl. Bakteriol. Parasitenkd. Infektionskr. Hyg. Abt. 1 Orig. С 2: 33–34; Int. J. Syst. Bacteriol. 32: 266–268, 1982) и включает один вид В. agrestis.
Мелкие л ал очки, подвижные за счет перитрихи- альных жгутиков. Грамотрицательные. Факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная для роста температура 30–37°С. D-глюкозу и другие углеводы катаболизируют с образованием кислоты и газа. Оксидазоотрнцательные и каталазоположительные. Индол не образуют; реакция Фогеса-Проскауэра отрицательная; проба с метиловым красным и результат теста на среде Симмонса с цитратом положительные; по лизиндекарбоксилазе положительные. Восстанавливают нитрат. Сбраживают L-арабинозу, D-ксилозу, лактозу, мальтозу, D-маннитол, D-маннозу, мелибиозу, L-рамнозу, рафинозу, салицин, трегалозу и целлобиозу. Выделены из пресной воды; у человека или животных не обнаружены.
Типовой (и единственный) вид: Buttiauxella agrestis.
Признаки вида: Фенотипически вид сходен с родом Kluyvera. Тесты, позволяющие дифференцировать эти роды, приведены в табл. 5.3.
Таблица 5.3. Отличия Buttiauxella agresbs от Enteric groups 63, 64 и рода Kluyvera1

Дифференциация видов рода Buttiauxella: В табл. 5.3 приведены отличия В. agrestis от штаммов, обозначенных как Энтерогруппа 63 (Enteric Group 63) и Энтерогруппа 64 (Enteric Group 64) (Farmer et al., J. Clin, Microbiol. 21: 46–76, 1985) и представляющих геновиды, близкородственные В. agrestis. Признаки В. agrestis приведены в табл. 5.2.
Род Cedecea
Палочки, 0,5–0,6 х 1–2 мкм. Грамотрицательные. Клетки большинства штаммов подвижные. Факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура 37°С. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты и обычно газа. Оксидазоотрицательные и каталазоположительные; индолотрицателькые; проба с метиловым красным обычно положительная; реакция Фогеса-Проскауэра различная. Растут на среде Симмонса с цитратом. По лизицдекарбоксилазе отрицательные, по липазе (кукурузное масло) положительные. Восстанавливают нитрат. Сбраживаемые углеводы включают D-арабитол, мальтозу, D-маннитол, D-маннозу, салицин, трегалозу и целлобиозу. Источником выделения служит клинический материал, причем более 50% штаммов выделено из дыхательных путей. В отдельных случаях вызывают оппортунистические инфекций.
Типовой вид: Cedecea davisae.
Штаммы Cedecea первоначально рассматривали как промежуточные формы между типичными штаммами Serratia и S. fonticola. Кроме того, они в некоторой степени сходны с Emngeila. Реакции, позволяющие дифференцировать Cedecea от указанных родов, приведены в табл. 5.5.
Признаки видов рода Cedecea: Виды Cedecea, включая не имеющие названия «Виды 3 и 5», дифференцируются по реакциям, перечисленным в табл. 5.4. Признаки видов Cedecea приведены в табл. 5.2.
Таблица 5.4. Дифференциация видов рода Cedecea1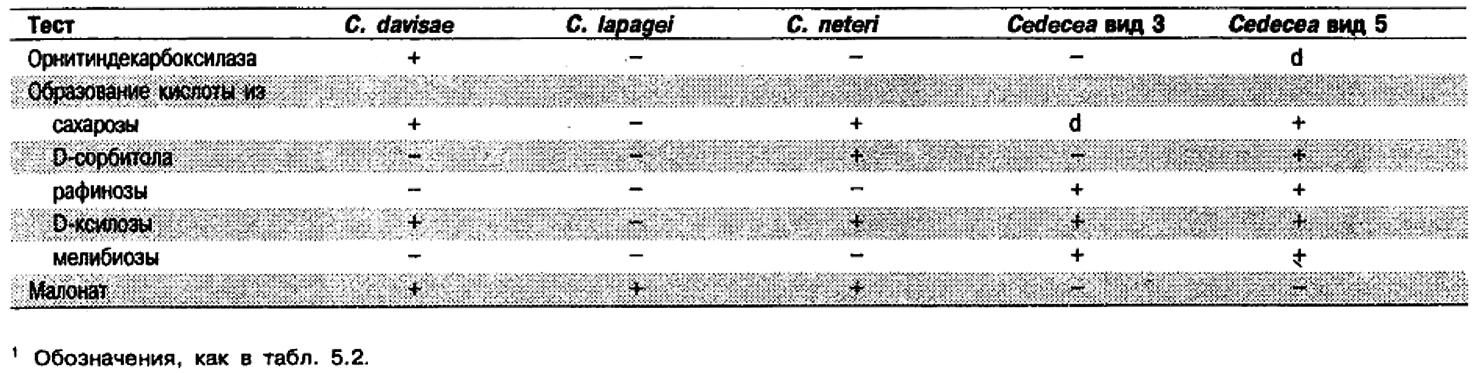
Таблица 5.5. Отличия Cedecea от Ewingella и Serratia1
Род Citrobacter
Прямые палочки, 1 х 2–6 мкм, одиночные и в парах. Грамотрицательные. Обычно подвижные за счет перитрихиальных жгутиков. Факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура 37°С. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты и газа. Оксидазоотрицательные и каталазоположительные; проба с метиловым красным положительная; обычно цитратположительные, реакция Фогеса-Проскауэра отрицательная; по лизиндекарбоксилазе отрицательные. Восстанавливают нитрат. Сбраживаемые углеводы обычно включают L-арабинозу, глицерол, D-ксилозу, мальтозу, D-маннитол, L-рамноэу, D-сорбитол, трегалозу и целлобиозу. Встречаются в фекалиях человека и животных, возможно как представители нормальной микрофлоры кишечника. Во многих случаях выделены из клинического материала как возбудители оппортунистических инфекций. С. diversus может вызывать менингит новорожденных. Обнаружены также в почве, воде, сточных водах и пищевых продуктах.
Типовой вид: Citrobacter freundii.
Признаки, отличающие Citrobacter от биохимически сходных родов, см. в табл. 5.6.
Дифференциация видов рода Citrobacter: См. табл. 5.7. Полностью признаки перечислены в табл. 5.2.
От редакции; Названия Citrobacter koseri и Levinea malonatica – это синонимы С. diversus, a Levinea amalonatica – синоним С. amalonaticus. Эти названия продолжают появляться в литературе и их следует считать синонимическими.
Таблица 5.6. Отличия рода Citrobacter от других родов1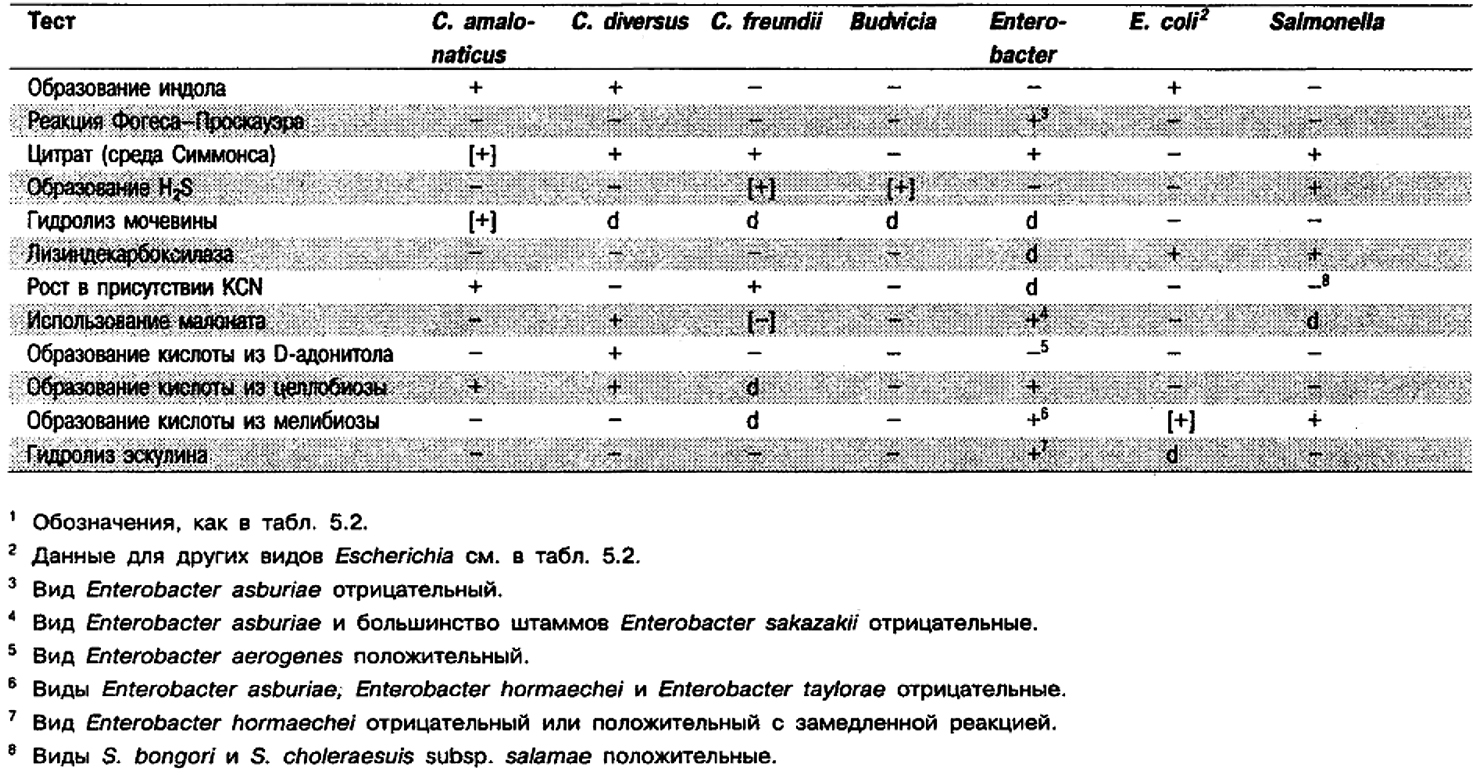
Таблица 5.7. Биохимическая дифференциация видов рода Citrobacter1
Род Edwardsiella
Мелкие прямые палочки, 1 х 2–3 мкм. Грамотрицательные. Подвижные за счет перитрихиальных жгутиков (клетки Е. ictaluri подвижны при 25 °С, но не при 37°С). Факультативные анаэробы; хемоорганотрофы, обладающие и дыхательным, и бродильным типами метаболизма. Оптимальная температура для роста 37°С; исключение составляют бактерии Е. ictaluri, для которых предпочтительна более низкая температура. Катаболизируют D-глюкозу и некоторые другие углеводы с образованием кислоты и часто газа, но по сравнению с большинством родов семейства Enterobacteriaceae не активны. Оксидазоотрицательные; каталазоположительные, реакция Фогеса-Проскауэра и результаты теста на среде Симмонса с цитратом отрицательные. Как правило, обладают активностями лизиндекарбоксилазы и орнитиндекарбоксилазы. Восстанавливают нитрат. Представители всех видов сбраживают мальтозу и D-маннозу. Наиболее часто встречаются в кишечнике у пойкилотермных животных и в их среде обитания, особенно в пресной воде, но обнаружены также и у гомойотермных животных и человека. Патогенны для угрей, зубаток и других животных, в редких случаях вызывают оппортунистические инфекции у человека.
Типовой вид: Edwardsiella tarda.
Биохимически бактерии Edwardsiella частично сходны с Escherichia coli, представителями Shigella и Salmonella, но их легко отличить по результатам полного набора биохимических тестов (табл. 5.8).
Дифференциация видов рода Edwardsiella: Дифференцирующие признаки видов Edwardsiella представлены в табл. 5.9. Дополнительные биохимические данные приведены в табл. 5.2.
Таблица 5.8. Отличия видов рода EdwardsieHa от других родов1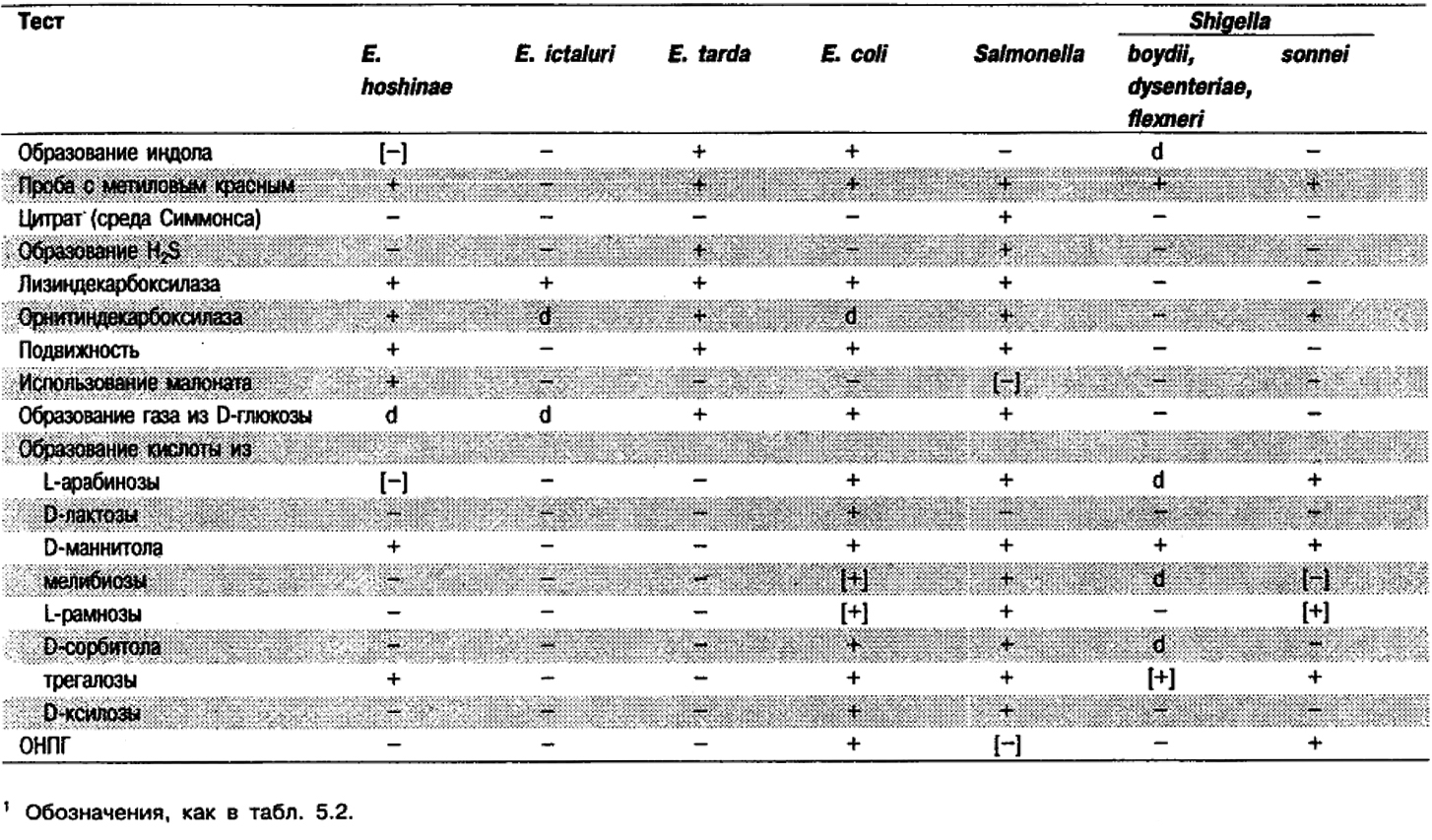
Таблица 5.9. Биохимическая дифференциация видов рода Edwardsiella1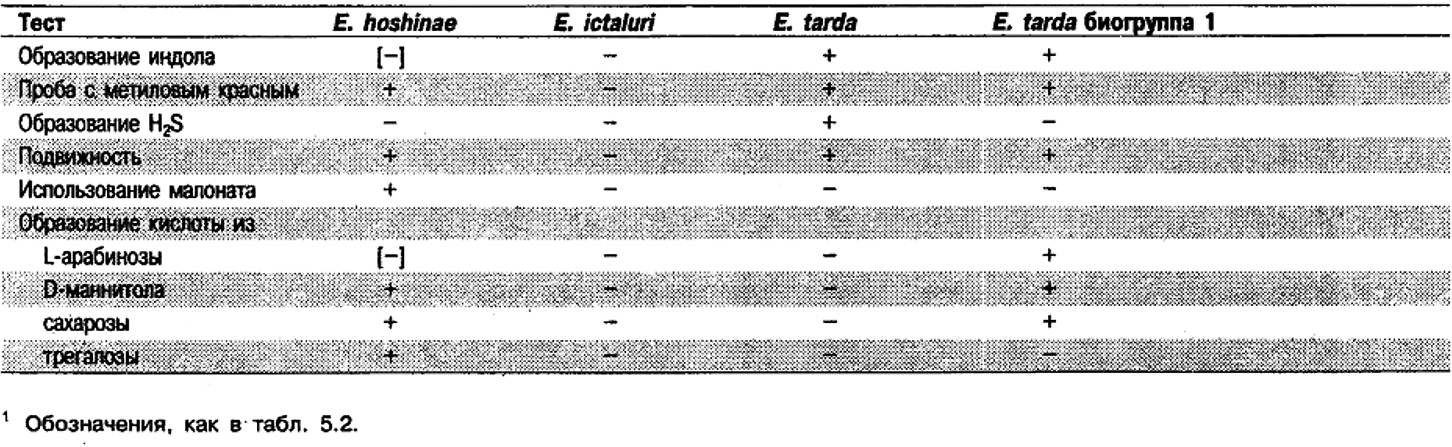
Род Enterobacter
Прямые палочки, 0,6–1,0 х 1,2–3,0 мкм, грамотрицательные. Подвижные за счет перитрихиальных жгутиков (за исключением Е. asburiae). Факультативные анаэробы; хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальный диапазон температуры 30–37°С. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты и газа. Индол отрицательные. У большинства штаммов реакция Фогеса-Проскауэра и результат теста на среде Симмонса с цитратом положительные. Проба с метиловым красным варьирует. Лизинотрицательные (за исключением Е. gergoviae) и орнитинположительные (за исключением Е. agglomerans), Большинство штаммов обычно использует малонат и медленно разжижает желатину (3–14 сут). H2S, дезоксирибонуклеазу и липазу не образуют. Все или большинство штаммов сбраживают следующие углеводы: L-арабинозу, мальтозу, D-маннитол, D-маннозу, салицин, трегалозу и целлобиозу. Широко распространены в природе: встречаются в пресной воде, почве, сточных водах, на растениях, овощах, а также в фекалиях человека и животных. Некоторые виды, в основном Е. cloacae, S. sakazakii, Е. aerogenes, Е. agglomerans и Е. gergoviae вызывают оппортунистические инфекции – ожоговые, раневые и мочевыводящих путей, а в некоторых случаях септицемию и менингит.
Типовой вид: Enterobacter cloacae.
Признаки, по которым виды Enterobacter отличаются от клебсиелл, приведены в табл. 5.10 и 5.11. В табл. 5.12 перечислены признаки, позволяющие отличить Enterobacter от Klebsiella, Hafnia и Serratia.
Дифферепция видов рода Enterobacter См. табл. 5.13.
Таблица 5.10. Отличия видов рода Enterobacter от клебсиелл1 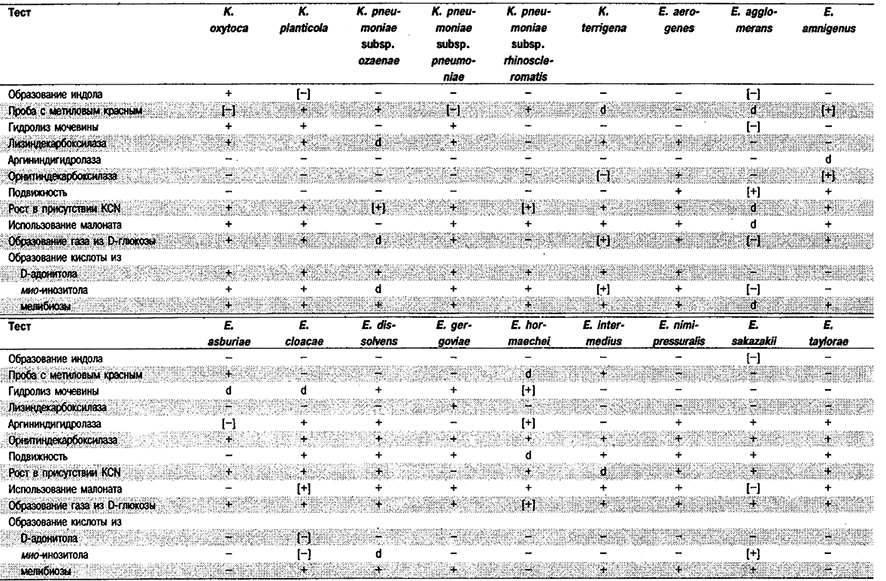
Таблица 5.11. Дополнительные тесты для различения видов Enterobacter и клебсиелл 1,2 
Таблица 5.12. Дифференциация Enterobacter и близких родов1 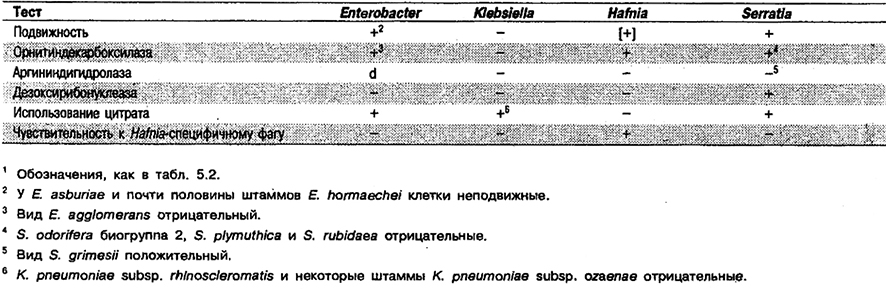
Табл. 5.13. Биохимическая дифференциация видов рода Enterobacter1 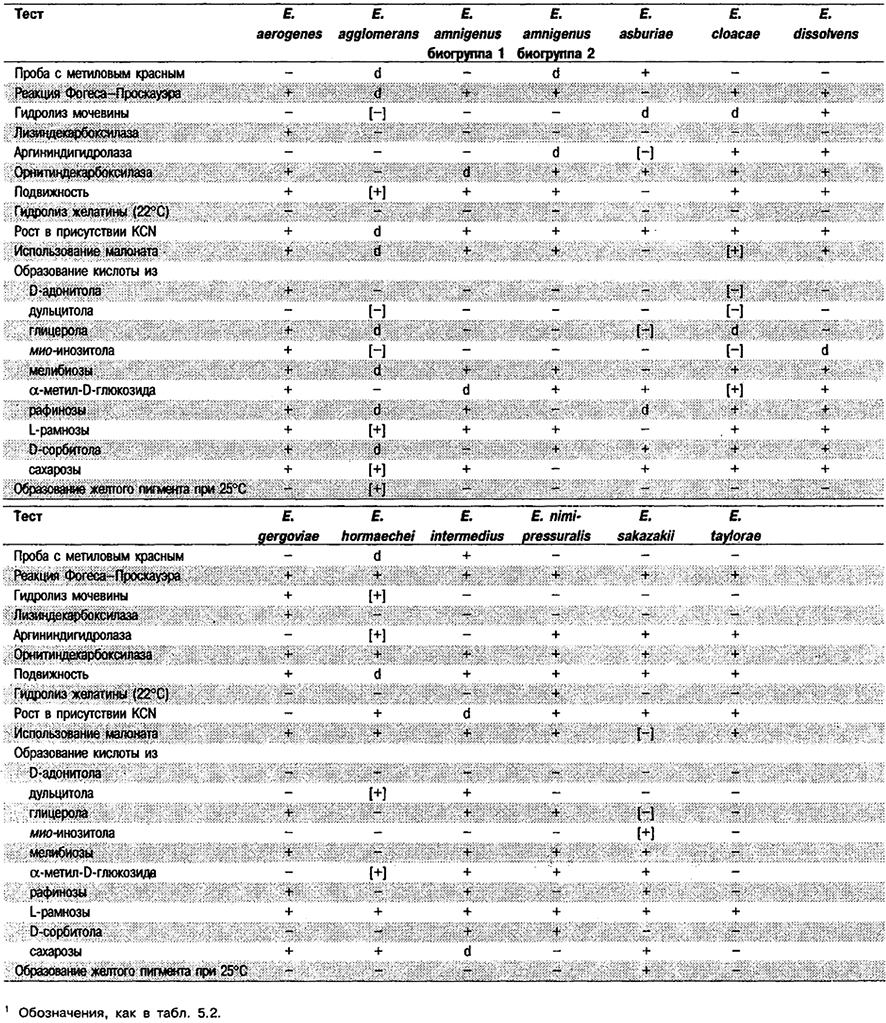
От редакции; Виды, перечисленные ниже, не приведены в «Bergey's Manual of Systematic Bacteriology» или кратко упомянуты в разд. «Другие организмы, относящиеся к роду Enterobacter». Enterobacter amnigenus описан в 1981 г, (Isard et al Int. J. Syst. Bacteriol. 31: 35–42); Enterobacter asburiae описан в 1986 г. (Brenner et al. J. Clin. Microbiol. 23: 1114–1120; Int. J. Syst. Bacteriol. 38: 220–222, 1988); Enterobacter cancerogenus перенесен в род Enterobacter из рода Erwinia в 1988 г. (Dickey, Zumoff, Int. J. Syst. Bacteriol. 38: 371–374; Enterobacter dissolvents перенесен в род Enterobacter из рода Erwinia в 1986 г. (Brenner et a. J. Clin. Microbiol. 23: 1114–1120; Int. J. Syst. Bacteriol. 38: 220–222); Enterobacter hormaechei описан в 1989 г. (О'Нага et al, J. Clin. Microbiol. 27: 2046–2049; Int. i, Syst. Bacteriol, 40: 105–106, 1990); Enterobacter intermedium описан в 1980 г. (Isard et al., Zentralbl. Bakteriol. Parasitenkd. Infektionskr. Hyg. Abt. 1 Orig. Reihe C, 1: 51-60; Int. J. Syst. Bacteriol. 30: 601, 1980). Род специфического эпитета «intermedium» не согласуется с названием рода и поэтому был изменен на intermedin (von Gravenitz, Int. J. Syst. Bacteriol. 40: 211, 1990). Enterobacter nimipressuralis перенесен в род Enterobacter из рода Erwinia в 1986 г. (Brenner et al., J. Clin. Microbiol. 23: 1114–1120; Int. J. Syst. Bacteriol. 38: 220–222, 1988); Enterobacter taylorae описан в 1985 г. (Farmer et al. J, Clin, Microbiol. 21: 77–81; Int. J. Syst. Bacteriol. 35: 223–225, 1985). Enterobacter agglomerans – это старший Субъективный синоним Erwinia herbicola и Erwinia milletiae (Beji et al., Int. J. Syst. Bacteriol. 38: 77–88, 1988). Предложено перенести Enterobacter agglomerans в новый род Pantoea (см. ниже). Название Enterobacter cancerogenus было предложено как старший субъективный синоним Enterobacter taylorae (Gri- mont, Ageron, Res. Microbiol. 140: 459–465, 1989).
Дифференцирующие признаки видов Enterobacter приведены в табл. 5.13. Дополнительные тесты указаны в табл. 5.2.
Род Erwinia
От редакции: Род Erwinia изучен в основном фитопатологами. Рекомендуется использовать тесты, указанные в табл. 5.13–5.15 и успешно применяемые фитопатологами. Многие тесты и условия их проведения существенно отличаются от тех, что перечисле ны для других представителей Enterobacteriaceae (табл. 5.2). Биохимические признаки эрвиний в табл, 5.2. представлены для сравнения с другими родами. Несколько видов иногда помещали в род Pectobacterium. Род Pectobacterium более не используется, хотя и был одобрен.
Прямые палочки, 0,5–1,0 х 1–3 мкм, одиночные, в парах и иногда в коротких цепочках. Грамотрицательные. Подвижные за счет перитрихиальных жгутиков (за исключением Е, stewartii). Факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура для роста 27–30°С. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты; газ большинство видов не образуют. Оксидазоотрицательные; каталазоположительные; по лизиндекарбоксилазе, аргининдигидролазе и орнитиндекарбоксилазе отрицательные. Нитрат большинство видов не восстанавливает. Сбраживают галактозу, β-метилглюкозид, сахарозу и фруктозу; обычно сбраживают D-маннитол, D-маннозу, рнбозу и D-сорбнтол; редко сбраживают адоннтол, декстрин, дульцитол и мелицитозу. Как источники углерода и энергии используют ацетат, глюконат, малат, сукцинат и фумарат, но не бензоат, оксалат или пропионат. Ассоциированы с растениями как патогенные организмы, сапрофиты или компоненты эпифитной флоры. Очень редко эрвинии выделяют у человека.
Типовой вид: Erwinia amylovora.
Обстоятельного сравнения Erwinia с другими родами Enterobacteriaceae не проведено, поскольку слишком малое число неэрвиний исследовано методами, используемыми для эрвиний и, наоборот, слишком малое число эрвиний изучено с помощью рутинного набора тестов, применяемых для других представителей Enterobacteriaceae. Таким образом, данные, приведенные в табл. 5.2 для дифференциации эрвиний от бактерий других родов, возможно, трудно сравнивать из-за несовпадения использованных для их получения методов.
Дифференциация видов рода Erwinia: См. табл. 5.14, 5.15, 5.16 и 5.2.
Таблица 5.14. Кулыурапьные, физиологические и биохимические признаки видов рода Erwinia 1,2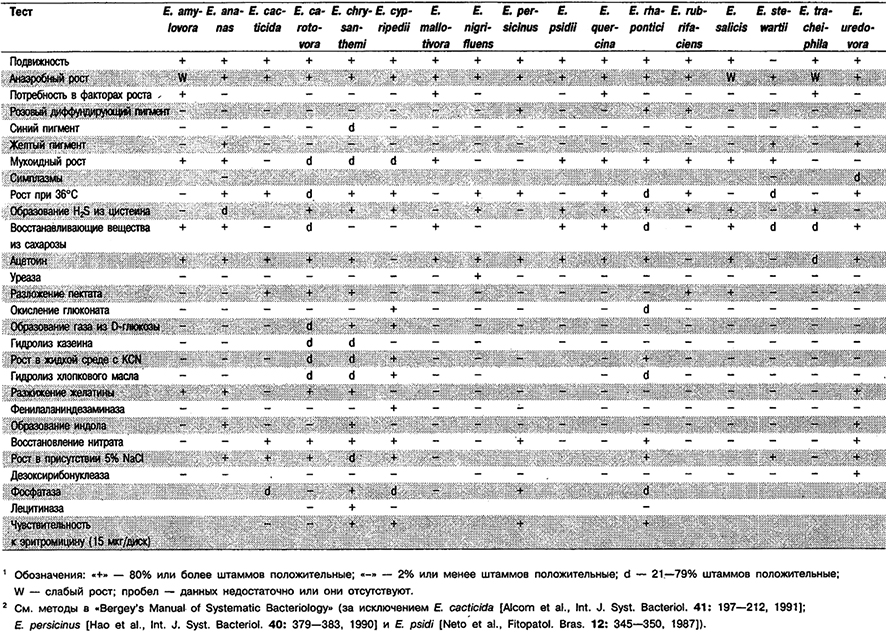
Таблица 5.15. Образование кислот из органических соединений у видов рода Erwinia 1,2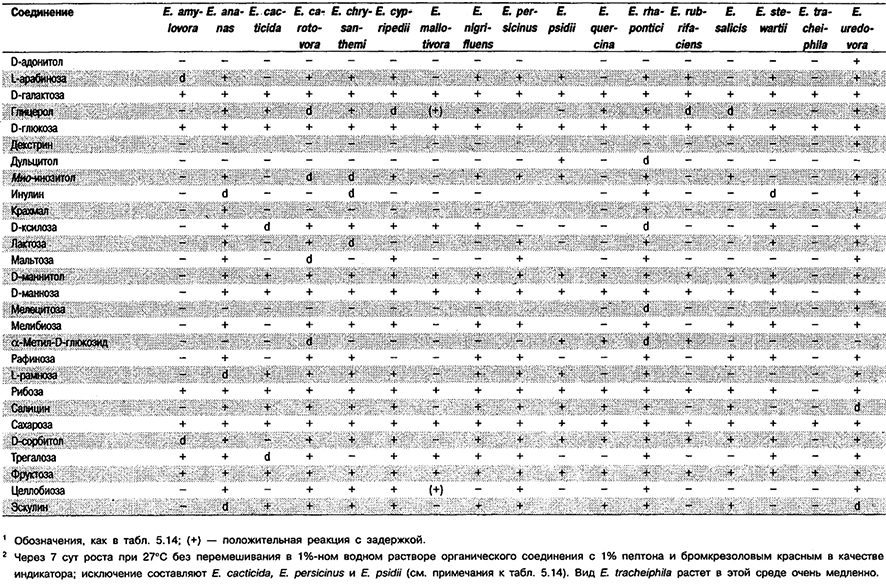
Таблица 5.16. Использование некоторых органических соединений
в качестве источников углерода и энергии у видов рода Erwinia 1,2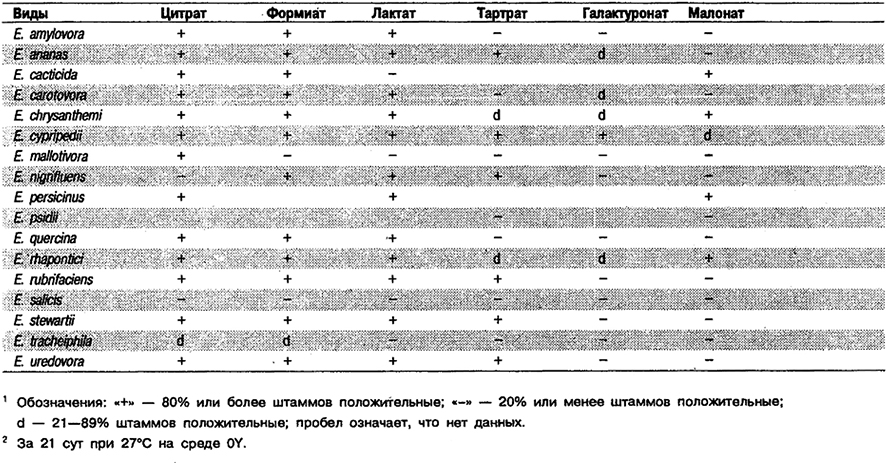
Виды, перечисленные ниже, не приведены в «Bergey's Manual of Systematic Bacteriology». Вид Erwinia cacticida описан в 1991 г. (Alcorn et al., Int. J. Syst. Bacteriol. 41: 197–212); вид Erwinia carotovora subsp. wasabiae описан в 1987 г. (Goto, Matsumoto, Int. J. Syst. Bacteriol. 37: 130–135); вид Erwinia persicinus описан в 1990 г. (Hao et al., Int. J. Syst. Bacteriol. 40: 379–383); Erwinia psidii описан в 1987 г. (Neto et al., Fitopatol. Bras, 12: 345–350; Int. J. Syst. Bacteriol. 38: 328, 1988).
Род Escherichia
Прямые палочки, 1,1–1,5 x 2,0–6,0 мкм, одиночные или в парах. Для многих штаммов характерны капсулы или микрокапсулы. Грамотрицательные. Подвижные за счет перитрихнальных жгутиков или неподвижные. Факультативные анаэробы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура 37 °С. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты и газа. Оксидазоотрииательные; каталазоположительные; проба с метиловым красным положительная; реакция Фогеса-Проскауэра отрицательная; обычно цитратотрицательные. Отрицательные по признакам образования H2S, гидролиза мочевины и активности липазы. Восстанавливают нитрат. Все или большинство штаммов сбраживают L-арабинозу, D-ксилозу, мальтозу, D-маннитол, D-маннозу, L-рамнозу и трегалозу. О-нитрофенил-β-D-галактопиранозидположительные. Встречаются как нормальная флора в нижнем отделе кишечника у гомойотермных животных, а в случае Е. blattae – у тараканов. Штаммы Е. coli, содержащие энтеротоксины и/или другие факторы вирулентности, включая инвазивность и факторы колонизации, вызывают желудочно-кишечные заболевания. Кроме того, Е. coli служит основной причиной инфекций мочевых путей и внутрибольничных инфекций, в том числе септицемии и менингита. Другие виды, за исключением Е. blattae, в редких случаях вызывают оппортунистические инфекции, причем обычно ассоциированы с раневыми инфекциями.
Типовой вид: Escherichia coli.
От редакции: Часто с целью идентификации и описания эпидемиологически патогенных штаммов вид Е. coli подразделяют по серологическим признакам или присутствию факторов вирулентности. Полное серотипирование включает определение соматических (О), капсульных (К) и жгутиковых (Н) антигенов.
Типичные по биохимическим признакам виды Escherichia нетрудно отличить от видов других родов (табл. 5,17), Однако в случае Е. coli существуют особые проблемы. Бактерии Е. coli и все шигеллы представляют один геновид, в котором шигеллы образуют биогруппы метаболически неактивных, неподвижных организмов. Метаболически неактивные штаммы Е. coli трудно отличить от шигелл. Особенно это относится к биогруппе штаммов Е, coli, представляющих нетипичные лактозоотрицательные, неподвижные и не образующие газ при сбраживании углеводов организмы. Другая биогруппа Е. coli характеризуется отсутствием активностей лизиндекарбоксилазы, аргининдигидролазы и орнитинде карбоксилазы, что делает ее сходной с Enterobacter (Pantoea) agglomerans и другими видами, также отрицательными по указанным признакам. В табл. 5.17 приведены дифференцирующие признаки, по которым можно отличить К coli и другие виды Escherichia от этих видов. И, наконец, штаммы Е. coli иногда обнаруживают нетипичные реакции в различных тестах, например на H2S, цитрат, уреазу, KCN, адонитол, инозитол и индол, поэтому прежде чем исключать Е. coli из рассмотрения, важно определить полностью весь биохимический профиль, а не только провести отдельные «ключевые» тесты.
Таблица 5.17. Отличия видов рода Escherichia от других видов1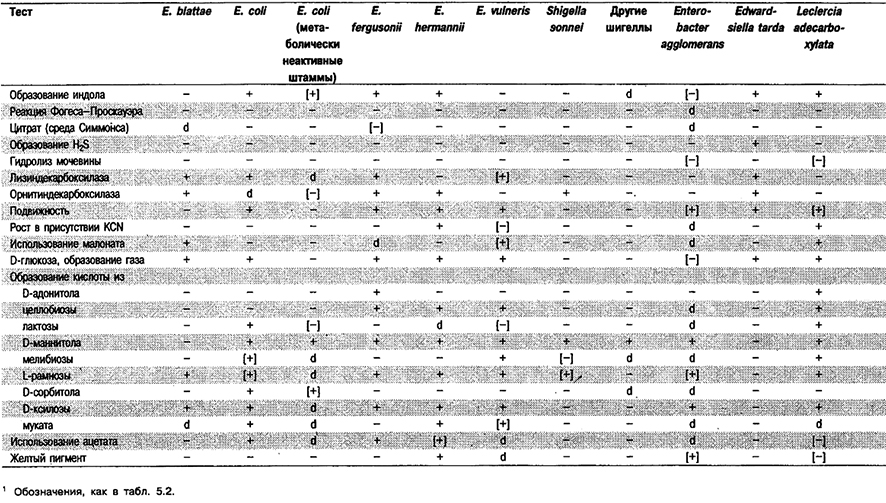
От редакции: Виды Escherichia fergusonii, Escherichia hermannii и Escherichia vulneris не приведены в «Bergey's Manual of Systematic Bacteriology». Вид Escherichia fergusonii описан в 1985 r. (Farmer et al., J. Clin. Microbiol. 21: 77–81; Int. J. Syst. Bacteriol. 35: 223–225, 1985). Вид Escherichia hermannii описан в 1982 г, (Brenner et al., J. Clin Microbiol. 15: 703–713; Int. J. Syst. Bacteriol. 33: 438– 440, 1983). Вид Escherichia vulneris описан в 1982 г. (Brenner et al., J. Clin. Microbiol. 15: 1113–1140: Int. J. Syst. Bacteriol. 33: 438-440, 1983).
Дифференциация видов рода Escherichia: Признаки, используемые для дифференциации видов Escherichia, представлены в табл. 5.18. Для сравнения в таблицу включены виды Obesumbacterium proteus и Leclercia adecarboxylata, поскольку первый из них наиболее близкородствен Escherichia по данным гибридизации ДНК, а второй был включен в род Escherichia. Полный перечень реакций приведен в табл. 5.2.
Таблица 5.18. Биохимическая дифференциация видов рода Escherichia и близких видов1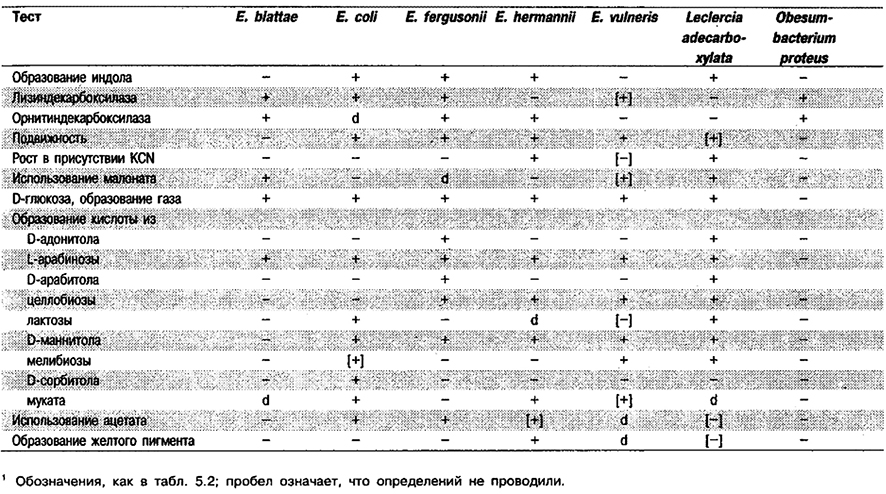
Род Ewingella
От редакции: Род Ewingella не приведен в «Вегgey's Manual of Systematic Bacteriology». Этот род описан в 1983 г. (Grimont et al., Arm. Microbiol. Inst. Pasteur 134 A: 39–52; Int. J. Syst. Bacteriol. 34: 91–92, 1984) и включает один вид Е, americana.
Прямые палочки, 0,6–0,7 х 1–1,8 мкм; по Граму окрашиваются отрицательно. Подвижные за счет перитрихнальных жгутиков. Факультативные анаэробы; хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура 37°С. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты. Оксидазоотрицательные; каталазоположительные; индолотрицательные; проба с метиловым красным положительная; реакция Фогеса-Проскауэра положительная. Способны расти на среде Симмонса с цитратом. По лизин- и орнитнцдекарбокенлазе, аргининдигидролазе, образованию H2S, уреазе и использованию малоната отрицательные. Восстанавливают нитрат. Сбраживаемые углеводы включают D-маннитол, D-маниозу и трeraлоз у. Источник выделения – клинический материал, наиболее часто мокрота, раны и кровь. В отдельных случаях вызывают оппортунистические инфекции.
Типовой (и единственный) вид: Ewingella americana.
Признаки вида: Представители Ewingella наиболее близки к видам Cedecea, от которых их можно отличить с помощью тестов, приведенных в табл. 5.5. Следует также дифференцировать Ewingella от других организмов, отрицательных по лизин- и орнитиндекарбоксилазе и аргининдигидролазе. Это можно легко сделать путем сравнения полных биохимических профилей, как в табл. 5.2.
Род Hafnia
Прямые палочки, ~1 х 2–5 мкм; по Граму окрашиваются отрицательно. Подвижные за счет перитрихнальных жгутиков; встречаются неподвижные штаммы. Факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальный диапазон температуры 30–37°С. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты и газа. Оксидазоотрицательные; каталазоположительиые; индол не образуют; на агаре Симмонса с цитратом не растут; реакция Фогеса-Проскауэра и проба с метиловым красным у большинства штаммов положительные. По лизин- и орнитиндекарбоксилазе положительные; по аргининдигидролазе отрицательные. H2S и уреазу не образуют; KCN-положительные. Восстанавливают нитрат. Сбраживают L-арабинозу, глкцерол, D-ксилозу, мальтозу, D-маннитол, D-маннозу, L-рамиозу и трегалозу. Встречаются в фекалиях человека и животных, в том числе птиц, сточных водах, почве, воде и молочных продуктах. Вызывают оппортунистические инфекции у человека, локализованные обычно в крови, мочевых путях или ранах, на фоне другого заболевания или при наличии провоцирующих факторов.
Типовой (и единственный) вид: Hafnia alvei.
От редакции: Организм, обозначенный как Биогруппа 1 Obesumbacterium proteus, в действительности представляет биогруппу метаболически неактивных бактерий H. alvei, адаптировавшихся к условиям пивоваренного производства (табл. 5.24).
Признаки вида: H. alvei отличают от Enterobacter и Serratia по реакциям, приведенным в табл. 5.19. Реакции, позволяющие отличать И. alvei от отдельных видов этих родов, указаны в табл. 5.2.
Таблица 5.19. Дифференцирующие признаки рода Hafnia и биохимически сходных родов1
Род Klebsiella
Прямые палочки, 0,3–1,0 х 0,6–6,0 мкм, одиночные, в парах или коротких цепочках. Клетки имеют капсулу. Грамотрицательные. Неподвижные. Факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура 37°С. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты и газа, однако встречаются и не образующие газ штаммы. Оксидазоотрицательные; катал азоположительные. По образованию индола, пробе с метиловым красным, реакции Фогеса-Проскауэра, а также способности расти на среде Симмонса с цитратом виды варьируют. Как правило, по лизивдекарбоксилазе положительные, по орнитиндекарбоксилазе и аргининдигидролазе отрицательные. Некоторые виды гидролизуют мочевину. Способны расти в присутствии KCN. H2S не образуют. Восстанавливают нитрат. Большинство видов сбраживает все обычно тестируемые углеводы, за исключением дульцитолз и эритритола. Встречаются в фекалиях человека и клиническом материале, почве, воде, зерне, на фруктах и овощах. К. pneumoniae, К. oxytoca и иногда другие виды вызывают оппортунистические инфекции у человека, в том числе бактериемию, пневмонию, инфекции мочевых путей. Часто вызывают внутрибольничные инфекции у новорожденных, урологических и гериатрических больных, а также у больных, находящихся в отделениях интенсивной терапии.
Типовой вид: Klebsiella pneumoniae.
Для К. pneumonia и К. oxytoca характерна массивная полисахарид ная капсула, определяющая образование крупных мукоидных колоний, особенно на средах, богатых углеводами. Этот признак отличает их от других представителей Enterobacteriaceae, за исключением некоторых штаммов Enterobacter aerogenes и Escherichia coli. Известно более 80 капсульных (К) антигенов, используемых для серотипирования, клебсиелл.
Признаки, отличающие виды Klebsiella от видов Enterobacter, приведены в табл. 5.10 и 5.12. Дополнительные признаки указаны в табл. 5.11, Наиболее трудно отличать штаммы К. pneumoniae от штаммов неподвижных представителей Enterobacter aerogenes, которые очень медленно разжижают желатину. Решающее значение в этих случаях может иметь тест на уреазу, (Бактерии К pneumoniae уреазоположительные.)
Дифференциация видов рода Klebsiella. Реакции, позволяющие дифференцировать виды и подвиды Klebsiella, приведены в табл. 5.20. Полный перечень тестов содержится в табл. 5.2.
Таблица 5.20. Дифференциация видов и подвидов рода Klebsiella1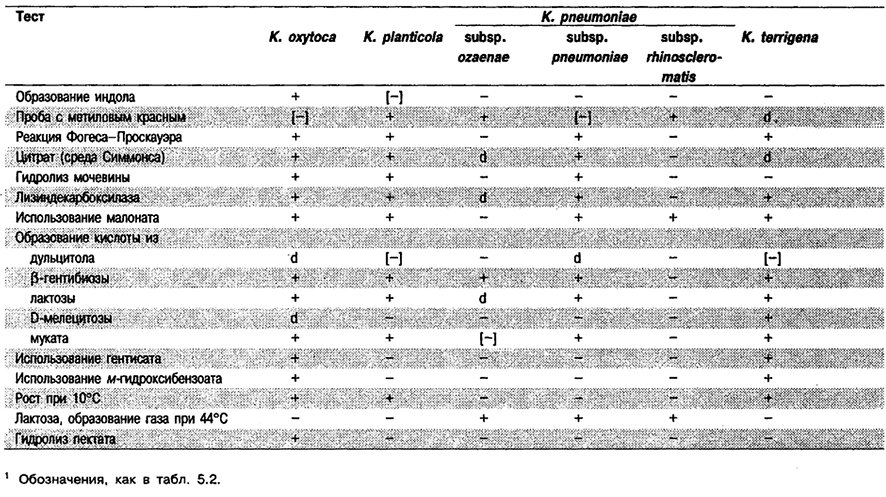
От редакции: Виды Klebsiella ornithinofytica и Klebsiella terrigena не приведены в «Sergey's Manual of Systematic Bacteriology». Вид К. ornithinolytica описан в 1989 г. (Sakazaki et al., Сигт. Microbiol. 18: 201-206; lilt J. Syst. Bacterid. 39: 495-497, 1989) для штаммов, ранее отнесенных к орнитанположительному виду К oxytoca, NIH (Japan) Group 12 (Sakazaki et al., Curr. Microbiol. 18: 201–206, 1989), CDC Klebsiella Group 47 (индолположительные, орнитинположительные) и индолположительной, орнитинположительной биогруппе К planticola (Farmer et al., J. Clin Microbiol. 21: 46–76, 1985). Между двумя указанными лабораториями нет разногласий в части фенотипических свойств этих организмов, но у них отсутствует единое мнение о том, указывают ли данные о родстве ДНК, что этот вид следует оставить как биогруппу К. planticola или же согласно им нужно считать его отдельным видом. К. terrigena описан в 1981 г. (Izard et al.. Int. J. Syst. Bacterid. 31: 116– 127). Существовавшие ранее виды Klebsiella ozaenае и Klebsiella rhinoscteromatis в 1984 г. было предложено рассматривать как подвиды Klebsiella pneumoniae (OrskoVj In: «Bergey's Manual of Systematic Bacteriology» vol. 1, p. 461–465; Int. J, Syst. Bacteriol. 34: 355–357, 1984). He вызывает сомнений, что К ozaenae и К rhinoscleromatis относятся к тому же геновиду, что и К. pneumoniae, однако большинство лабораторий все еще рассматривает и называет их в публикациях как отдельные виды.
Род Kluyvera
Прямые палочки, 0,5–0,7 х 2–3 мкм, по Граму окрашиваются отрицательно. Подвижные за счет редких перитрихиальных жгутиков. Факультативные анаэробы; хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура 37°С. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты и газа. Оксидаз о отри дательные; каталазоположительные; индол образуют; проба с метиловым красным и, как правило, тест на среде Симмонса с цитратом положительные; реакция Фогеса-Проскауэра отрицательная; по орнитиндекарбоксилазе положительные; по аргининдигидролазе отрицательные. Используют малонат и растут в присутствии KCN. H2S не образуют; мочевину не гидролизуют. Восстанавливают нитрат. Сбраживают большинство углеводов, но обычно не сбраживают многоатомные спирты. Встречаются в пищевых продуктах, почве, сточных водах и клиническом материале (наиболее часто присутствуют в дыхательных путях, мочевых путях и фекалиях, иногда в крови). В отдельных случаях вызывают оппортунистические инфекции.
Типовой вид: Kluyvera ascorbata.
Виды Kluyvera наиболее сходны с видом Buttiauxella agrestis (у человека не выделен) и в некоторой степени с Е. coli, видами Citrobacter и Enterobacter. Тесты, позволяющие отличить представителей Kluyvera от этих организмов, приведены в табл. 5.3 и 5.2.
Дифференциация видов рода Kluyvera: См, табл. 5.21. Дополнительно следует отметить, что для К cryocrescens обычно характерны обширные зоны подавления роста под действием карбенициллина и цефалотина, а для К. ascorbata – зоны гораздо меньших размеров.
Таблица 5.21. Дифференциация видов рода Kluyvera1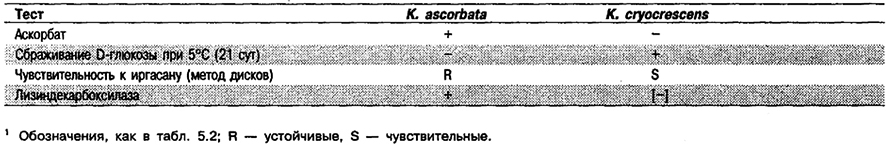
Род Leclercia
От редакции: Род Leclercia не приведен в «Вег- gey's Manual of Systematic Bacteriology». Этот род описан в 1986 г. (Tamura et al, Curr Microbiol. 13: 179–184: Int. J. Syst. Bacteriol. 37: 179– 1,80, 1987) и включает один вид L. adecarboxylata.
Прямые палочки; по Граму окрашиваются отрицательно; подвижные за счет перитрихиальных жгутиков. Факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура 37°С. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты и газа. Оксидазоотрицательные; каталазоположительные; индол не образуют; проба с метиловым красным и реакция Фогеса-Проскауэра отрицательные; на среде Симмонса с цитратом не растут. По лизин- и орнитиндекарбоксилазе и аргининдигидролазе отрицательные. Используют малонат; способны расти в присутствии KCN. H2S и уреазу не образуют. Некоторые штаммы продуцируют желтый пигмент. Восстанавливают нитрат. Сбраживают большинство углеводов, за исключением мио-инозитола и D-сорбитола. Встречаются в клиническом материале, чаще всего в мокроте и крови, а также в пищевых продуктах, воде и других природных источниках. В отдельных случаях вызывают оппортунистические инфекции у человека.
Типовой (и единственный) вид: Leclercia adecar- boxyiata.
От редакции: Ранее L. adecarboxylata был известен как Escherichia adecarboxylata и рассматривался многими как синоним Enterobacter agglomerans. Вид включен в «Bergey's Manual of Systematic Bacteriology» как species insertae sedis внутри рода Escherichia.
Признаки вида: Leclercia следует отличать от других родов, которые дают отрицательные реакции в тестах на лизин- и орнитиндекарбоксилазу и аргининдигидролазу (табл. 5.2). От лизин-, аргинин- и орнитинотрицательных штаммов Е. coli Leclercia отличают с помощью тестов на рост в присутствии KCN, использование малоната и сбраживание D-адонитола, D-сорбитола и целло- биозы (табл. 5.18).
Род Leminorella
От редакции: Род Leminorella не приведен в «Вегgey's Manual of Systematic Bacteriology». Этот род описан в 1985 г. (Hickman-Brenner et al., J. Clin. Microbiol. 21: 234–239; Int. J. Syst. Bacteriol, 35: 375–376, 1985) и включает два вида: L. grimontii и L, richardii,
Прямые палочки. Грамотрицательные. Неподвижные. Факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура 37°С. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты. Образование газа из D-глюкозы у одних штаммов отсутствует, у других варьирует или происходит с задержкой. Оксидазоотрицательные; каталазоположительные. Индолотрицательные; реакция Фогеса-Проскауэра отрицательная; проба с метиловым красным и тест на среде Симмонса с цитратом у одного вида положительные, а у другого отрицательные. По лизин- и орнитиндекарбоксилазе и аргининдигидролазе отрицательные. Образуют H2S. По уреазе, устойчивости к KCN и использованию малоната отрицательные. Восстанавливают нитрат. Сбраживают L-арабинозу и D-ксилозу; почти все остальные обычно тестируемые углеводы не сбраживают. Источники выделения – кал и моча человека. Медицинское значение неизвестно.
Типовой вид; Leminorella grimontii.
Подобно видам Proteus виды Leminorella образуют H2S, просветляют среду с тирозином и не сбраживают D-маннозу, однако в отличие от Proteus сбраживают L-арабинозу, не гидролизуют мочевину и не обладают фенилаланиндезаминазной активностью. Тесты, позволяющие отличить Leminorella от Proteus и Providencia, а также от Budvicia, приведены в табл. 5.22, от Pragia – в табл. 5.26, от других лизин-, аргинин- и орнитинотрицательных видов – в табл. 5.2.
Дифференциация видов рода Leminorella: См. табл. 5.23. Другие биохимические признаки перечислены в табл. 5.2.
Таблица 5.22. Отличия LeminoreUa от Budvicia, Proteus и Providentia1
Таблица 5.23. Дифференциация видоа рода Leminorella1
Род Moellerella
От редакции: Род Moellerella не приведен в «Вегgey's Manual of Systematic Bacteriology». Это род описан в 1984 г. (Hickman-Brenner et al., J. Clin. Microbiol. 19: 460–463; int. J. Syst. Bacteriol. 34: 355–357, 1984) и включает один вид М. wiscon- sensis.
Прямые палочки. Грамотрицательные. Неподвижные; факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура 37°С. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты, но не газа. Оксидазоотрицательные, каталазоположительные. Индолотрицательные; реакция Фогеса-Проскауэра отрицательная; проба с метиловым красным положительная; способны расти на агаре Симмонса с цитратом. Активностями лизин- и орнитиндекарбокснлазы и аргининдигидролазы не обладают. H2S не образуют; мочевину не гндролизуют; малонат не используют. Большинство штаммов растет в присутствии KCN. Восстанавливают нитрат. Сбраживаемые углеводы включают адонитол, D-арабитол, D-галактозу, лактозу, D-маннозу, мелибиозу, рафинозу и сахарозу. Глицерол и D-маннитол сбраживают за 7 сут. Встречаются в кале человека, а также в воде. Ассоциированы с желудочно-кишечными заболеваниями у человека, но как возбудители не определены.
Типовой (и единственный) вид: Moellerella wisconsensis.
Признаки вида: Следует отличать Moellerella от других родов, отрицательных по лизин- и орнитнндекарбоксилазе и аргининдигидролазе, в том числе от биогруппы штаммов Е. coli, отрицательных по указанным признакам. Для определения этих признаков используют тесты на индол, рост на среде Симмонса с цитратом, подвижность, образование газа из D-глюкозы и сбраживание ряда углеводов (табл. 5.2).
Род Morganella
Прямые палочки, 0,6–0,7 х 1,0–1,7 мкм. Грамотрицательные. Подвижные за счет перитрихиальных жгутиков. Роение отсутствует. Факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура 37°С. Из обычно тестируемых углеводов катаболизируют с образованием кислоты и обычно газа (иногда с задержкой) только D-глюкозу и D-маннозу. Оксидазоотрицательные; каталазоположительные; индолположмтельиые; проба с метиловым красным положительная; реакция Фогеса-Проскауэра отрицательная; на агаре Симмонса с цитратом не растут; по лизиндекарбокснлазе и аргининдигидролазе отрицательные; по орнитнндекарбоксилазе положительные. Осуществляют окислительное дезаминириванне фенилаланина и триптофана. Гидролизуют мочевину. Способны к росту в присутствии KCN. H2S не образуют. Разлагают тирозин, вызывая просветление среды, содержащей эту нерастворимую аминокислоту. Восстанавливают нитрат. Встречаются в фекалиях человека, собак, других млекопитающих и рептилий. Служат возбудителями вторичных оппортунистических инфекций, выделяемыми при бактериемии, инфекциях дыхательных путей, ран и мочевых путей.
Типовой (и единственный) вид: Morganella тогganii.
Признаки вида: Признаки, отличающие Morganella от Proteus и Providencia, приведены в табл. 5.27.
Род Obesumbacterium
Плеоморфные палочки, 0,8–2,0 х 1,5–100 мкм; при росте в пивном сусле в присутствии живых дрожжей преобладают короткие, «толстые» палочки, при росте в бактериологических средах – длинные плеоморфные палочки. Грамотрицательные. Неподвижные; факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура ~32°С. Катаболизируют D-глюкозу и несколько других углеводов с образованием кислоты. Оксидазоотрицательные; каталазоположительные, индолотрицательные; реакция Фогеса-Проскауэра отрицательная; проба с метиловым красным у большинства штаммов отрицательная; на среде Симмонса с цитратом не растут. По лизин- и орнитиндекарбоксилазе положительные; по аргининдигидролазе отрицательные. H2S не образуют; мочевину не гидролизуют; в присутствии KCN не растут; малонат не используют. Восстанавливают нитрат. Большинство штаммов сбраживает D-маннозу и трегалозу, но редко какие-либо другие углеводы. Во многих биохимических тестах, обычно используемых для дифференциации Enterobacteriaceae, реакция отрицательная или с задержкой. Растут медленно, образуя за 24 ч колонии диаметром менее 0,5 мм. Встречаются как примесь в пиве; могут сохранять жизнеспособность и расти в присутствии живых дрожжей в процессе производства пива.
Типовой (и единственный) вид: Obesumbacterium proteus.
Признаки вида: Биохимические тесты, позволяющие отличить Obesumbacterium от других родов, приведены в табл. 5.24.
От редакции; Внутри вида О. proteus различают две биогруппы. В пивоваренной промышленности для обеих принято название «Haftiia protea», которое незаконно. Биогруппа 2 представлена О. proteus sensu stricto. Био группа 1 – это на самом деле биогруппа необычных (метаболически неактивных) бактерий Hafnia alvei, адаптировавшихся к условиям пивоваренного производства (табл. 5.24).
Таблица 5.24. Биохимическая дифференциация Hafnia atvei, Obesumbacterium proteus и Escherichia blattae1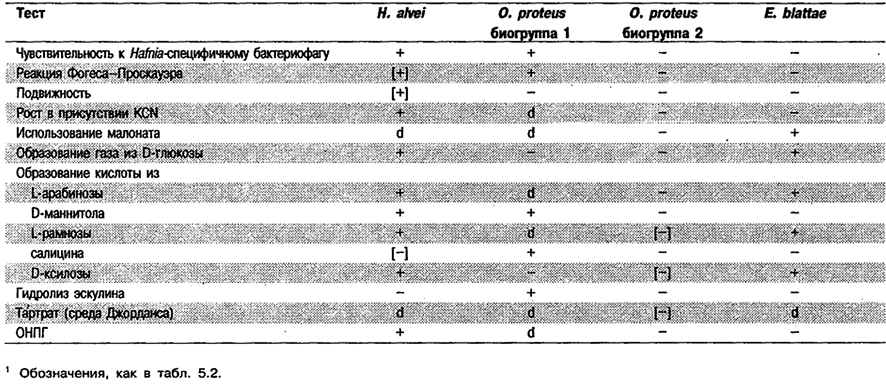
Род Pantoea
От редакции: Род Pantoea не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1989 г. (Gavini et al., Int. J. Syst. Bacteriol. 39: 337–345) и включает два вида; P. agglomerans (синонимы: Enterobacter agglomeransErwinia herbicola, Erwinia milletiae) и P. dispersa.
Прямые палочки, 0,5–1,0 x 1–3 мкм. Грамотрицательные. Подвижные за счет перитрихнальных жгутиков. Большинство штаммов образует желтый пигмент. Факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура 30°С, Катаболизируют D-глюкозу и другие углеводы с образованием кислоты, но не газа. Оксидазоотрицательные; каталазоположительные; индолотрицательные; реакция Фогеса-Проскауэра положительная; способны расти на среде Снммонса с цитратом; проба с метиловым красным варьирует. По лизин- и орнитиндекарбоксилазе и аргининдигидролазе отрицательные (хотя, согласно данным более ранних исследований, по орнитиндекарбоксилазе все штаммы отрицательные, позднее установлено, что 30% штаммов Р. agglomerans обладают активностью орнитиндекарбоксилазы, см. Gavini et al.); H2S не образуют; мочевину не гццролизуют. Большинство штаммов растет в присутствии KCN. По способности использовать малонат виды различаются. Восстанавливают нитрат. В число углеводов, сбраживаемых всеми или большинством штаммов, входят L-арабиноза, D-галактоза, D-ксилоза, мальтоза, D-маннитол, D-манноза, L-рамноза, сахароза и трегалоза, Выделены с поверхности растений, семян, из почвы и воды, а также из ран, крови и мочи человека и животных. Вызывают оппортунистические инфекции у человека.
Типовой вид: Pantoea agglomerans.
Дифференциация видов рода Pantoea: См. табл. 5.25.
Таблица 5.25. Дифференциация видов рода Pantoea1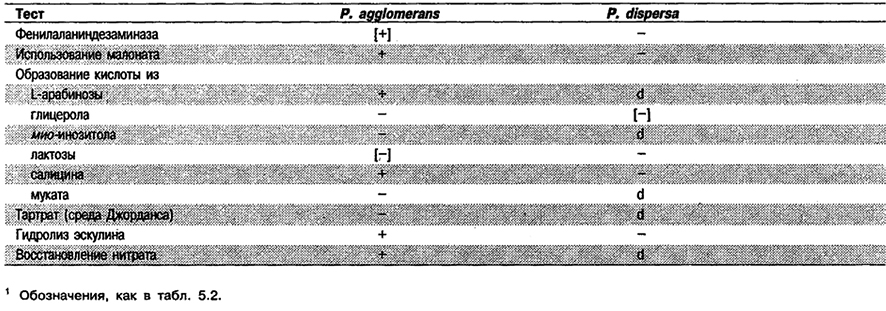
Признаки, отличающие виды Pantoea от представителей других родов, приведены в табл. 5.2.
От редакции: Для вида P. agglomerans часто используют синонимические названия, в основном Enterobacter agglomerans. В настоящем определителе данный вид включен и в род Enterobacter, и в род Pantoea.
Род Pragia
От редакции: Род Pragia не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1988 г. (Aldova et al., Int. J. Syst. Bacteriol. 38: 183–189) и включает один вид A fontium.
Прямые палочки. Грамотрицательные. Подвижные за счет перитрихнальных жгутиков. Факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура 37°С. Катаболизируют D-глюкозу и немногие другие углеводы с образованием кислоты, но не газа. Оксидазоотрицательные; каталазоположительные; индолотрицательные; реакция Фогеса-Проскауэра отрицательная; проба с метиловым красным положительная; как правило, способны расти на агаре Снммонса с цитратом. По лизин- и орнитиндекарбоксилазе и аргининдигидролазе отрицательные. Образуют H2S. Мочевину не гидролизуют; способны расти в присутствии KCN; используют малонат. Восстанавливают нитрат. Сбраживают D-галактозу; 50–75% штаммов сбраживают глицерол (с задержкой), мио-инозитол (с задержкой) и салицин; другие обычно тестируемые углеводы не сбраживают. Встречаются в питьевой воде; один штамм выделен из кала человека. Данных о патогенности нет.
Типовой (и единственный) вид: Pragia fontium.
Признаки вида: В табл. 5.26 приведены признаки, по которым Pragia отличают от других лизин-, аргинин- и орнитинотрицательных родов. Полный перечень биохимических тестов дан в табл. 5.2.
Таблица 5.26. Отличия Pragia от Budvicia и Leminorella1 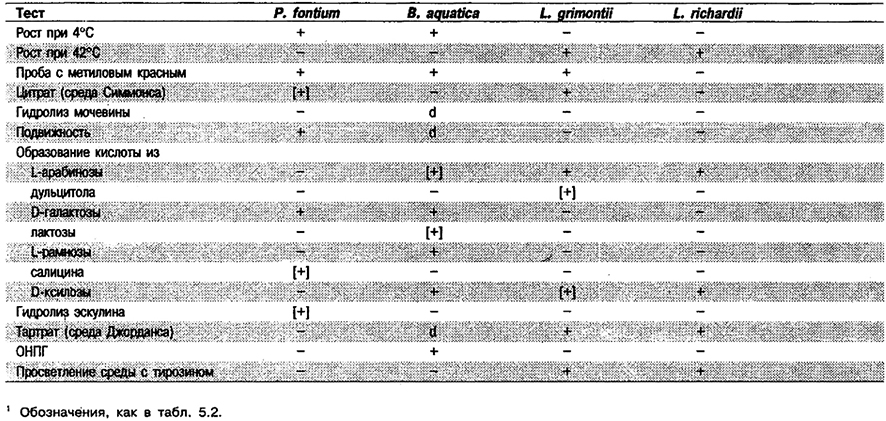
Род Proteus
Прямые палочки, 0,4–0,8 х 1–3 мкм. Грамотрицательные. Подвижные за счет перитрихнальных жгутиков. У большинства штаммов имеет место роение с периодическими циклами миграции, приводящей к образованию концентрических зон или распространению в виде однородной пленки по влажной поверхности: питательной среды, отвержденной желатиной или агаром, Факультативные анаэробы; хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура 37°С. Катаболизируют D-глюкозу и немногие другие углеводы с образованием кислоты и обычно газа. Оксидазоотрицательные; каталазоположительные; реакция с метиловым красным положительная. По образованию индола, реакции Фогеса-Проскауэра и пробе на среде Симмонса с цитратом виды различаются. Лизиндекарбоксилаза и аргининдигидролаза отсутствуют. Декарбоксилировать орнитин способен только P. mirabilis. Осуществляют окислительное дезаминирование фенилаланнна и триптофана; гидролизуют мочевину. Разлагают тирозин» вызывая просветление агаризованиой среды, содержащей нерастворимую аминокислоту. Растут в присутствии KCN. Обычно образуют H2S. Малой ат не используют.
Восстанавливают нитрат. Один или более видов сбраживают глицерол, D-ксилозу, мальтозу, сахарозу и трегалозу. Встречаются в кишечнике у человека и разнообразных животных, а также в навозе, почве и загрязненных водах. P. myxofaciens выделен только из личинки шелкопряда непарного. Патогенны для человека: вызывают инфекции мочевых путей, а также вторичные поражения, приводящие к образованию септических очагов, часто при ожогах.
Типовой вид: Proteus vulgaris.
Отличить Proteus, Providencia и Morganella от других представителей Enterobacteriaceae можно по признаку наличия фенилаланиндезаминазы (или триптофандезаминазы). Признаки, позволяющие отличить Proteus от Providencia и Morganella, указаны в табл. 5.27.
Таблица 5.27. Биохимическая дифференциация Proteus, Providencia и Morganella morganii1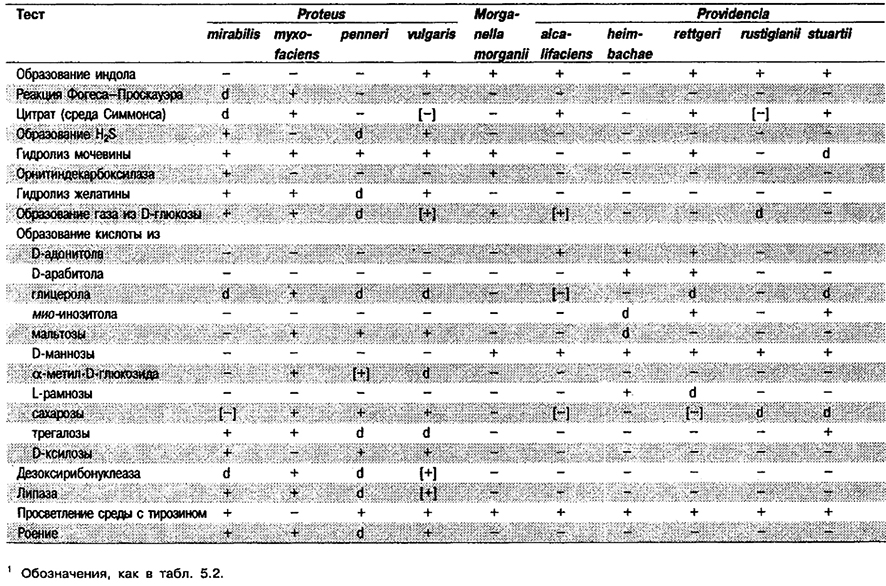
Дифференциация видов рода Proteus: См. табл. 5.28. Дополнительные биохимические характеристики приведены в табл. 5.2.
Таблица 5.28. Дифференцирующие признаки видов рода Proteus1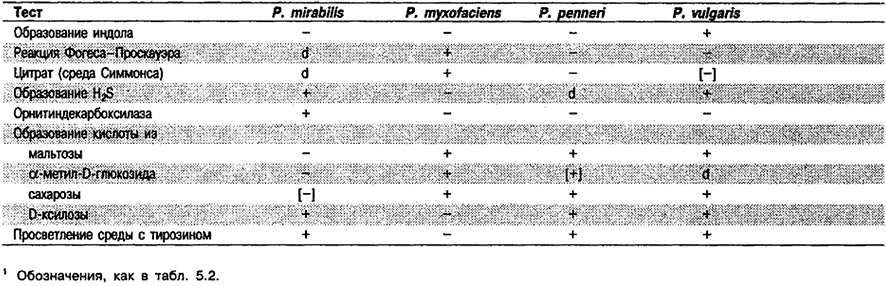
От редакции: Вид Proteus penneri не приведен в «Bergey's Manual of Systematic Bacteriology». Этот вид описан в 1982 г. (Hichman et ah, J. Clin. Microbiol. 15: 1097–1102; Int. J. Syst. bacteriol. 33: 438-440, 1983).
Род Providencia
Прямые палочки, 0,6–0,8 x 1,5–2,5 мкм. Грамотрицательные. Подвижные за счет перитрихиальных жгутиков. Роение отсутствует. Факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура 37°С. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты; некоторые штаммы образуют газ. Оксидазоотрицательные; каталазоположительные; индолположителыше (за исключением P. heimbachae); проба с метиловым красным обычно положительная; реакция Фогеса-Проскауэра отрицательная. Некоторые виды способны расти на среде Симмонса с цитратом. По лизин- и орнитиндекарбоксилазе и аргининдигидролазе отрицательные. Осуществляют окислительное дезаминирование триптофана. Разлагают тирозин, вызывая просветление агаризованиой среды, содержащей эту нерастворимую аминокислоту. H2S не образуют; мочевину не гндролизуют (исключение составляет P. rettgeri). Способны расти в присутствии KCN (за исключением Р. heimbachae). Малонат не используют. Сбраживают маннозу, а также один или несколько из следующих многоатомных спиртов: адонитол, D-арабитол, эритритол, мио-инозитол, D-маннитол. Выделены при желудочно-кишечных расстройствах из кала, при инфекциях мочевых путей, из ран и ожогов, при бактериемии, а также из пингвинов. Патогенны для человека.
Типовой вид: Providencia alcalifaciens.
Признаки, отличающие виды Providencia от Proteus и Morganella, перечислены в табл. 5.27.
Дифференциация видов рода Providencia: См. табл. 5.29. Другие признаки этих видов приведены в табл. 5.2.
Таблица 5.29. Дифференциация видов рода Providencia 1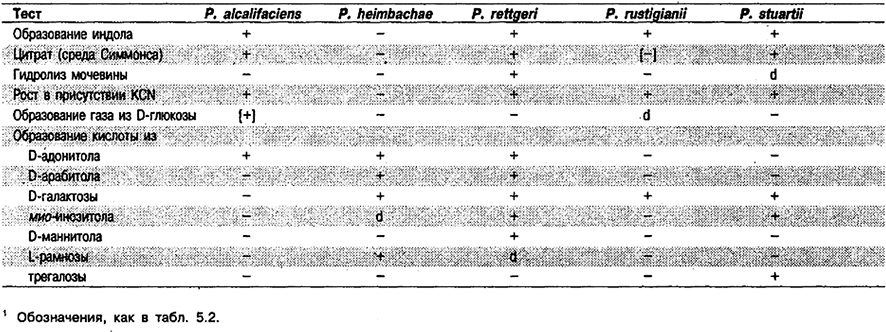
От редакции: Providencia heimbachae и Providencia rustigianii не приведены в «Bergey's Manual of Systematic Bacteriology». Вид Providencia heimbachae описан в 1986 г. (Muller et ah, Int. J. Syst. bacteriol. 36: 252–256), a Providencia rustigianii (ранее известный как биогруппа 3 Providencia alcalifaciens) в 1983 г. (Hickman-Brenner et al., J. Clin. Microbiol. 17: 1057–1060: Int. J. Syst. Bacteriol. 33: 672-674, 1983).
Род Rahnella
Мелкие палочки, 0,5–0,7 х 2–3 мкм. Грамотрицательные. Неподвижные при З6°С и подвижные за счет перитрихиальных жгутиков в условиях роста при 25°С. Факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Хорошо растут в диапазоне температуры 25–37°С. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты и (большинство штаммов) газа. Оксидазоотрицательные; каталазоположительные; индолотрицательные. У большинства штаммов проба с метиловым красным положительная. Реакция Фогеса-Проскауэра положительная; способны расти на среде Симмонса с цитратом; по лизин- и орнитиндекарбоксилазе и аргининдигидролазе отрицательные. H2S не образуют; мочевину не гидролизуют; используют малонат. В присутствии KCN не растут. Слабо положительные по фенилаланиндезаминазе. Восстанавливают нитрат. В число сбраживаемых углеводов входят D-арабиноза, D-ксилоза, дульцитол, лактоза, мальтоза, D-маннитол, D-манноза, мелибиоза, рафиноза, L-рамноза, салицин, D-сорбитол, сахароза, трегалоэа и целлобиоза. Встречаются в пресной воде. Иногда источником вьщеления служит клинический материал; медицинское значение неизвестно.
Типовой (и единственный) вид: Rahnella aquatilis.
Признаки вида: Признаки, отличающие Rahnella от других лизин-, аргинин- и орнитинотрицательных родов, приведены в табл. 5.2.
Род Salmonella
Прямые палочки, 0,7–1,5 х 2–5 мкм. Грамотрицательные. Обычно подвижные за счет перитрихиальных жгутиков. Факультативные анаэробы. Хемоорганотрофы; обладают и дыхательным и бродильным типами метаболизма. Оптимальная температура 37°С. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты и обычно газа. Оксидазоотрицательные; каталазоположительные; индолотрицательные; реакция Фогеса-Проскауэра отрицательная, проба с метиловым красным положительная. Способны расти на среде Симмонса с цитратом. По лизин- и орнитиндекарбоксилазе положительные; по аргининдигидролазе различаются. Образуют H2S; мочевину не гидролизуют; по способности к росту в присутствии KCN и использованию малоната различаются. Восстанавливают нитрат. Как правило, сбраживаемые углеводы включают L-арабинозу, D-ксилозу, мальтозу, D-маннитол, D-маннозу, L-рамнозу, D-сорбитол и трегалозу. Встречаются у человека, гомойотермных и пойкилотермных животных, в пищевых продуктах и природной среде. Патогенны для человека и многих видов животных. Вызывают брюшной тиф, кишечные инфекции, гастроэнтерит и септицемию.
Типовой вид: Salmonella choleraesuis.
Дифференциация видов рода Salmonella: Большинство сальмонелл – газообразующие бактерии, однако Salmonella choleraesuis серо вар typhi (Salmonella typhi; Salmonella серовар typhi) (важное исключение) и представители некоторых других сероваров газ никогда не образуют. Признаки, позволяющие отличить род Salmonella от других представителей Enterobacteriaceae, приведены в табл. 5.30.
Таблица 5.30. Отличия рода Salmonella от других родов1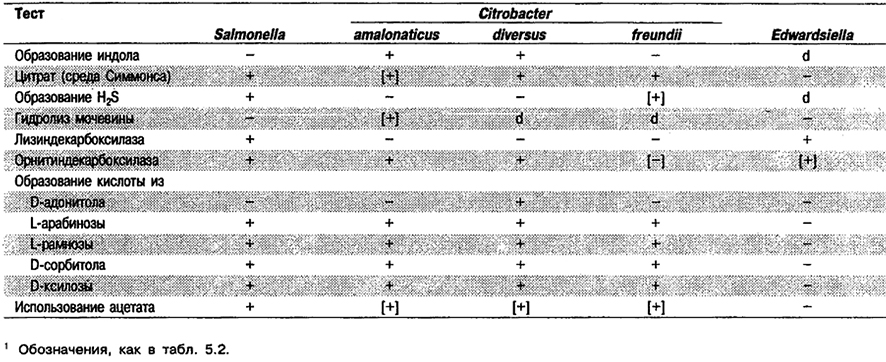
Деление на серовары: В схеме Кауфмана-Уайта серовары обозначены цифрами и буквами, означающими различные антигены – соматические (О), капсульные (Vi) и жгутиковые (Н). Антигенная формула (например 6,7:r: 1,7) несет следующую информацию: О-антигены (6,7); Н-антигены 1-й фазы (r) и Н-антигены 2-й фазы (1,7) соответственно. Сальмонеллы с конкретным О-антигеном собраны в одну О-группу и расположены внутри группы в алфавитном порядке по Н-антигенам. Схема служит для идентификации как видов Salmonella, так и всех подвидов (см. ниже). В свою очередь серовары могут быть подразделены на основании результатов биохимических тестов, что может представлять интерес с эпидемиологической точки зрения (например, выделены ксилозоположительные и ксилозоотрицателъные штаммы Salmonella серовар typhi).
Фаготипирование: Фаготипирование проводят для эпидемиологического контроля за определенными сероварами Salmonella в основном за сероварами typhi и enteritidis. Для сальмонелл высокоспецифичен бактериофаг 01, который получили Феликс (Felix) и Кэллоу (Callow); он лизирует более 98% штаммов, идентифицированных путем рутинного определения Salmonella (Kallings, Acta Pathol. Microbiol. Scand. 70: 446-454, 1967).
Дифференциация видов и подвидов рода Salmonella: Признаки, позволяющие дифференцировать виды и подвиды Salmonella, приведены в табл. 5.31. Полный перечень биохимических тестов представлен в табл. 5.2.
Таблица 5.31. Дифференциация видов, подвидов и некоторых сероваров рода Salmonella1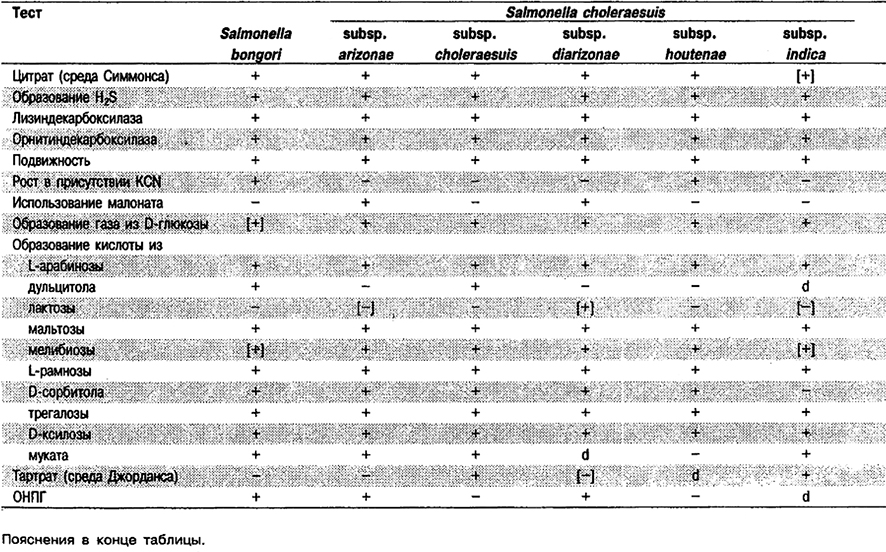
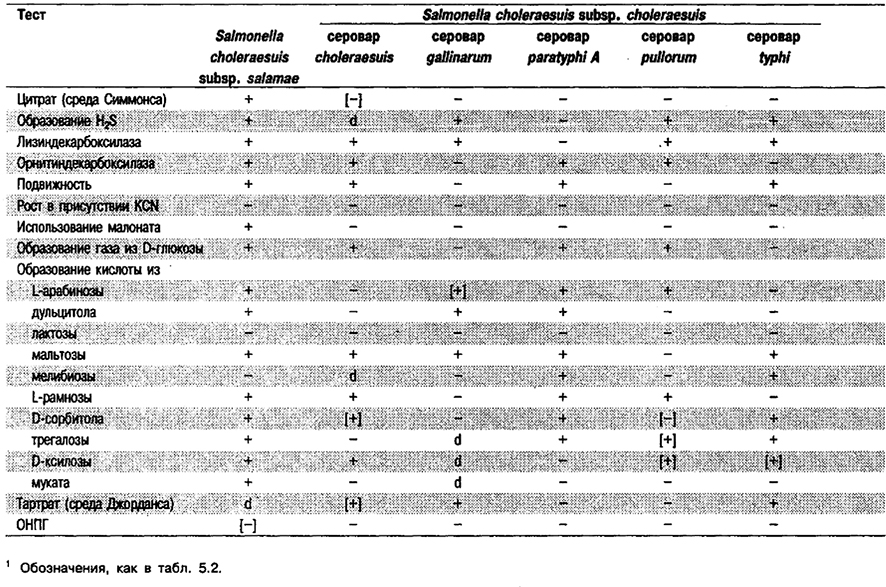
От редакции: Salmonella bongori и подвиды Salmonella choleraesuis не приведены в «Bergey's Manual of Systematic Bacteriology». Вид Salmonella bongori (прежнее название Salmonella choleraesuis subsp. bongori) описан в 1989 г. (Reeves et al., J. Clin Microbiol. 27: 313–320; Int. J. Syst. Bacteriol. 39: 371, 1989). Подвиды Salmonella choleraesuis subsp. arizonae, choleraesuis, diarizonae, houtenae и salamae описаны в 1982 г. (Le Minor et al., Ann. Microbiol. 133 B: 245–254: Int. J. Syst. Bacteriol, 35: 375–376, 1985), Подвид Salmonella choleraesuis subsp. indica описан в 1986 г. (Le Minor et al., Ann. Inst. Pasteur. 137 B: 211–217; Int. J. Syst. Bacteriol 37: 179-180, 1987).
Номенклатура: Все серовары Salmonella (включая «Arizona») относятся к двум видам. Salmonella bongori содержит менее 10 очень редких сероваров. Все остальные 2 500 сероваров выделены внутри вида Salmonella choleraesuis, который может быть по фенотипическим и генетическим критериям разделен на 6 подвидов. Все серовары внутри подвида choleraesitis имеют названия, тогда как в других подвидах серовары (за исключением некоторых в подвидах salamae и houtenae) не имеют названий. Диагностическим лабораториям рекомендовано для имеющих названия сероваров Salmonella использовать эти названия, а не имеющие названий серовары обозначать с помощью антигенной формулы и указанием подвида. Например: Salmonella typhi или Salmonella серовар typhi; typhimurium или Salmonella серовар typhi - murium; Salmonella subsp. arizonae серовар 50:z4, z24; (прежнее название – «Arizona hinshawii»), Salmonella subsp. salamae серовар 56:z10:e,n,x. Следует также отметить, что группа Arizona [ранее классифицированная в США как «Arizona hinshami», а в настоящее время подвид arizonae (однофазные штаммы) и подвид diarizonae (двухфазные штаммы)] была серотипирована в США с использованием схемы «Arizona», а в настоящее время ее серотипируют по схеме серотипирования Salmonella. В связи с этим (по крайней мере в США) следует приводить обе антигенные формулы: например, Salmonella серовар 50:z4, z24- (прежнее название – «Arizona hinshawi» 9a, 9b:1,3, 11:–). Сложные вопросы номенклатуры внутри Salmonella подробно рассмотрены в публикациях Ле Минор (Le Minor) с соавт. (Bergey's Manual of Systematic Bacteriology vol. 1, p. 427–448, 1984; Ann. Microbiol. 133 B: 223-243, 1982: Ann. Microbiol. 133 B: 245–254, 1982) и Фармер (Fanner) с соавт. (Clin. Microbiol. Newsl. 6: 63– 66, 1984).
Род Serratia
Прямые палочки, 0,5–0,8 x 0,9–2,0 мкм. Грамотрицательные. Обычно подвижные за счет перитрихиальных жгутиков. Факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Хорошо растут при 30–37°С. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты и часто газа. Индолотрицательные, за исключением некоторых штаммов S. odorifera. По результатам пробы с метиловым красным различаются. Способны расти на среде Симмонса с цитратом. Реакция Фогеса-Проскауэра обычно положительная, исключение составляет S. fonticola. Большинство штаммов положительные по лизнндекарбоксилазе, отрицательные по аргининдигидролазе и положительные по орнитнндекарбоксилазе. H2S не образуют; мочевину не гидролизуют; малонат, как правило, не используют. Большинство штаммов образуют ДНКазу и гндролизуют кукурузное масло. Как правило, гидролизуют желатину. Восстанавливают нитрат. Углеводы, сбраживаемые всеми или большинством штаммов, включают мальтозу, D-маннитол, D-маннозу, салицин, сахарозу и трегалозу. Встречаются в клиническом материале, почве, воде, на поверхности растений и в других природных источниках, а также в пищеварительном тракте грызунов и насекомых. S. marcescens вызывает оппортунистические инфекции у госпитализированных больных – септицемию и инфекции мочевых путей. Некоторые другие виды могут быть выделены при бактериемии либо из мокроты, но не имеют при этом клинического значения. Вызывают мастит у коров и другие инфекции у животных.
Типовой вид: Serratia marcescens.
Признаки, отличающие Serratia от биохимически сходных таксонов, приведены в табл. 5.32 и 5.33. Признаки, отличающие каждый вид Serratia от других таксонов, представлены в табл. 5.34.
Таблица 5.32. Дифференцирующие признаки рода Serratia и биохимически сходных таксонов1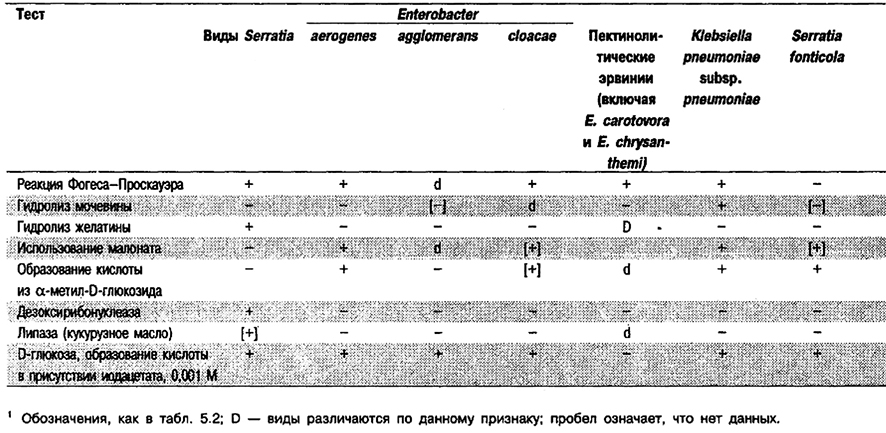
Таблица 5.33. Дополнительные тесты,
позволяющие отличить бактерии рода Serratia от биохимически сходных таксонов1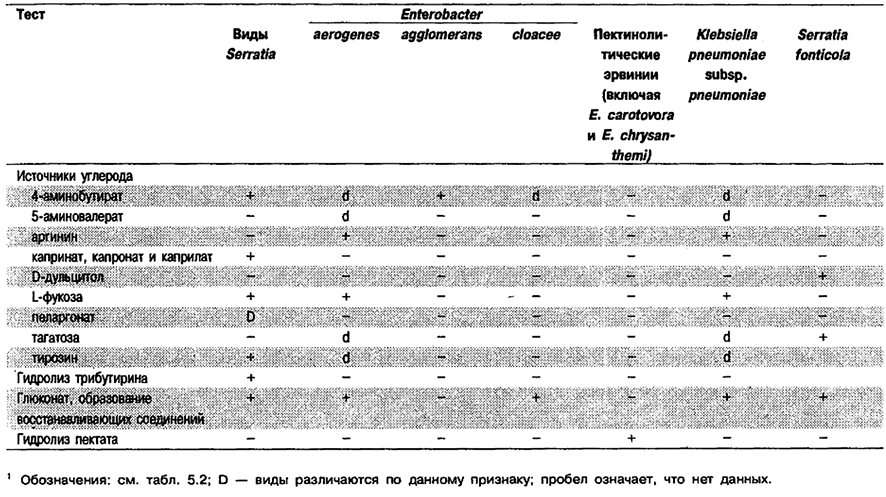
Таблица 5.34. Дифференциация видов рода Serratia1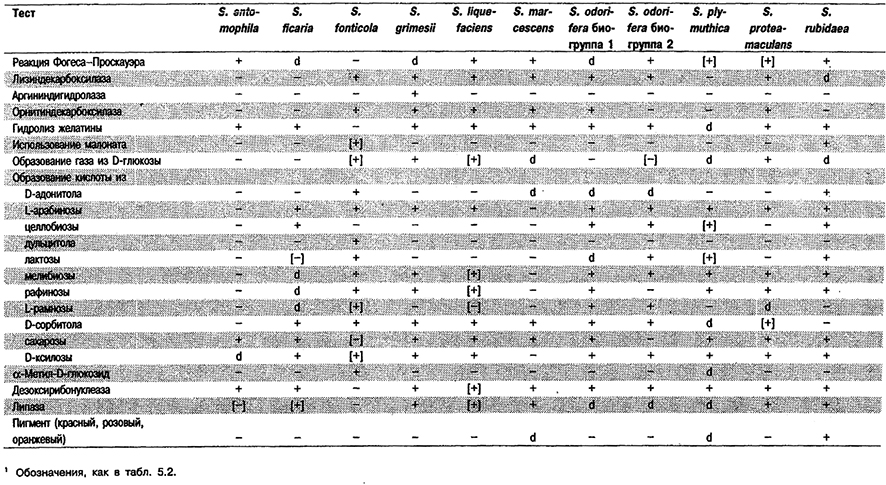
Дифференциация видов рода Serratia: См. табл. 5.35 и 5.36. Поскольку виды S. liquefaciens, S. grimesii и S. proteamaculans трудно различить с помощью стандартных биохимических тестов, в большинстве лабораторий соответствующие штаммы идентифицируют как S. liquefaciens или группа S. liquefaciens.
Таблица 5.35. Дифференциация видов рода Serratia – liquefaciens, proteamacutans и grimesii 1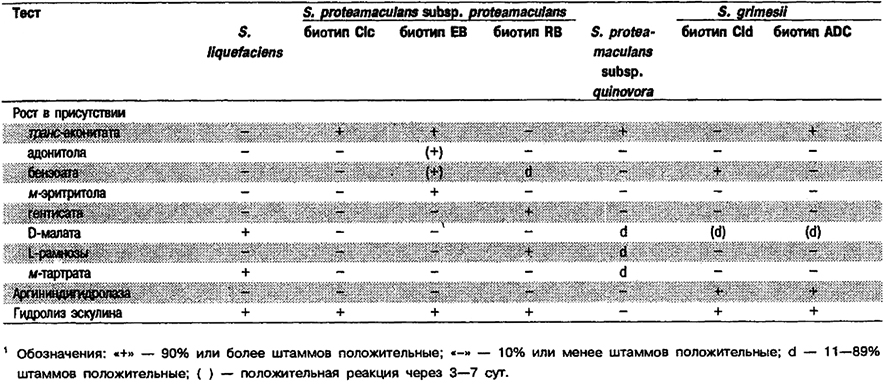
Таблица 5.36. Днфференциацив подвидов вида Serratia rubidaea1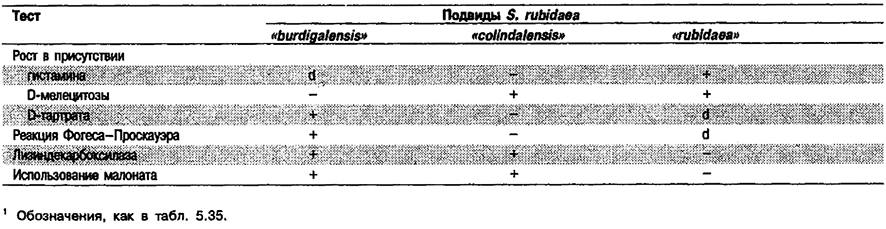
От редакции: Виды Serratia entomophila, Serratia grimesii, Serratia proteamaculans, Serratia proteamaculans subsp. proteamaculans, Serratia proteamaculans subsp. quinovora, «Serratia rubidaea subsp. burdigalen- sis», «Serratia rubidaea subsp. colindalensis», и «Serratia rubidaea subsp. rubidaea» либо упомянуты в «Bergey's Manual of Systematic Bacteriology» без их описания, либо не приведены в этом издании. Вид Serratia entomophila описан в 1988 г. (Grimont et al., Int. J. Syst. Bacteriol, 38: 1–6), Вид Serratia grimesii и вид Serratia proteamaculans, включающий два подвида – proteamaculans и quinovora, описаны в 1982 г. (Grimont et al., Curr, Microbiol. 7: 69–74; Int. J. Syst. Bacteriol. 33: 438–440, 1983). Подвиды burdigalensis, colindalensis и rubidaea внутри вида Serratia rubidaea описаны в 1990 г. (Grimont et al.; P. A. D. Grimont, личное сообщение).
Род Shigella
Прямые палочки. Грамотрицательные, Неподвижные. Факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура 37°С. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты; небольшое число штаммов образует газ. Оксидазоотрицательные; каталазоположительные. По образованию индола различаются. Проба с метиловым красным положительная; реакция Фогеса-Проскауэра отрицательная; на среде Симмонса с цитратом не растут; по лизиндекарбоксилазе и аргининдигидролазе отрицательные; по орнитиндекарбоксилазе различаются; H2S не образуют; мочевину не гидролизуют; малонат не используют, в присутствии KCN не растут. Восстанавливают нитрат. Сбраживаемые углеводы включают D-маннитол, D-маннозу и, как правило, мальтозу и трегалозу. Возбудители кишечных инфекций у человека и приматов, вызывающие бактериальную дизентерию.
Типовой вид; Shigella dysenteriae.
Часто бывает трудно отличить шигелл от неподвижных, лактозоотрицательных, не образующих газ штаммов и других неактивных штаммов Е. coli. Идентификация осложняется еще и тем, что некоторые штаммы Е. coli, как и Shigella, способны вызывать бактериальную дизентерию. Биохимические признаки, позволяющие отличить шигеллы от Е. coli, приведены в табл. 5.17.
Дифференциация видов рода Shigella: Род Shigella включает 4 вида: S, dysenteriae, S. flexneri, S. boydii и S. sonnei. Часто их определяют как. подгруппы А, В, С и D соответственно. Чтобы отличить от других шигелл вид S. sonnei, часто достаточно провести биохимические тесты (табл. 5.37); однако различить S. boydii, S. dysenteriae и S. flexneri по биохимическим признакам невозможно. Для надежной идентификации видов шигелл необходимо типирование по соматическим (О) антигенам.
Таблица 5.37. Биохимические отличия Shigella sonnei от других шигелл1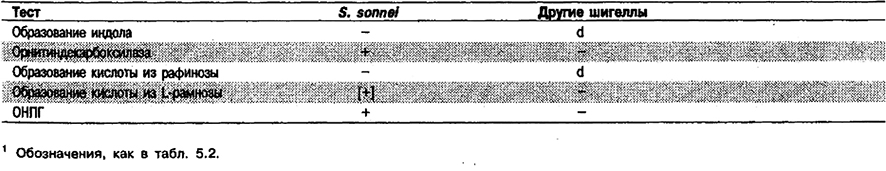
Род Tatumella
Мелкие палочки, 0,6–0,8 х 0,9–3 мкм. Грамотрицательные. Неподвижные при З6°С; более чем у половины штаммов клетки подвижные. Факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Хорошо растут при 25–36°С, но метаболически более активны при 25°С. Используют D-глюкозу и другие углеводы с образованием кислоты, но не газа. Оксидазоотрицательные; каталазоположительные. Отрицательные по следующим признакам: образование индола, проба с метиловым красным, реакция Фогеса-Проскауэра, рост на среде Снммонса с цитратом, лизин- и орнитиндекарбоксилаза, аргининдигидролаза, образование H2S, гидролиз мочевины, рост в присутствии KCN, использование малоната. По фенил аланиндезаминазе положительные. Восстанавливают нитрат. Сбраживаемые углеводы включают D-маннозу, сахарозу и трегалозу. Источниками выделения служат клинический материал (в основном из дыхательных путей), иногда кровь, а также животные. Вероятно, в отдельных редких случаях вызывают оппортунистические инфекции у человека.
Типовой (и единственный) вид: Tatumella ptyseos.
Признаки вида: Этот вид легко отличить от других представителей Enterobacteriaceae по признаку того, что он метаболически относительно неактивен при 36°С. Следует отличать данный вид от видов, положительных по фенилаланиндезаминазе, как указано в табл. 5.38. Полный перечень биохимических тестов приведен в табл. 5.2.
Таблица 5.38. Отличия Tatumella от других видов и родов, положительных по фенилаланиндезаминазе1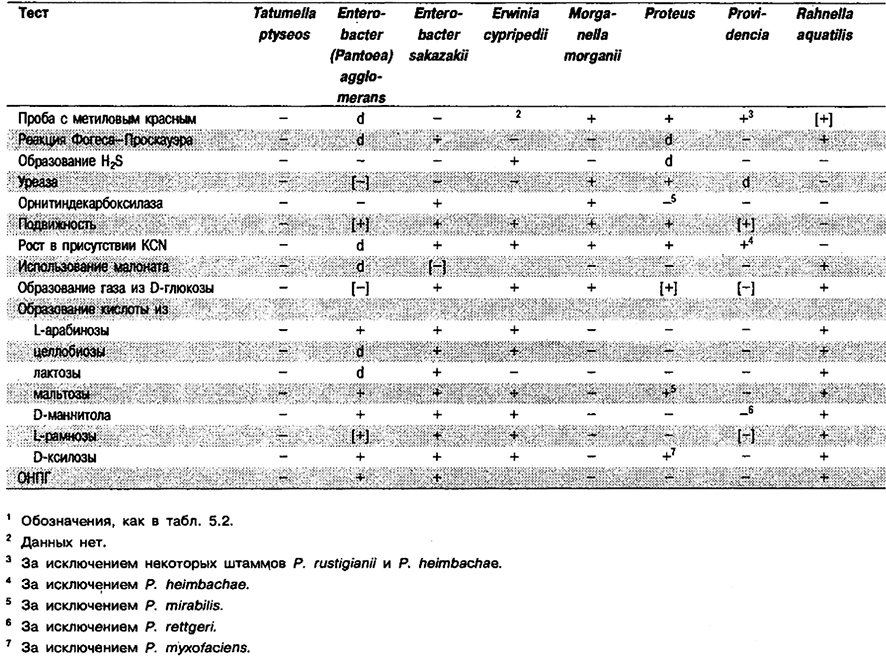
Род Xenorhabdus
Палочки, 0,3–2 х 2–10 мкм, иногда нити длиной 15–50 мкм. В старых культурах клетки содержат кристаллические включения (не поли-β-гидроксибутират). В последней трети экспоненциальной фазы роста встречаются сферопласгы, или кокковидные тела, образующиеся в результате дезинтеграции клеточной стенки. Грамотрицательные. Подвижные за счет перитрихнальных жгутиков. Факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура 25°С; при 36°С растут плохо или не растут. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты, но не газа; процесс слабо выражен или протекает с задержкой, и кислота образуется в небольших количествах. Оксидазотрицательные; каталазоотрицательные за исключением X. luminescens. Большинство штаммов индолотрицательные. Проба с метиловым красным и реакция Фогеса-Проскауэра отрицательные. Лизин- и орнитиндекарбоксилаза, а также аргининдигидролаза отсутствуют. H2S не образуют; большинство штаммов не гидролизует мочевину; в присутствии KCN не растут; малонат не используют. Нитрат, как правило, не восстанавливают. Единственный из углеводов, сбраживаемый почти всеми штаммами, – D-маниоза. Все виды за исключением одного образуют различные пигменты. Для всех штаммов характерны фазовые варианты (различные формы колоний). Естественные места обитания – просвет кишечника у энтомопатогенных нематод и внутренние полости тела у насекомых, инфицированных этими нематодами. Один геновид выделен из ран и крови человека (Fanner et al., J. Clin. Microbiol. 27: 1594–1600, 1989).
Типовой вид: Xenorhabdus nematophilus.
От редакции: Данные о биохимических признаках представителей Xenorhabdus, приводимые разными группами исследователей, существенно различаются (Akhurst, Boemare, J. Gen. Microbiol. 134: 1835–1845, 1988; Farmer et al., J. Clin. Microbiol. 27: 1594–1600, 1989; Grimont et al., Int. J.Syst. BacterioL 34: 378–388, 1984). Вероятно, многие различия обусловлены замедленностью и слабой выраженностью реакций, а также различиями в используемых средах. Полностью биохимические признаки установлены только у X. luminescens и X. nematophilus. В семействе Enterobacteriaceae нет родов, фенотипически сходных с Xenorhabdus. По источникам выделения, плохому росту или его отсутствию при 37°С и слабой активности биохимических процессов в целом род Xenorhabdus фенотипически уникален.
Дифференциация видов рода Xenorhabdus, См. табл. 5.39. Дополнительные признаки приведены в табл. 5.2.
Таблица 5.39. Дифференциация видов рода Xenorttabdus1,2
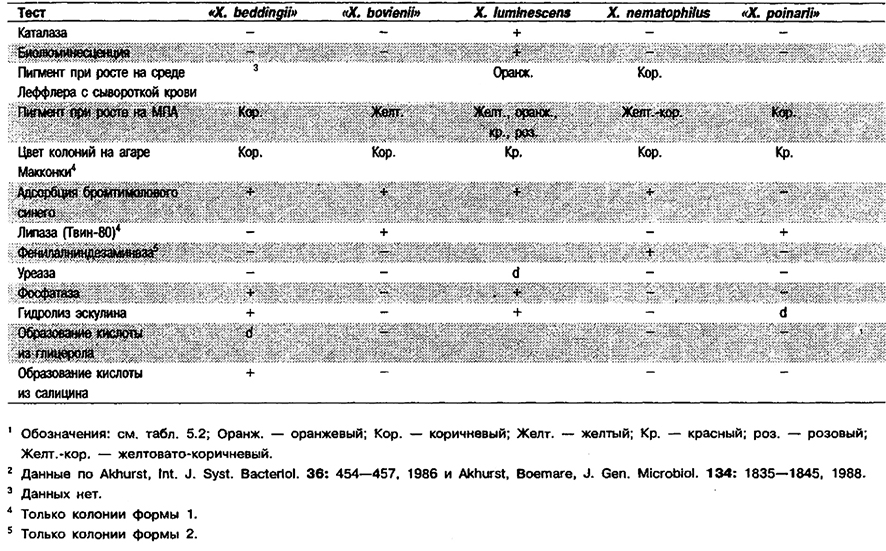
От редакции: Виды «Xenorhabdus beddingii», «Xenorhabdus bovienii» и Xenorhabdus poinarii» не приведены в «Bergey's Manual of Systematic Bacteriology». Эти виды описаны в 1988 г. (Akhurst, Boemare, J. Gen. Microbiol. 134: 1835–1845), но это опубликование не узаконено. Внутри вида X. luminescens выделено 4 дополнительных геновид а, не получивших названия, поскольку по фенотипическим признакам они неотличимы от Xluminescens (Farmer et al., J. Clin. Microbiol. 27: 1594– 1600, 1989).
Род Yersinia
Прямые палочки, иногда приобретающие сферическую форму, диаметром 0,5–0,8 и длиной 1–3 мкм. Грамотрицательные. Неподвижные при 37°С, но при температуре ниже 30°С подвижные за счет перитрихиальных жгутиков; исключение составляют некоторые штаммы Y. ruckeri и вид Y. pest is, представители которого всегда неподвижные. Факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура 28–30°С. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты и небольшого количества газа или газ не образуют. Оксидазоотрицательные; каталазоположительные; по образованию индола виды различаются. Проба с метиловым красным обычно положительная. Реакция Фогеса-Проскауэра отрицательная; на среде Симмонса с цитратом при 37°С не растут, а при 25–28°С результаты варьируют. По лизиндекарбоксилазе и аргининдигидролазе, как правило, отрицательные; по карбоксилазе положительные, за исключением Y. pestis, Y. pseudotuberculosis и Y. rohdei. H2S не образуют. Мочевину обычно гидролизуют, за исключением Y, bercovieri, Y. pestis и Yruckeri. Небольшое число штаммов растет в присутствии KCN; малонат не используют. Восстанавливают нитрат. Сбраживаемые всеми или большинством видов углеводы включают L-арабинозу, мальтозу, D-маннитол, D-маннозу и трегалозу. Встречаются в разнообразных местах обитания, включая человека, животных, особенно грызунов и птиц, почву, воду, молочные и другие пищевые продукты. Y. pestis – возбудитель чумы, болезни, главным образом, диких грызунов. Переносчиками Y. pestis, распространяющими возбудитель среди диких грызунов, служат блохи, в которых бактерии размножаются, закупоривая пищевод и глотку. Блохи отрыгивают бактерии при очередном кровососании и при этом могут передавать возбудитель, если не находят других хозяев, человеку. В результате укуса инфекционных, блох у человека развивается типичная бубонная форма чумы и может возникнуть вторичная пневмония. При воздушно-капельной передаче инфекции может распространяться первичная легочная чума. Вид К pseudotuberculosis патогенен для многих видов животных и иногда для человека; его представители вызывают мезентеральный лимфаденит, хроническое желудочно-кишечное расстройство и тяжелую септицемию, Представители Y. enterocolitica вызывают сходные заболевания у животных и человека. Бактерии Y. ruckeri вызывают болезнь «красный рот» у рыб. Другие виды в отдельных случаях вызывают оппортунистические инфекции у человека или непатогенны.
Типовой вид: Yersinia pestis.
Признаки, отличающие Yersinia от других родов, приведены в табл. 5.40. Следует также учитывать помещенные в табл. 5.41 данные по ключевым признакам Yersinia для двух температур – 28°С и 36°С, поскольку многие виды Yersinia метаболически более активны при более низкой температуре инкубации.
Дифференциация видов рода Yersinia: См. табл. 5.42. Дифференциация различных биогрупп Y. enterocolitica представлена в табл. 5.43. Полный перечень биохимических признаков для условий 37°С приведен в табл. 5.2.
Таблица 5.40. Отличия Yersinia от других радов1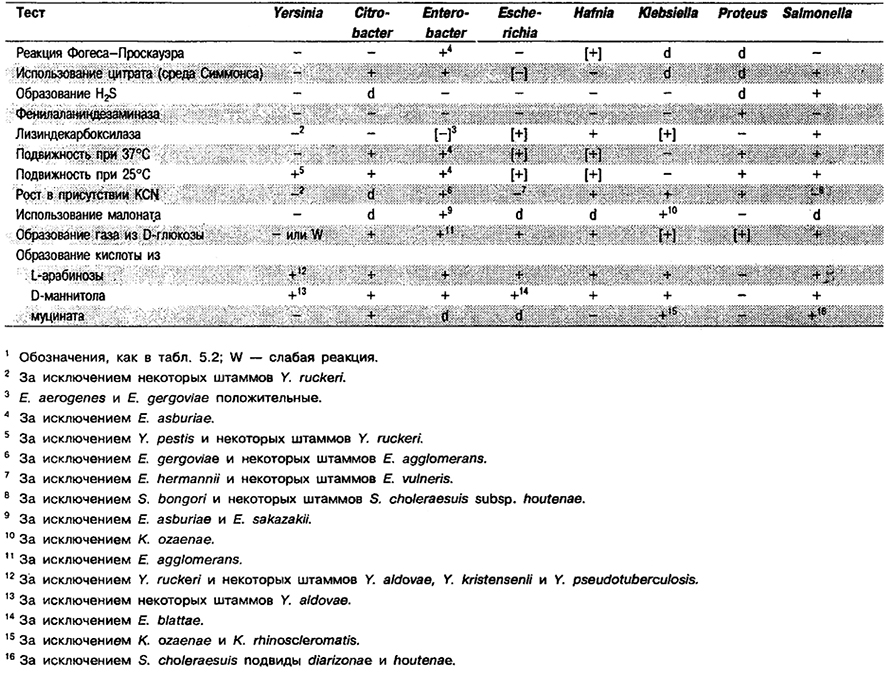
Таблица 5.41. Реакции йерсиний при 25–28°С и 37°С1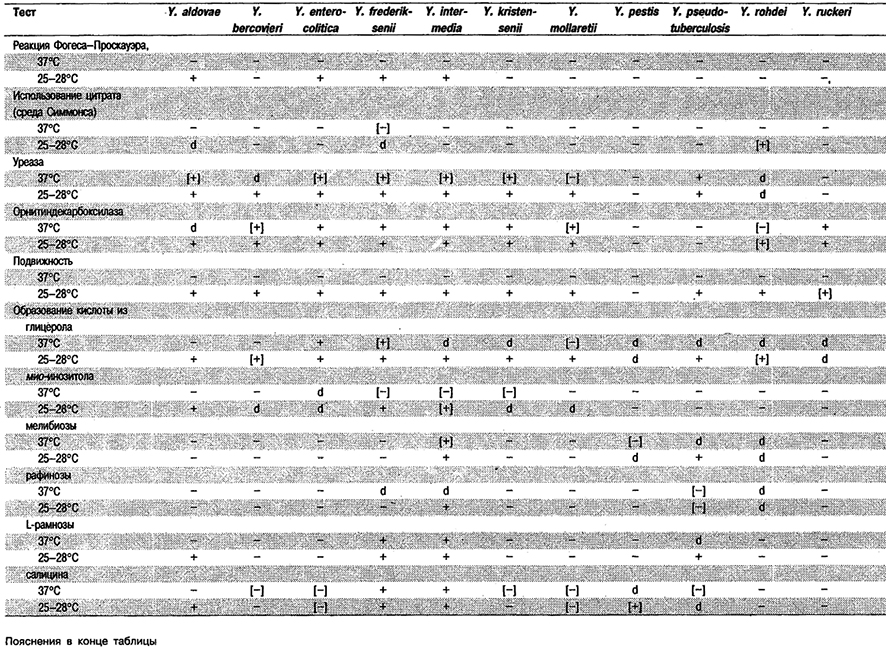
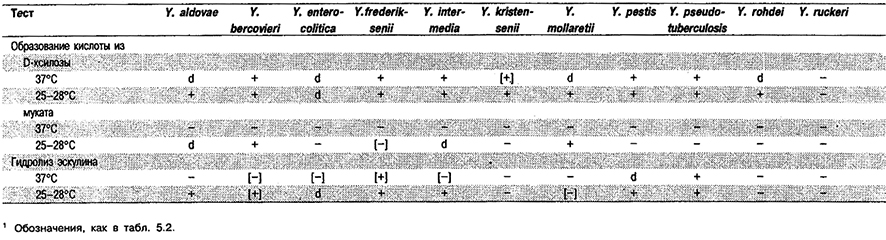
Таблица 5.42. Дифференциация видов рода Yersinia1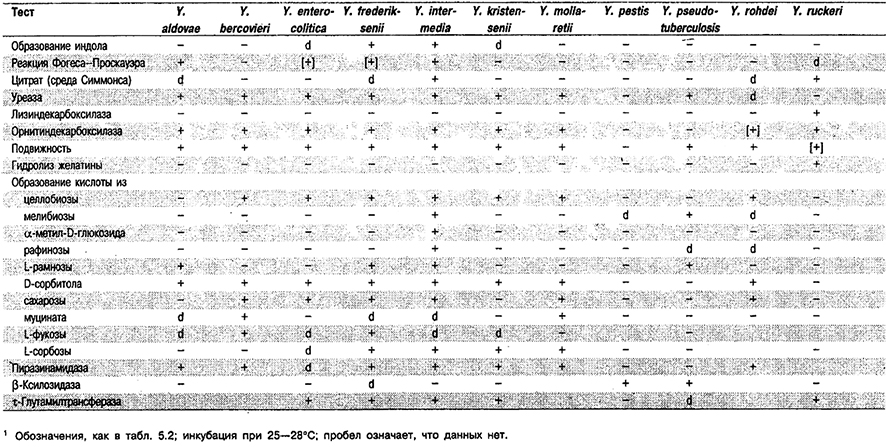
Таблица 5.43. Дифференциация биотипов вида Yersinia enterocolitis 1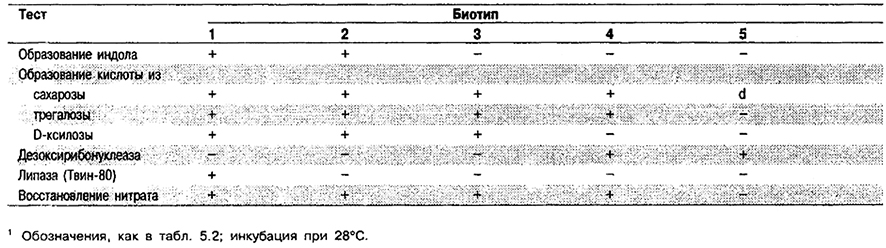
От редакции: Yersinia aldovae, Yersinia bercovieri, Yersinia mollaretii и Yersinia rohdei не приведены в «Bergey's Manual of Systematic Bacteriology». Вид Yersinia aldovae описан в 1984 г. (Bercovier et al., Int. J. Syst. Bacteriol. 34: 166–172). Ранее этот вид был известен как группа Х2 внутри вида Yersinia enterocolitica. Виды Yersinia bercovieri и Yersinia mollaretii описаны в 1988 г. (Wauters et al., Int. J. Syst, Bacteriol. 38: 424–429). Вид Yersinia bercovieri ранее носил название Yersinia enterocolitica биогруппа 3B, a Yersinia mollaretii – Yersinia enterocolitica биогруппа ЗА. Вид Yersinia rohdei описан в 1987 г. (Aleksic et al., Int. J. Syst. Bacteriol. 37: 327–332).
Род Yokenella
От редакции: Род Yokenella не приведен в «Sergey's Manual of Systematic Bacteriology». Этот род описан в 1984 г. (Kosako et al., Japan J. Med. Sci. Biol. 37: 17–124; Int. J. Syst. Bacteriol, 35: 223– 225, 1985) и включает один вид К regensburgei. Род Koserella, включающий единственный вид К. trabulsii, описан в 1985 г. (Hickman–Brenner et al., J, Clin, Microbiol. 21: 39–42; Int. J. Syst. Bacteriol. 35: 223–225, 1985). К regenburgei и К. trabulsii в 1987 г. были определены как субъективные синонимы (Kosako et al., Int. J. Syst. Bacteriol. 37: 127– 129). Для этого организма существуют и другие синонимы: Hafnia группа 3 по гибридизации, Enteric Group 45 (CDC), NIH биогруппа 9 (Japan). Хотя название К. trabulsii имело приоритет в связи с более ранним упоминанием в алфавитном порядке в Int. J. Syst. Bacteriol. (Одобренный список № 17), обе группы исследователей согласились, что название рода должно быть Yokenella.
Прямые палочки: Грамположительные. Подвижные за счет перитрихиальных жгутиков. Факультативные анаэробы. Хемоорганотрофы, обладающие и дыхательным и бродильным типами метаболизма. Оптимальная температура 37°С. Катаболизируют D-глюкозу и другие углеводы с образованием кислоты и газа. Оксидазоотрицательные; каталазоположительные; индолотрицательные; реакция Фогеса-Проскауэра отрицательная, проба с метиловым красным положительная; растут на среде Симмонса с цитратом; по лизин- и орнитиндекарбоксилазе положительные; по аргининдигидролазе отрицательные. H2S не образуют; мочевину не гцдролизуют; не растут в присутствии KCN; малонат не используют. Восстанавливают нитрат. Сбраживаемые углеводы включают L-арабинозу, D-ксилозу, мальтозу, D-маннитол, D-маннозу, мелибиозу, L-рамнозу, трегалозу и целлобиозу. Источники выделения – раны, моча, мокрота и кал человека, а также кишечник насекомых. Клиническое значение неизвестно.
Типовой (и единственный) вид: Yokenella regensburgei.
Признаки вида: Первоначально выделенные у человека и насекомых штаммы Yokenella относили к Hafnia alvei или Hafnia-подобным видам. В табл. 5.44 приведены признаки, позволяющие дифференцировать Yokenella от Hafnia. Дополнительные биохимические тесты приведены в табл, 5,2. Род Yokenella по некоторым признакам сходен с видами родов Citrobacter и Escherichia. Часто штаммы Yokenella в нескольких биохимических тестах дают положительные реакции с задержкой (3–7 сут) (табл. 5.45).
Таблица 5.44. Отличия Yokenella от Hafnia 1
Таблица 5.45. Положительные реакции с задержкой у Yokenella regensburgei 
