Эта группа в общем соответствует по составу семейству Bacteroidaceae, как оно представлено в «Beigey's Manual of Systematic Bacteriology», т. 1. Вместе с тем группа стала настолько разнообразной, что по существу включенные в нее организмы объединяет только то, что их считают анаэробами и по Граму они окрашиваются отрицательно. На самом деле не все представители группы анаэробы, например виды Wolinelta, Bacteroides gracilis и Bacteroides ureolyticus – это, как теперь установлено, микроаэрофильные организмы, способные к кислородному дыханию и включенные в Группу 6 из соображений удобства. Хотя организмы этой группы окрашиваются по Граму отрицательно, некоторые из них (например, Butyrivibrio), как показывают данные электронной микроскопии, имеют клеточную стенку грамположительного типа.
Ряд представителей группы обладает особыми свойствами. Так, для одного рода, Sporomusa, характерно образование устойчивых к нагреванию эндоспор, восемь родов составляют термофильные бактерии, два рода – экстремально галофильные.
Представители группы встречаются в разнообразных местообитаниях, но основные источники их выделения – ротовая полость или кишечный тракт человека и животных либо бескислородные илы и ил сточных вод.
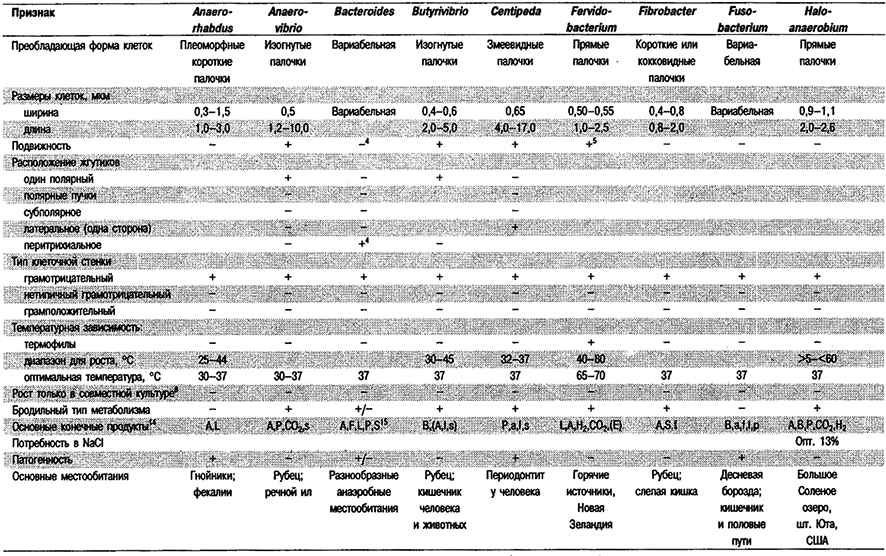
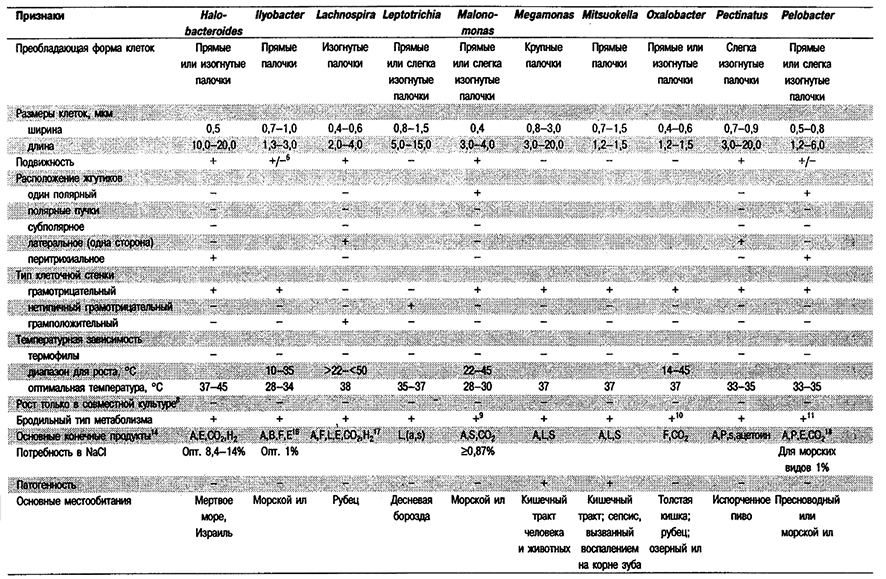
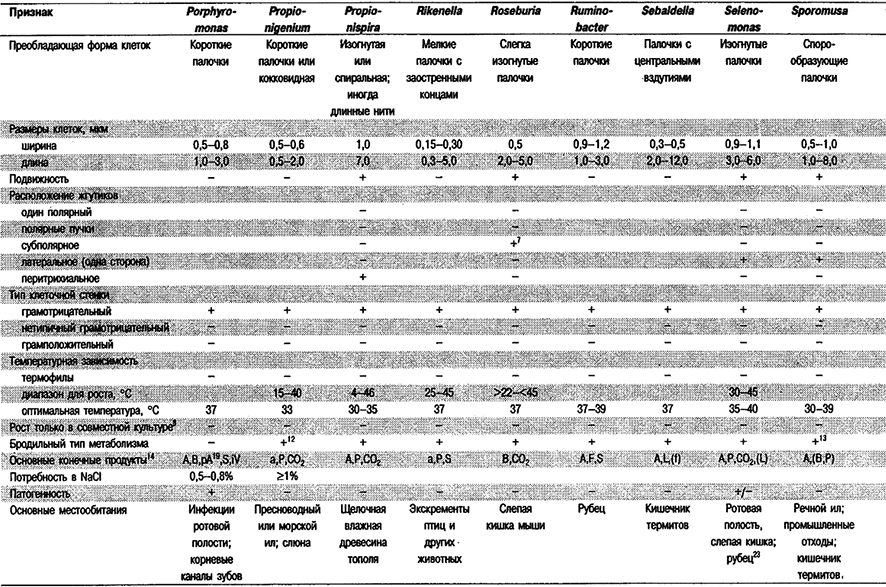
Дифференциация родое Группы 6: См. табл. 6.1.
Таблица 6.1. Дифференцирующие признаки грамотрицательных,
анаэробных, прямых, изогнутых и спиральных бактерий1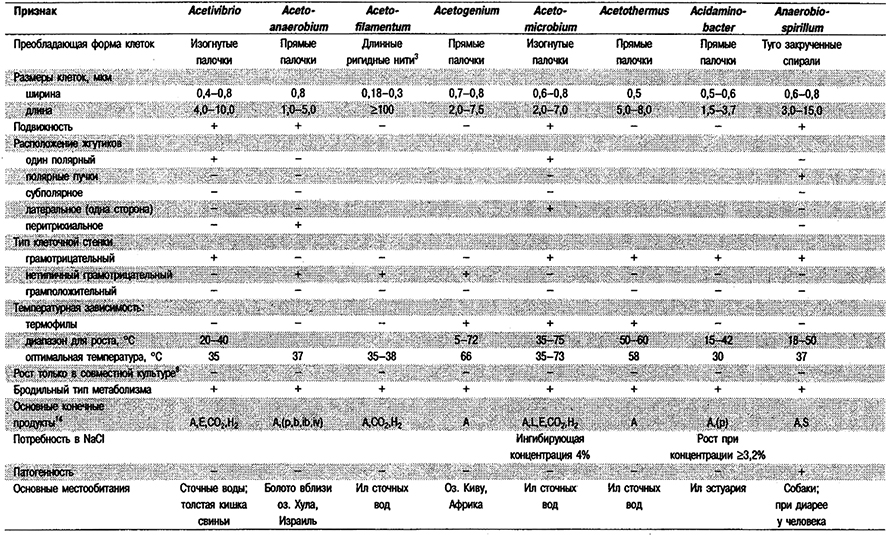

Род Acetivibrio
Слегка изогнутые палочки, 0,4–0,8 х 4–10 мкм, в парах или коротких цепочках. Подвижные за счет единственного жгутика на вогнутой стороне клетки, прикрепленного в точке, делящей эту сторону клетки в соотношении 2:1, или за счет пучка жгутиков в середине вогнутой стороны клетки. Мезофилы; растут лучше всего при 35°С. Хемоорганотрофы сбраживают углеводы с образованием в основном ацетата. Другие продукты брожения – этанол, СО2 и Н2. Пропионат и лактат не образуют. Выделены из ила сточных вод и экскрементов свиней.
Типовой вид: Acetivibrio cellulolyiicus.
Дифференциация видов рода Acetivibrio: См. табл. 6.2.
Таблица 6.2. Дифференцирующие признаки видов рода Acetivibrio1
Род Acetoanaerobium
От редакции: Род Acetoanaerobium не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1985 г. (Sleat et al.t Int. J. Syst. Bacteriol. 35: 10–15) и включает единственный вид A. noterae.
Прямые палочки, 0,8 х 1,0–5,0 мкм, Грамотрицательные, но имеют клеточную стенку нетипичного строения и не лизируются под действием КОН. Неспоро образующие. Подвижные за счет 3–4 перитрихиальных жгутиков. Сбраживают ограниченное количество субстратов, в том числе дрожжевой экстракт, глюкозу и мальтозу. При росте в среде с глюкозой и мальтозой образуют только ацетат; в среде с дрожжевым экстрактом образуют, кроме того, пропионат, бутнрат, изобутират и и зовалерат. В присутствии 80% Н2 и 20% СО2 в составе газовой фазы и 0,2% дрожжевого экстракта образуют ацетат из Н2 и СО2. Оптимальная температура 37°С; оптимальный рН 7,6. Выделены из болотного ила, район оз. Хула, Галилея, Израиль.
Типовой (и единственный) вид: Acetoanaerobium noterae.
Отличия Acetoanaerobium от других Н2-окисляющих ацетогенных бактерий: см. Sleat et al., Int. J. Syst. Bacteriol. 35: 10-15, 1985, табл. 2.
Признаки вида: Как для рода.
Род Acetofilamentum
От редакции: Род Acetofilamentum не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1988 г. (Dietrich et al., Syst, Appl. Microbiol 10: 273–278; Int. J, Syst. Bacteri- ol. 39: 93, 1989) и включает единственный вид А, rigidum.
Длинные, тонкие, ригидные нити, шириной 0,18–0,30 мкм и длиной 100 мкм или более. Грамотрицательные, но имеют клеточную стенку, нетипичную по составу ни для грамотрицательных, ни для грамположительных бактерий. Неспорообразующие. Под действием додецилсульфата натрия нити распадаются на отдельные клетки длиной 2–7 мкм, что указывает на присутствие вокруг нитей белкового чехла. Неподвижные. Оптимальная температура 35–38°С Хемоорганотрофы; используют несколько различных гексоз, пентоз или аминокислот; нуждаются для роста в дрожжевом экстракте. Глюкозу сбраживают до ацетата, СО2 и Н2; возрастающие концентрации Н2 в газовой фазе (5–20%) пропорционально ингибируют рост. Выделены нз ила городских сточных вод.
Типовой (и единственный) вид: Acetofilamentum rigidum.
Признаки вида: Как для рода.
Род Acetogenium
От редакции: Род Acetogenium не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1983 г. (Leigh, Wolfe, Int. J. Syst. Bacteriol. 33: 886) и включает единственный вид A. kivui.
Прямые палочки, 0,7–0,8 х 2,0–7,5 мкм. По Грамму окрашиваются отрицательно, но клеточная стенка грамположительного типа. Неспорообразующие; неподвижные. Термофилы; оптимальная температура 66°С; диапазон температуры 50–72°С. Оптимальный рН 6,4. Сбраживают глюкозу, маннозу, пируват и фруктозу, образуя практически только ацетат. Ацетат образуют также из Н2 и СО2 (80% Н2 и 20% СО2) давление 2 атм). Для роста нуждаются в сульфиде и/или цистеине. Выделены из озерного ила.
Типовой (и единственный) вид; Acetogenium kivui.
Признаки вида: Как для рода. Вид выделен из ила оз. Киву, Африка.
Род Acetomicrobium
От редакции: Род Acetomicrobium не приведен в «Beigey's Manual of Systematic Bacteriology». Этот род был описан в 1984 г. (Soutschek et al., Syst. Appl. Microbiol. 5: 377–390; Int. J. Syst. Bacteriol. 35: 223, 1985) и первоначально включал единственный вид A. fiavidum. В 1987 г, в него был добавлен второй вид A. faecalis (sic) (Winter at al., Syst. Appl. Microbiol. 9: 71–76; Int. J. Syst. Bacteriol 38: 136, 1988).
Изогнутые палочки, 0,6–0,8 x 2,0–7,0 мкм. Неспорообразующие. Подвижные за счет единственного субполярного жгутика или нескольких латеральных жгутиков. Термофилы; оптимальная температура 58–73°С. Хемоорганотрофы. Сбраживают разнообразные гексозы и пентозы. Нуждаются для роста в дрожжевом экстракте. Глюкозу сбраживают до ацетата, СО2 н Н2 или ацетата, лактата, этанола, СО2 и Н2. Выделены из ила сточных вод.
Типовой вид: Acetomicrobium fiavidum.
Дифференциация видов рода Acetomicrobium: См. табл. 6.3.
Таблица 6.3. Дифференцирующие признаки видов рода Acetomicrobium1
Род Acetothermus
От редакции: Род Acetothermus не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1988 г. (Dietrich et al., Syst, Appl. Microbiol. 10: 174–179; Int. Syst. Bacteriol. 38: 328, 1988) и включает единственный вид А. paucivorans.
Прямые палочки, 0,5 х 5–8 мкм при достаточной концентрации витамина В12. В отсутствие этого витамина образуют длинные нити (>50 мкм). Неспорообразующие, неподвижные. Термофилы; оптимальная температура 58°С; диапазон температуры 50–60°С. Хемоорганотрофы. Нуждаются для роста в дрожжевом экстракте и, особенно, витамине В12. Сбраживают глюкозу и фруктозу до ацетата, СО2 и Н2. Другие сахара не катаболизируют. Выделены из сточных вод, сбраживаемых при 60°С.
Типовой (и единственный) вид: Acetothermus paucivorans.
Бактерии рода Acetothermus сходны с бактериями Acetomicrobium по местообитанию (сточные воды) и образованию ацетата, СО2 и Н2 из глюкозы, но при этом отличаются от них тем, что а) не обладают подвижностью, б) нуждаются в витамине В12, в) сбраживают только глюкозу и фруктозу и г) характеризуются более высоким содержанием G+С в ДНК (62 против 48 мол.%).
Признаки вида: Как для рода.
Род Acidaminobacter
От редакции: Род Acidaminobacter не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1984 г, (Stams, Hansen, Arch. Microbiol. 137: 329–337; Int. J. Syst. Bacteriol. 35: 223, 1985) и включает единственный вид А. hydrogenoformans.
Прямые палочки, 0,5–0,6 х 1,5–3,7 мкм, с заостренными концами, одиночные или в парах. Неподвижные. Оптимальная температура 30°С; диапазон температуры 15–42°С. Хемоорганотрофы. Сбраживают аминокислоты, особенно активно глуха миновую кислоту, с образованием в качестве основного конечного продукта ацетата и в меньших количествах NH3, формиата, СО2, Н2, пропионата и иногда других жирных кислот в зависимости от используемой аминокислоты. Лактат, суюшнат, бутират и этанол не образуют. Образуют индол из триптофана. Рост на многих субстратах стимулируется и может зависеть от присутствия бактерий, использующих Н2, таких как Methanospirillum или Desulfovibrio. Выделены из черного ила эстуария.
Типовой (и единственный) вид: Acidaminobacter hydrogenoformans.
Бактерии рода Acidaminobacter можно отличить от выделенных из бескислородного ила бактерий Pelobacter, представляющих собой грамотрицательные анаэробные мезофильные палочки, по следующим признакам; 1) клетки Acidaminobacter имеют заостренные, а не закругленные концы; 2) они используют различные аминокислоты, тогда как штаммы Pelobacter используют только необычные субстраты, такие как галловая кислота, этиленгликоль или полиэтилен гликоль и 3) штаммы Acidaminobacter образуют индол из триптофана, тогда как штаммы Pelobacter не образуют индол. Выделенные из сточных вод бактерии рода Syntrophobacier, представляющие собой грамотрицательные неподвижные палочки, отличаются от Acidaminobacter тем, что растут только в совместной культуре с Desulfovibrio и используют только пропионат.
Признаки вида: Как для рода.
Род Anaeroblospirilfum
Спиральные клетки с округлыми концами, 0,6–0,8 х 3–15 мкм; длина витка примерно 1,5–2,0 мкм. Иногда встречаются клетки длиной до 32 мкм. Обладают подвижностью по типу ввинчивания, обеспечиваемой биполярными пучками жгутиков. Осевых фибрилл не имеют. Неспорообразующие. Хемоорганотрофы. Сбраживают углеводы, главным образом до уксусной и янтарной кислот. Выделены из глотки и толстого кишечника гончих собак и у человека при диарее.
Типовой (и единственный) вид: Anaerobiospirillum succiniproducens.
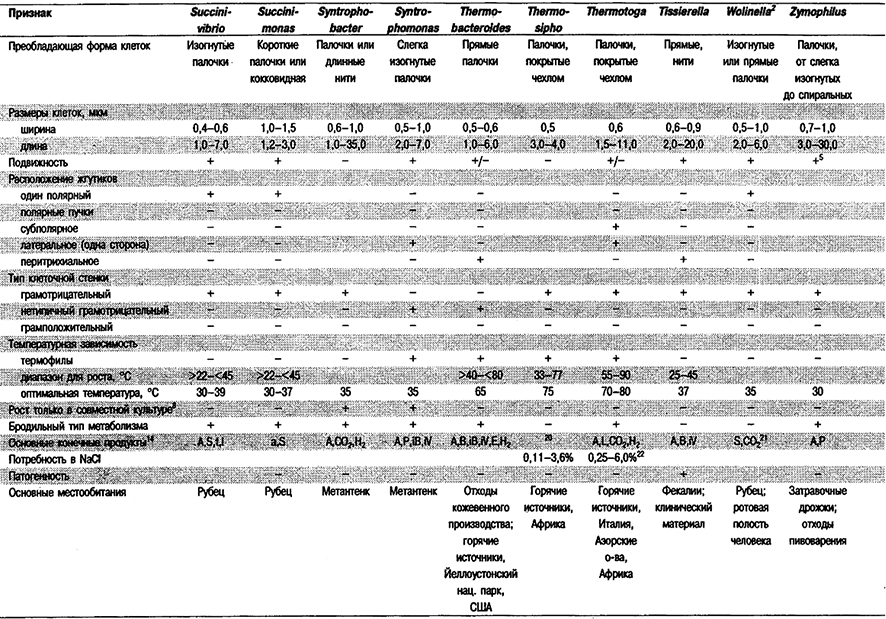
Признаки, отличающие Anaerobiospirillum от сходных родов грамотрицательных подвижных организмов, представлены в табл. 6.4.
Таблица 6.4. Отличия Anaerobiospirillum от сходных грамотрицательных подвижных организмов1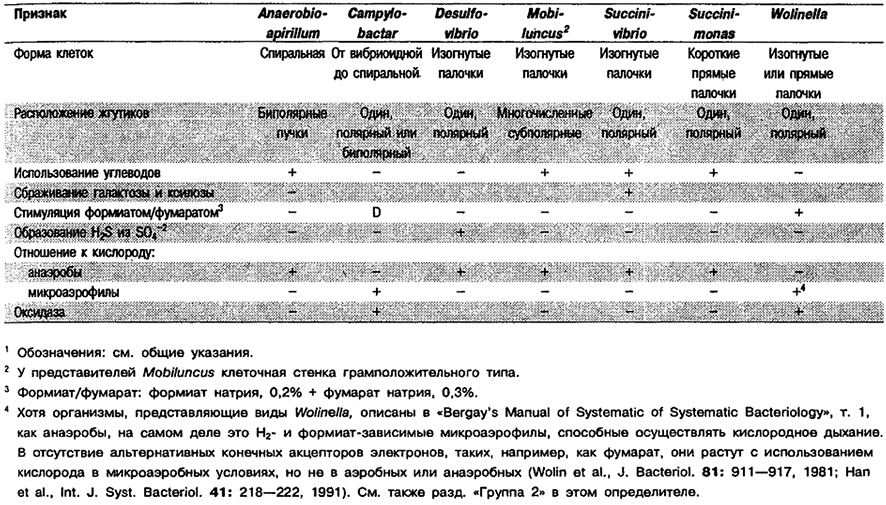
Признаки вида: Большинство штаммов сбраживает глюкозу, лактозу, рафинозу и сахарозу; некоторые могут также образовывать кислоту из фруктозы, мальтозы, трегалозы, мелибиозы и других сахаров. Весьма сходные организмы были обнаружены при септицемии у человека (Park et al., Am. J. Clin. Pathol. 85: 73–76, 1986) и у больных диареей (Malnick et al., J. Clin. Microbiol. 28: 1380–1384, 1990). Штаммы A. succiniproducens выделяют из экскрементов здоровых собак и кошек, но не здоровых людей. Штаммы, выделенные при диарее у человека, отличаются от исходных штаммов, выделенных у гончих собак, по характеру брожения и, возможно, представляют отдельный вид. Предложена селективная среда для Anaerobiospirillum (Malnick et. al., J. Clin, Microbiol. 28: 1380–1384, 1990). Из разнообразных обычно используемых лабораторных сред наилучший рост обеспечивает «шоколадный» агар (Park et ai,, А. J. Clin. Pathol. 85: 73-76, 1986).
Род Anaerorhabdus
От редакции: Род Anaerorhabdus не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1986 г. (Shah, Collins, Syst. Appl. Microbiol. 8: 86–88; Int. J. Syst. Bacteriol. 36: 573, 1986) и включает единственный вид A. furcosus (прежнее название – Bacteroides furcosus).
Плеоморфные короткие палочки, 0,3–1,5 х 1–3 мкм, одиночные, в парах или коротких цепочках; некоторые клетки вилкообразной или Y- образной формы. Неспорообразующие; неподвижные. В основном несахаролитические, хотя слабо сбраживают несколько углеводов. Главные конечные продукты метаболизма – уксусная и молочная кислоты.
Сфинголипидов и менахинонов не содержат; негидроксилированные клеточные жирные кислоты в основном не разветвленные, насыщенные и ненасыщенные; метил разветвленные кислоты отсутствуют или присутствуют лишь в следовых количествах. Выделены из абсцесса аппендикса, при легочных и брюшных абсцессах, а также в ряде случаев из фекалий человека и свиньи.
Типовой (и единственный) вид Anaerorhabdus furcosus.
Anaerorhabdus отличается от Bacteroides fragilis и сходных видов отсутствием сфинголипидов и менахинонов, а также тем, что для него характерны негидроксилированные жирные кислоты преимущественно не разветвленного строения.
Признаки вида: Слабо сбраживают глюкозу, фруктозу и иногда сахарозу. Гидролизуют эскулин. Оптимальная температура 30–37°С. Жидкость рубца вызывает стимуляцию роста, желчь оказывает ингибирующее действие.
Род Anaerovibrio
Изогнутые или спиральные палочки, 0,5 х 1,2–10,0 мкм. Неспорообразующие. Подвижные за счет единственного полярного жгутика. Оптимальная температура 30–37°С. Хемоорганотрофы; используют очень ограниченное число сахаров; конечные продукты брожения – пропионат, ацетат и сукцинат. Некоторые штаммы используют только глицерол или диолеин, образуя только пропионат. Другие штаммы типично липолитические – гидролизуют триглицериды до глицерола и жирных кислот и образуют в основном пропионат и сукцинат из глицерола. Обнаружены в рубце крупного рогатого скота и овец, в бескислородных пресноводных илах и в иле сточных вод.
Типовой вид: Anaerovibrio lipolytica.
Дифференциация видов рода Anaerovibrio: См. табл. 6,5. Признаки, отличающие A. lipolytica от морфологически сходных подвижных анаэробных организмов рубца, указаны в табл. 6.6.
Таблица 6.5. Дифференцирующие признаки видов рода Anaerovibrio1
Таблица 6.6. Отличия Anaerovibrio lipolytica
от морфологически сходных подвижных анаэробных бактерий рубца1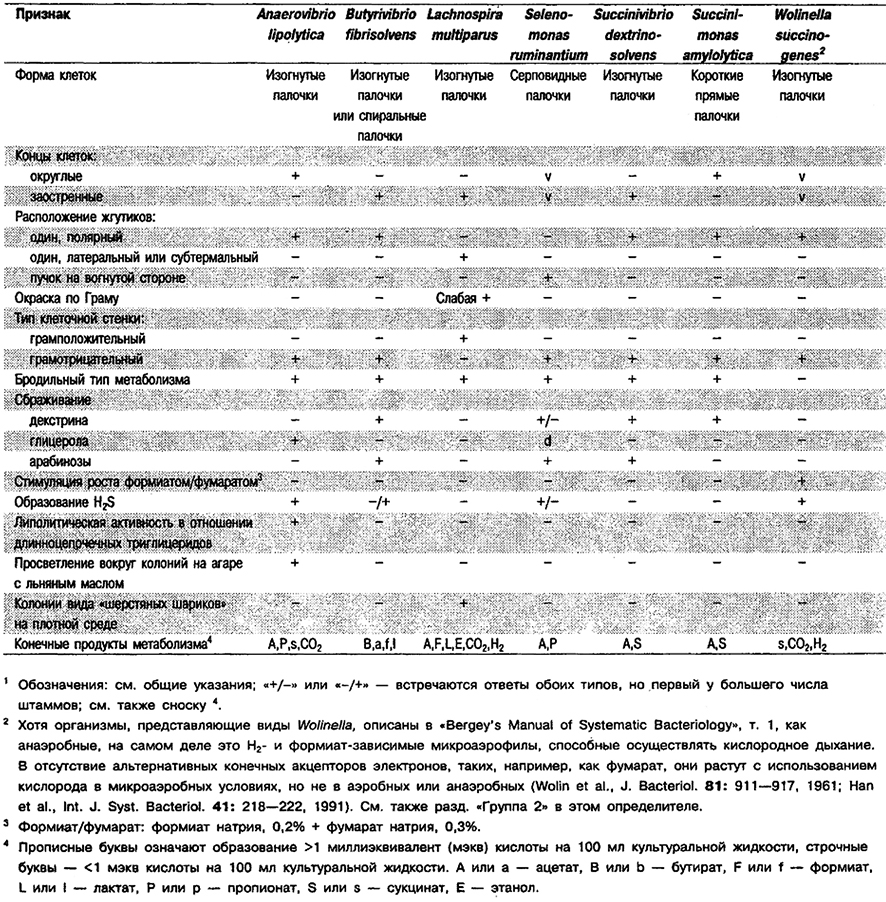
Род Bacteroides
Палочковидные организмы различных размеров. У многих видов клетки плеоморфные, с терминальными или центральными вздутиями; содержат вакуоли и образуют нити. Как правило, неподвижные (бактерии двух видов подвижные, другие могут обладать подвижностью типа рывков). Анаэробы (см., однако, указание ниже); хемоорганотрофы; метаболизируют углеводы, пептон или промежуточные продукты метаболизма. Продукты брожения включают ацетат, сукцинат, лактат, формиат или пропионат, особенно характерные для строго сахаролитических видов. Бутират обычно не является основным продуктом, но если образуется, то присутствуют также изобутнрат и изовалерат. Многие виды отличаются высоким содержанием разветвленных жирных кислот, в основном С15-кислот антеизо-строения, а также сфинголипидов (Shah, Collins, J. Appl. Bacterid. 55: 403–416, 1983). Рост многих видов значительно стимулируют гемин и витамин К, которые обычно добавляют в среды для выращивания бактероидов. Источниками выделения служат разнообразные анаэробные местообитания; десневая борозда, пищеварительный тракт (слепая кищка и рубец), ил сточных вод, очаги инфекции и нагноения у человека и животных.
Типовой вид: Bacteroides fragilis.
Хотя в «Bergey's Manual of Systematic Bacteriology», т. 1, B. ureolyticus и В, gracilis описаны как анаэробы, на самом деле они Н2- и формиат-за- висимые микроаэрофилы, способные к кислородному дыханию. В отсутствие альтернативных конечных акцепторов электронов, таких как фумарат, они растут за счет использования кислорода при его низком содержании в среде, но не в анаэробных или аэробных условиях (Wolin et al., J: Bacteriol, 81: 911–917, 1961; Han et al., Int. J. Syst. Bacteriol. 41: 218–222, 1991).
У двух из описанных видов, 5. polypragmatus и В. xylanolyticus, клетки подвижные за счет перитрихиальных жгутиков; бактерии В. galacturonicus также имеют перитрихиальные жгутики, но, по-видимому, не обладают подвижностью. У двух других видов, В. ureolyticus и В. gracilis, описана подвижность типа рывков, но не настоящее движение с перемещением. Описание подвижности типа рывков см. в оригинальной статье (Henrich- sen, Acta Pathol. Scand. В. 83: 171–178, 1975).
Как считали прежде, бактерии рода Bacteroides отличает от большинства других грамотрицательных организмов (в частности, от Fusobacterium) отсутствие кетодезоксиоктоната (КДО) в составе липополисахарида. Однако на самом деле бактерии рода Bacteroides содержат КДО, но в фосфорилированной форме, поэтому его можно выявить только после гидролиза под действием сильной кислоты (Kurhada et al., FEMS Microbiol. Lett. 51: 77-80, 1988).
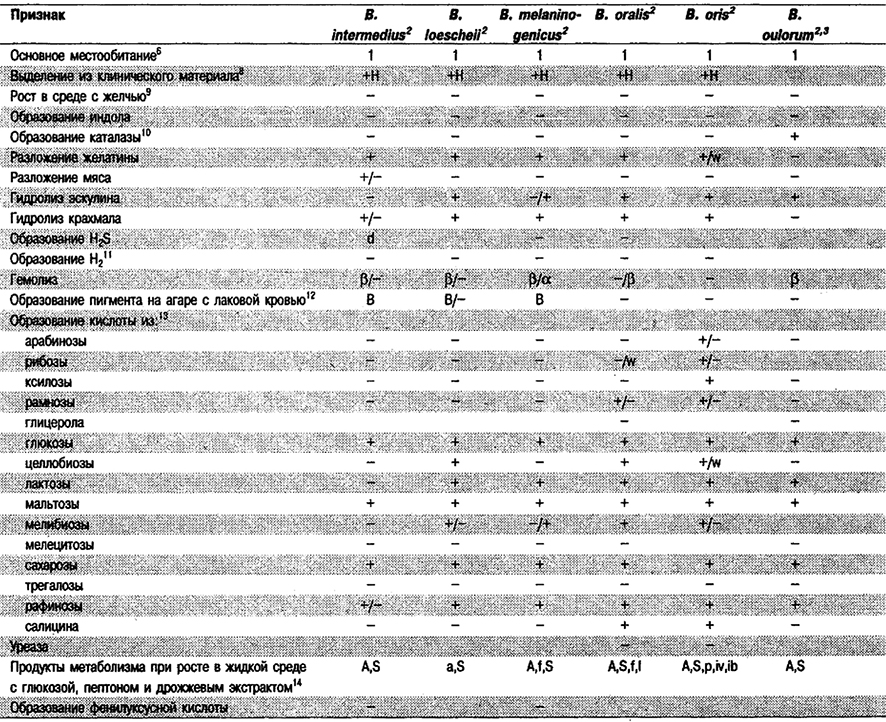
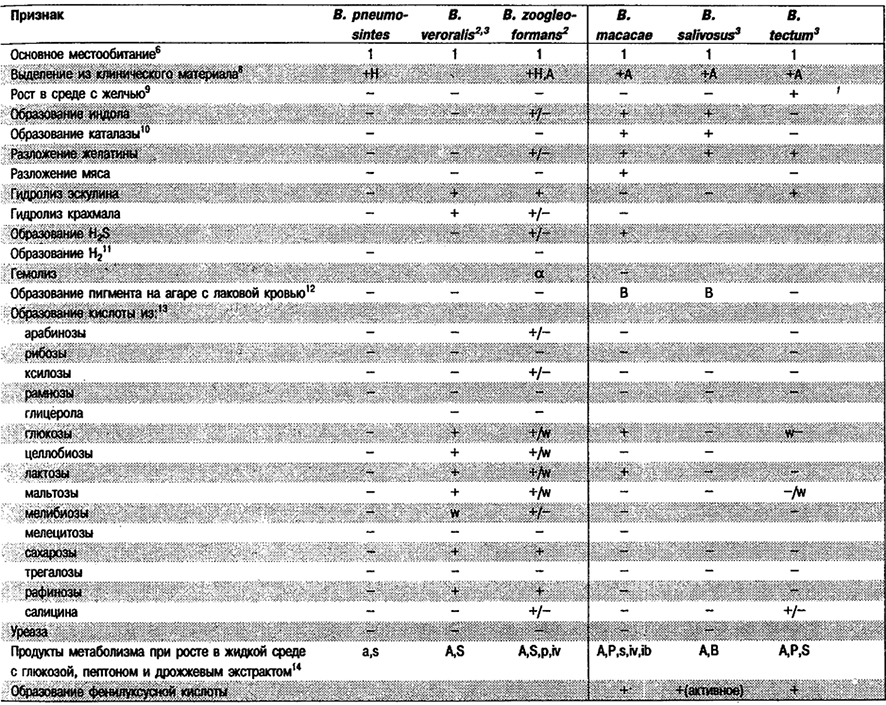
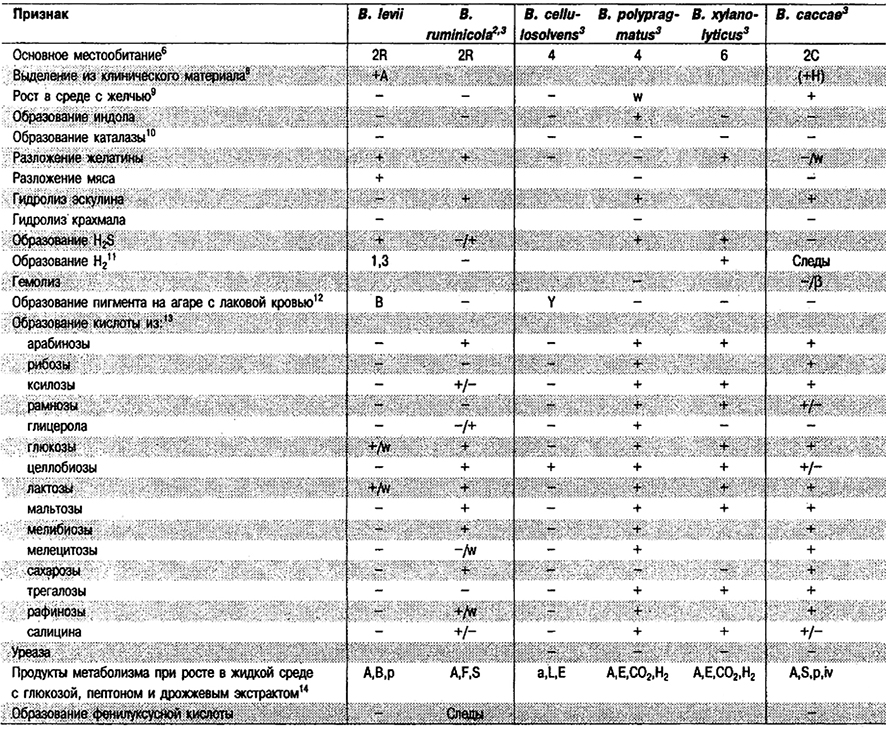
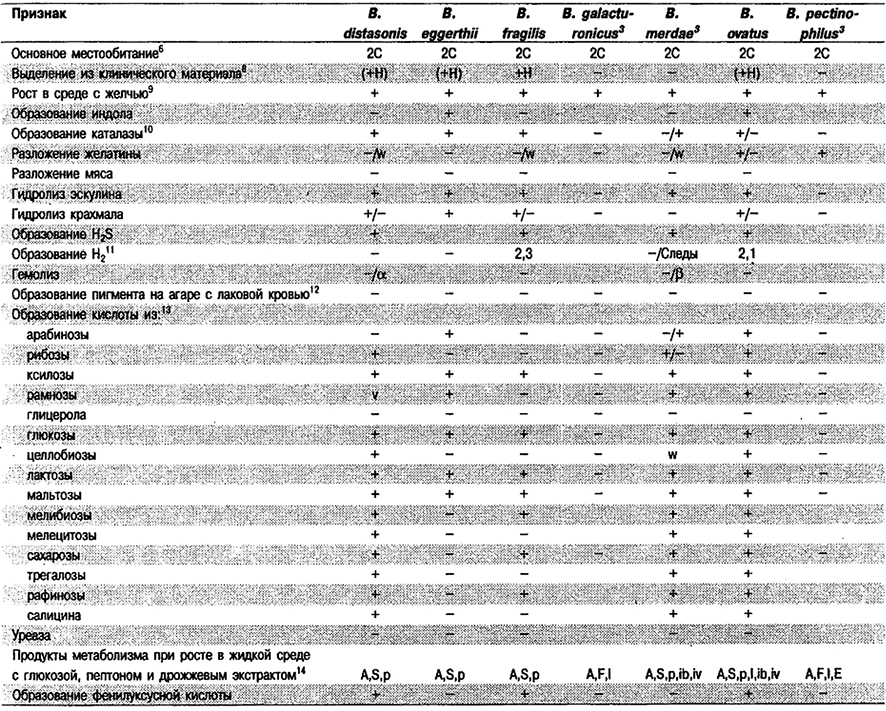
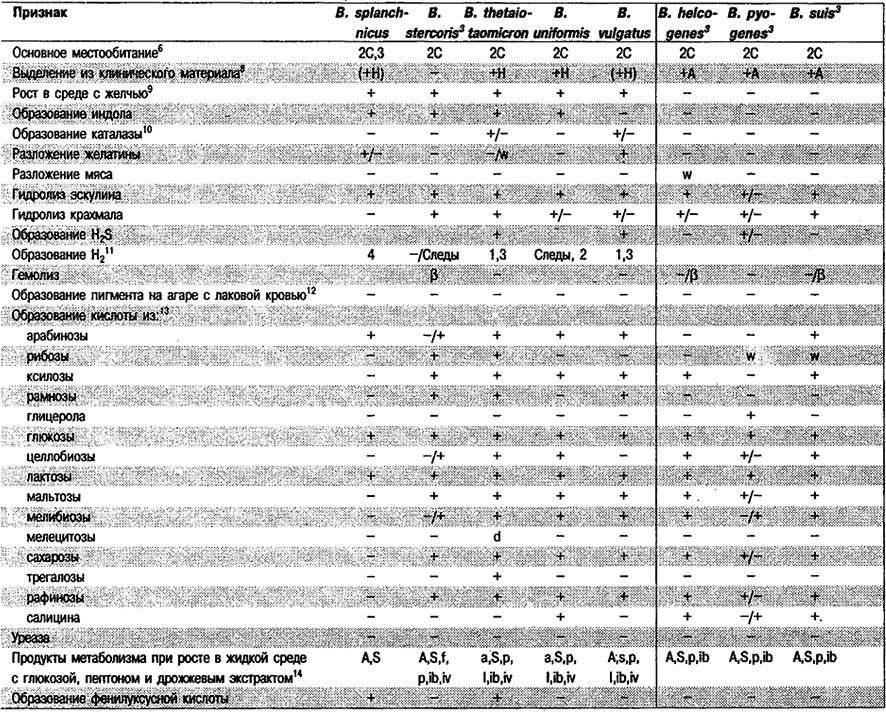
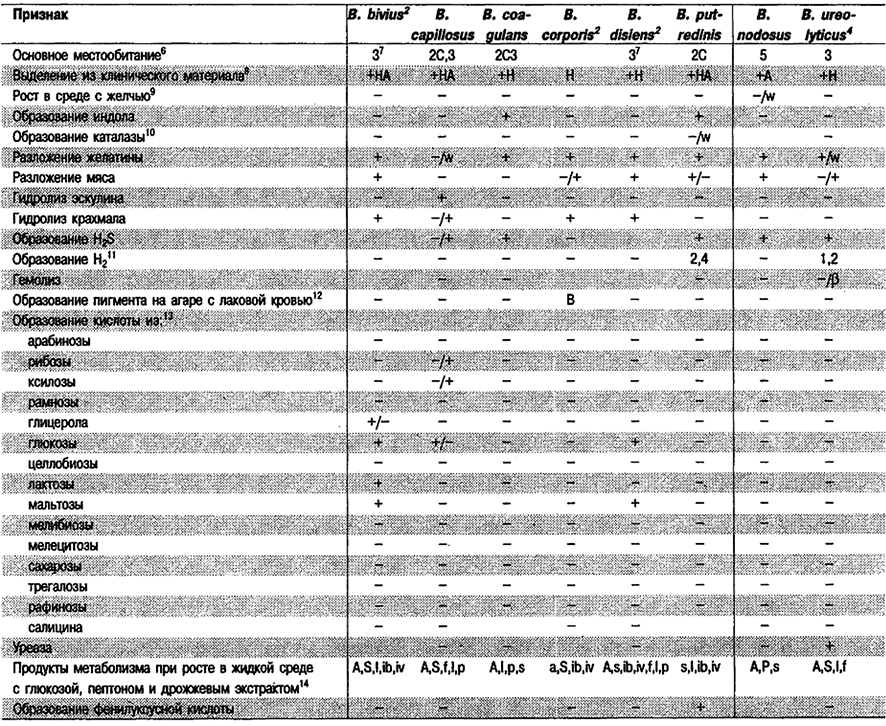
Дифференциация видов рода Bacteroides: См. табл. 6.7.
Таблица 6.7. Дифференцирующие признаки видов рода Bacteroides1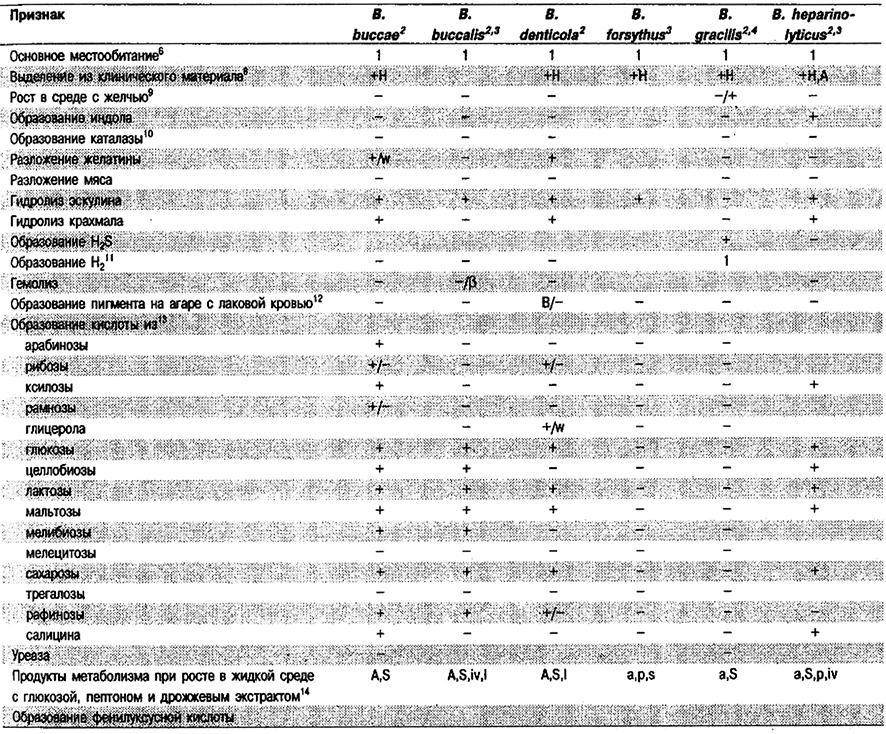

Род Butyrivibrio (см. также разд. «Группа 20»)
Изогнутые палочки, 0,4–0,6 х 2-5 мкм. Грамотрица тельные, но клеточная стенка грампо ложи тельного типа, с необычно тонким слоем пептидоглнкана (Cheng, Costerton, J. Bacteriol. 129: 1506–1512, 1977; Cheng et al., Can. J. Microbiol. 35: 274–282, 1989). Подвижные за счет нескольких полярных или субполярных жгутиков. Метаболизм бродильного типа. Используют ряд углеводов, образуя в качестве основного конечного продукта масляную кислоту. Большинство штаммов образует внеклеточные полисахариды; у некоторых клетки окружены капсулами (Stack et al., Appl. Environ. Microbiol. 54: 878–883, 1988).
Типовой вид; Butyrivibrio fibrisolvens.
Дифференциация видов рода Butyrivibrio: См. табл. 6.8.
Таблица 6.8. Дифференцирующие признаки видов рода Butyrivibrio1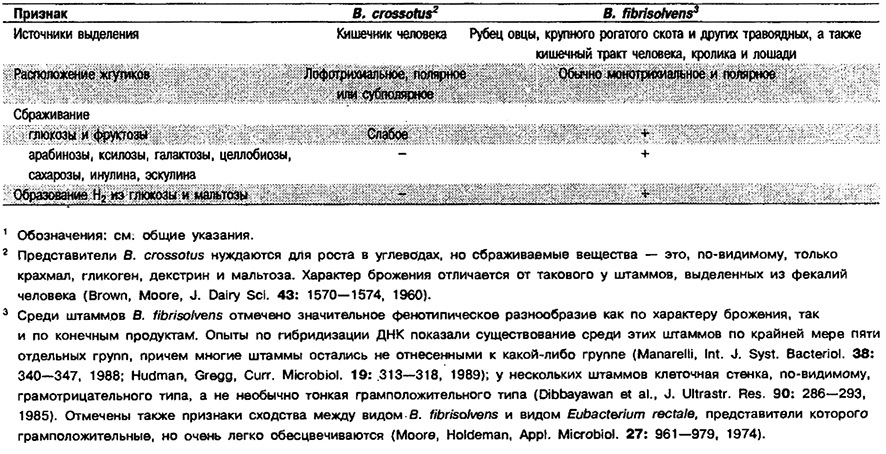
Род Centipeda
От редакции: Род Centipeda не приведен в «Bergey's Manual of Systematic Bacteriology», Этот род описан в 1983 г. (Lai et al, Int. J, Syst, Bacteriol. 33: 628–635) и включает единственный вид С. periodontii.
Змеевидные палочки, 0,65 х 4–17 мкм. Неспо- рообразующие. Подвижные за счет жгутиков, расположенных по спирали вдоль клетки (Males et al., J. Gen. Microbiol. 130: 185–191, 1984). Движение осуществляется путем изгибания всей клетки и вращения ее вокруг продольной оси. Диапазон температуры для роста 32–37°С. Хемоорганотрофы. Сахаролитики, способные сбраживать разнообразные сахара с образованием кислоты, но не газа. Основной продукт разложения сахаров – пропионовая кислота, образуются также в небольших количествах уксусная, молочная и янтарная кислоты. Выделены из субгингивальной области при периодонтите у человека.
Типовой (и единственный) вид: Centipeda periodontii.
Признаки вида: Как для рода. С. periodontii можно ошибочно принять за выделенные из ротовой полости штаммы видов Selenomonas. Главные различия приведены в табл. 6.9.
Таблица 6.9. Отличия Centipeda periodontii от видов Selenomonas, выделенных из ротовой полости1
Род Fervidobacterium
От редакции: Род Fervidobacterium не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1985 г. (Patel et al., Arch. Microbiol. 141: 63-69; Int. J. Syst. Bacteriol. 35: 535, 1985) и включает два вида, F. nodosum и F. islandicum,
Преимущественно прямые палочки, 0,50–0,55 х 1,0–2,5 мкм, одиночные, в парах или коротких цепочках. У многих клеток имеются терминальные вздутия (сфероиды). Встречаются, кроме того, округлые тела, содержащие несколько цитоплазматических единиц. Не спорообразующие; подвижные (расположение жгутиков не описано). Термофилы. Оптимальная температура 70°С; диапазон температуры 40–80°С. Хемоорганотрофы; сбраживают разнообразные сахара до ацетата, лактата, СО2, Н2 и этанола. Выделены из горячих источников, расположенных в Роторуа и Ваймангу, Новая Зеландия, а также в Исландии.
Типовой вид: Fervidobacterium nodosum.
Дифференциация видов рода Fervidobacterium: Два вида, F. nodosum и F. islandicum, различаются по соотношению нуклеотидов в составе ДНК, гомологии ДНК/ДНК и способности использовать целлюлозу (вид F. islandicum положительный).
Род Fibrobacter
От редакции: Род Fibrobacter не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1988 г. (Montgomery et al., Int. J. Syst. Bacteriol. 38: 430–435) и включает два вида, F, succinogenes (прежнее название – Bacteroides succinogenes) и F. intestinalis.
Палочки или плеоморфные овальные клетки, 0,4–0,8 х 0,8–2,0 мкм. Неспорообразуютцие, Неподвижные. Сбраживают целлюлозу и целлобнозу, но лишь небольшое число других сахаров, что составляет характерную особенность. Основные продукты брожения – уксусная и янтарная кислоты; иногда образуется небольшое количество муравьиной кислоты. Нуждаются для роста в СО2, летучих жирных кислотах, аммиаке и одном или более витаминах. Выделены из желудочно-кишечного тракта у млекопитающих.
Типовой вид: Fibrobacter succinogenes.
Дифференциация видов рода Fibrobacter: См. табл. 6.10.
Таблица 6.10. Дифференцирующие признаки видов рода Fibrobacter1
Род Fusobacterium
Палочки веретенообразной или другой формы» часто очень плеоморфные. Неспорообразующие. Неподвижные. Хемоорганотрофы; метаболизируют пептон или углеводы, но способность к брожению обычно лишь слабо выражена. Основной конечный продукт метаболизма – бутират, часто вместе с ацетатом и лактатом; в меньших количествах образуют пропионат, сукцинат и формиат. Изобутират и изовалерат не образуют. Встречаются главным образом в десневой борозде, кишечнике и половых путях, выделены также из культур крови и различных очагов нагноения у человека и животных, а также из тропических язв.
Типовой вид: Fusobacterium nucleatum.
Отличия Fusobacterium от других родов неподвижных анаэробных организмов: Согласно классическому описанию, бактерии рода Fusobacterium – это грамотрицательные, облигатно анаэробные палочки в форме веретена, образующие большие количества масляной кислоты. Однако не все фузобактерии имеют типичную веретенообразную форму; их иногда трудно отличить от штаммов Bacteroides или от штаммов Clostridium и Eubacterium, клетки которых легко обесцвечиваются. Культуры необходимо исследовать во всех фазах роста, проверить на устойчивость к нагреванию и окрасить для выявления спор. Бактерии рода Bacteroides, как правило, не образуют бутират в качестве конечного продукта, а если образуют, то вместе с изобутиратом и изовалератом. Представители Leptotrichia образуют большие количества молочной кислоты, но не бутират.
Все исследованные штаммы Fusobacterium чувствительны к фосфомицину в концентрации 300 мкг/мл, тогда как почти все штаммы Bacteroides устойчивы к нему (Bennet, Duerden, J. Appl. Bacteriol. 59: 171 – 181, 1985). (См. также Rodriguez et a!., J. Appl. Bacteriol. 41: 251–254, 1974; Essers, J. Appl. Bacteriol. 52: 319-323, 1982.)
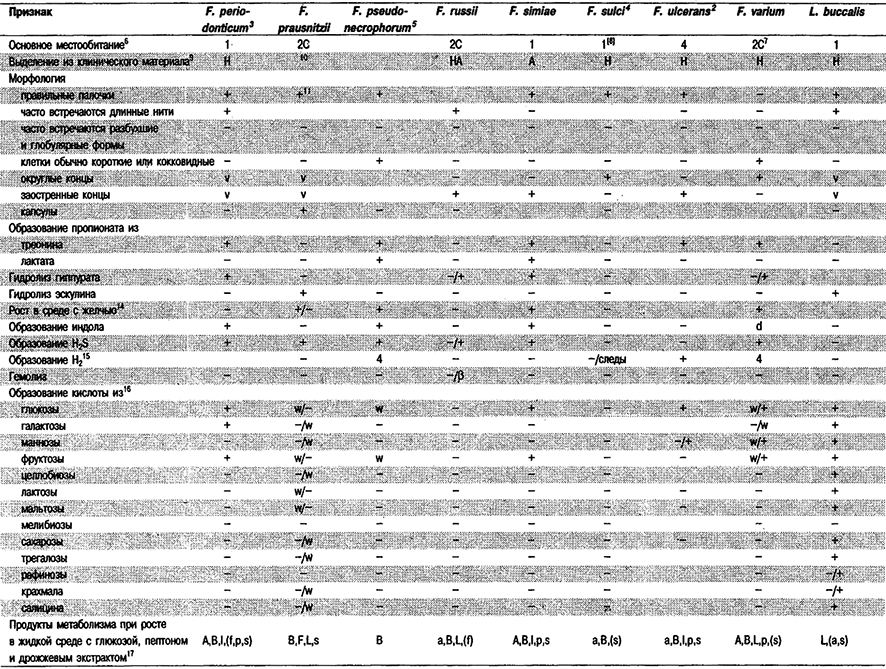
Дифференциация видов рода Fusobacterium: См. табл. 6.11. Как видно из таблицы, одна из проблем при идентификации представителей данного рода состоит в том, что большинство их не проявляет активности в обычных тестах. Данные по составу жирных кислот (газовая хроматография) (Jantzen, Hofstad, J. Gen. Microbiol. 123: 163–171, 1981), электрофорезу глутаматдегидрогеназы (ГДГ) (Gharbia, Shah, J. Gen. Microbiol. 134: 327–332, 1988), а также пиролизу и масс-спек- трометрии (Magee et al., J. Med. Microbiol. 28: 236, 1989) позволяют выделить группы, которые хорошо соответствуют уже признанным видам. Результаты электрофореза ГДГ и пиролиза показывают также, что группа штаммов F. nucleatum гетерогенна, и это детально подтверждают данные по гибридизации ДНК и электрофорезу растворимых белков (Dzink et al., Int, J. Syst. Bacteriol. 40: 74–78, 1990). Авторами последней работы предложены три подвида F. nucleatum: subsp. nucleatum, subsp. polymorphum и subsp. vincentii. С помощью обычно используемых фенотипических тестов эти подвиды различить невозможно.
Таблица 6.11. Дифференцирующие признаки видов Fusobacterium и Leptotrichia buccalis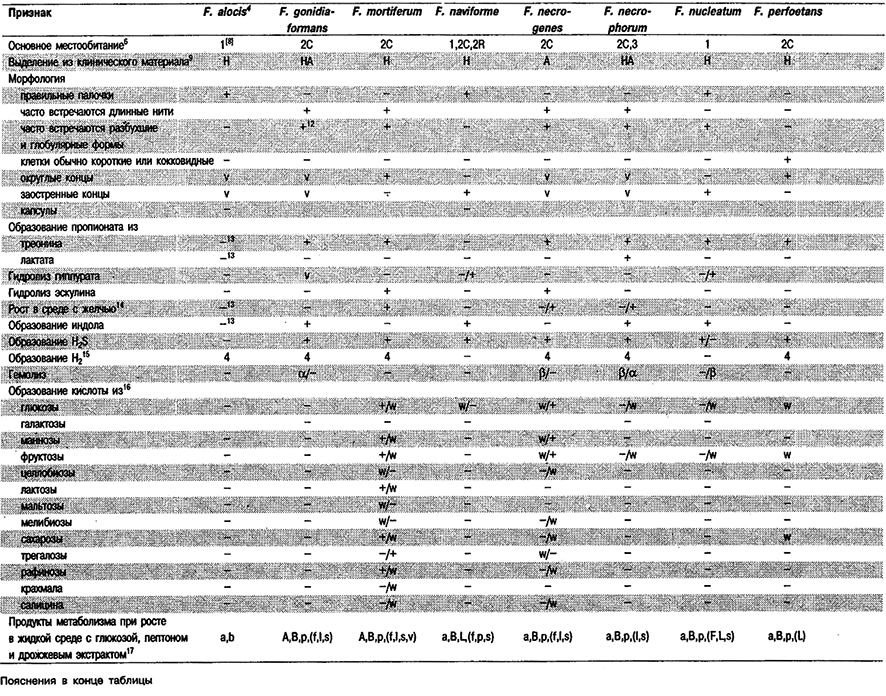

Род Haloanaerobium
От редакции: Род Haloanaerobium не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1983 г. (Zeikus et al, Curr. Microbiol. 9: 225–234; Int. J. Syst. Bacteriol. 34: 503, 1984) и включает единственный вид Н. praevalens.
Прямые палочки, 0,9–1,1 х 2,0–2,6 мкм. Неспорообразующие; неподвижные. Галофилы; лучше всего растут в присутствии 13% NaCI. Оптимальная температура 37°С. Хемоорганотрофы; используют углеводы, пектин, аминокислоты и аминосахара. Основные продукты брожения – ацетат, бутират, пропионат, Н2 н СО2. Выделены из бескислородных осадков Большого Соленого озера, шт. Юта, США.
Типовой (и единственный) вид: Haloanaerobium praevalens.
Признаки вида: Как для рода. Отличия Н. praevalens от Halobacteroides halobius представлены в табл. 6.12.
Таблица 6.12. Отличия Halobacteroides halobius от Haloanaerobium praevalens1
Род Halobacteroides
От редакции: Род Halobacteroides не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1984 г. (Оrеn et al., Syst. Appl. Microbiol. 5: 58–70; Int. J. Syst. Bacteriol, 3: 355, 1984).
Прямые или изогнутые палочки, 0,5 х 10–20 мкм. Неспорообразующие. Подвижные за счет перитрнхиальных жгутиков. Галофилы; лучше всего растут в присутствии 8,4–14% NaCI. Оптимальная температура 37–42°С. Хемоорганотрофы; сбраживают рад Сахаров и некоторые органические кислоты. Конечные продукты сбраживания глюкозы – ацетат, этанол, Н2 и СО2. Выделены из осадков Мертвого моря.
Типовой (и единственный) вид: Halobacteroides halobius.
Признаки вида: Клетки выглядят гибкими и могут иметь завиток на одном из концов, а в старых культурах принимают сферическую форму. Чувствительны к хлорамфениколу (30 мкг/мл) и пенициллину (20 ЕД/мл), Нуждаются для роста в биотине и ламинобензойной кислоте. Другие свойства – как для рода. Отличия Halobacteroides halobius от Hahanaerobium praevalens представлены в табл. 6.12.
Род llyobacter
От редакции: Род llyobacter не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род был описан в 1984 г. (Stieb, Schink, Arch. Microbiol. 140: 139–146; Int. J. Syst. Bacteriol. 35: 375, 1985) и первоначально включал один вид L polytropus. В том же году к нему был добавлен второй вид, I. tartaric us (Schink, Arch. Microbiol. 139: 409–414, 1984; Int. J. Syst. Bacteriol. 35: 375, 1985).
Обычно короткие прямые палочки, 0,7–1,0 х 1,3–3,0 мкм, в парах или цепочках. Неспорообразующие. Могут быть подвижными или неподвижные. Как правило, нуждаются для хорошего роста в 1% NaCl. Оптимальная температура 28–34°С, Хемоорганотрофы; сбраживают разнообразные сахара и органические кислоты, такие как 3-гидроксибутират, кротонат, L-тартрат, цитрат, пируват, малат, фумарат, глюкоза, фруктоза и глицерол, с образованием различных конечных продуктов в зависимости от субстрата. Глюкозу сбраживают до ацетата, формиата и этанола. Выделены из бескислородных морских илов.
Типовой вид: llyobacter polytropus.
Дифференциация видов рода llyobacter: См. табл. 6.13.
Таблица 6.13. Дифференцирующие признаки видов рода llyobacter1
Род Lachnosptra (см. также разд. «Группа 20»)
Изогнутые палочки, 0,4–0,6 х 2,0–4,0 мкм. Грамотрицательные, но клеточная стенка грамположительного типа. Клетки молодых культур окрашиваются по Граму положительно. Неспорообразуюшие. Подвижные за счет одного жгутика, расположенного латерально или субполярно. Хемоорганотрофы; метаболизм бродильного типа. Конечные продукты сбраживания глюкозы – ацетат, формнат, лактат, этанол, Н2 и СО2. При сбраживании пектина дополнительно образуется метанол за счет активности пектинметилэстераэы. Выделены из бычьего рубца.
Типовой (и единственный) вид: Lachnospira multiparus.
Признаки вида: Как для рода. Отличия от Anaerovibrio lipolytica и других морфологически сходных бактерий рубца представлены в табл. 6.6.
Род Leptotrichla
Прямые или слегка изогнутые палочки, 0,8–1,5 х 5–15 мкм. Как правило, окрашиваются по Граму отрицательно, хотя клетки очень молодых культур могут окрашиваться по Граму положительно; клеточная стенка грамотрицательного типа, но отличается от типичной. Неспорообразующие. Неподвижные, Лучше всего растут в присутствии 5–10% СО2 в газовой фазе. Оптимальная температура 35–37°С. При температуре ниже 25°С не растут. Хемоорганотрофы; сбраживают углеводы. Основной конечный продукт разложения глюкозы – молочная кислота. Обнаружены главным образом в зубных бляшках, однако сходные организмы выделены из женских половых путей.
Типовой (и единственный) вид: Leptotrichia buccalis.
Признаки вида: Палочки, по форме сходные с фузобактериями, часто в парах, где соприкасающиеся концы уплощены. Могут встречаться клетки в форме длинных нитей. Для хорошего роста необходима жидкая среда с дрожжевым экстрактом (0,3%), глюкозой (0,5%), гидрохлоридом цистеина (0,1%) и сывороткой крови (5%). Можно использовать среду Казаи (Kasai, J. Dent. Res. 40; 800– 811, 1961). Нуждаются для роста в СО2 (5–10%). Колонии на кровяном агаре через 48 ч имеют форму «голова медузы», диаметр 1–2 мм.
При морфологическом сходстве с некоторыми видами Fusobacterium бактерии L. buccalis в отличие от них сильные сахаролитики, образующие большие количества молочной кислоты (см. табл. 6.11), Другие нитевидные организмы из зубного материала, Corynebacterium matruchotii (Bacterione- ma matruchotii), в отличие от L. buccalis аэробные, грамположительные, по форме похожие на кнутовище.
Род Malonomonas
От редакции: Род Malonomonas не приведен в «Sergey's Manual of Systematic Bacteriology». Этот род описан в 1989 г. (Dehmng, Schink, Arch. Microbiol. 151: 427–433; Int. J. Syst. Bacteriol. 40: 320, 1990) и включает единственный вид М. rubra.
Палочки, от прямых до слегка изогнутых, 0,4 х 3,1–4,0 мкм, с округлыми концами, одиночные, в парах, коротких цепочках или крупных агрегатах. Неспорообразующие. Подвижные за счет 1–2 полярных жгутиков. Оптимальная температура 28–30°С; диапазон 22–45°С. Хемоорганотрофы; лучше всего растут в анаэробных условиях, но устойчивы к концентрации 02 до 5%. Способны использовать для роста в качестве единственного источника углерода и энергии малонат, который декарбоксилируют до ацетата. Сбраживают фумарат и малат до сукцината и СО2. Другие органические кислоты, сахара или спирты не используют. Содержат большое количество цитохрома с. Выделены из бескислородных морских илов.
Типовой (и единственный) вид: Malonomonas rubra.
Признаки вида: Нуждаются для роста по крайней мере в 150 мМ (0,88%) NaCl. Колонии на плотных средах дисковидные, красного цвета за счет большого количества цитохрома. Другие свойства – как для рода.
Род Megamonas
От редакции: Род Megamonas не приведен в «Betsey's Manual of Systematic Bacteriology». Этот род описан в 1982 г. (Shah, Collins, Zentralbl. Bak- teriol. Hyg. C3: 394–398; Int. J. Syst. Bacteriol. 33: 439, 1983) и включает единственный вид М, hypermegas (прежнее название – Bacteroides hypermegas).
Крупные палочки, 0,8–3,0 х 3–20 мкм, с округлыми концами. Содержимое клеток обычно выглядит гранулярным из-за присутствия волютина. Неспорообразующие. Неподвижные. Хемоорганотрофы; сбраживают разнообразные углеводы. Конечные продукты – уксусная, пропионовая и молочная кислоты. Сфинголипидов или менахинонов не содержат; негидроксилированньге жирные кислоты преимущественно не разветвленные. Выделены из кишечника человека, животных и домашней птицы.
Типовой вид: Megamonas hypermegas.
Признаки вида: Активные сахаролитики, но не протеолитики. Гидролизуют эскулин. Другие признаки – как для рода.
Род Mitsuokella
От редакции: Род Mitsuokella не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род был описан в 1982 г. (Shah, Collins, Zentralbl. Bakteriol. Hyg. C3: 491–494; Int. J. Syst. Bacteriol. 33: 439; 1983) и первоначально включал единственный вид М. multiacidus (прежнее название – Bacteroides multiacidus). В 1986 г. в него добавлен второй вид, М. dentalis (Haapasalo et al., Int. J. Syst. Bacteriol. 36: 566–568).
Правильные или овальные палочки, 0,7–1,5 х 1,2–1,5 мкм. Неспорообразующие. Неподвижные. Могут иметь фимбрии; у некоторых штаммов клетки окружены капсулами. Сбраживают углеводы, иногда очень активно. Конечные продуты сбраживания глюкозы – уксусная и янтарная кислоты, а также в том или ином количестве молочная кислота. Сфинголипидов или менахинонов не содержат; разветвленные жирные кислоты присутствуют лишь как минорные компоненты. Выделены из фекалий человека и свиньи, а также у человека из корневых каналов зубов при воспалении.
Типовой вид: Mitsuokella multiacidus.
Дифференциация видов рода Mitsuokella: См. табл. 6.14.
Таблица 6.14. Дифференцирующие признаки видов рода Mitsuokella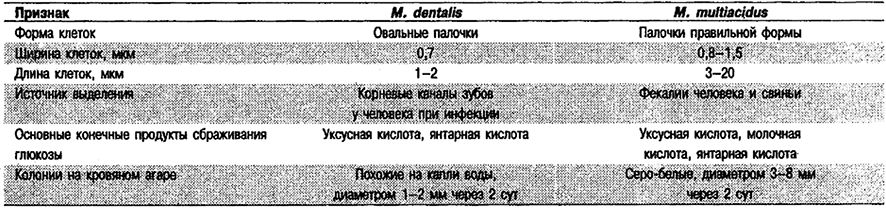
Род Oxalobacter
От редакции: Род Oxalobacter не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1985 г. (Allison et al., Arch Microbiol. 141: 1–7; Int. J. Syst. Bacteriol. 35: 375, 1985) и включает единственный вид О. formigenes.
Прямые или изогнутые палочки, 0,4–0,6 х 1,2–1,5 мкм, с округлыми концами. Неспорообразующие. Неподвижные. Единственным источником углерода и энергии служит оксалат; конечные продукты метаболизма – формиат н СО2. Выделены из рубца и толстой кишки у животных и человека, а также из озерных осадков.
Типовой (и единственный) вид: Oxalobacter formigenes.
Признаки вида: При росте на агаризованной среде, содержащей оксалат кальция, образуют зоны просветления вокруг колоний. Другие свойства – как для рода.
Род Pectinatus
Слегка изогнутые палочки, 0,7–0,9 х 3–30 мкм или больших размеров, с округлыми концами. Неспорообразующие. Подвижные за счет жгутиков, расположенных в виде гребешка на вогнутой стороне клетки. Оптимальная температура 30°С. Сбраживают сахара с образованием уксусной и пропионовой кислот и в небольших количествах янтарной и молочной кислот. Выделены из испорченного пива и задаточных дрожжей.
Типовой вид: Pectinatus cerevisiiphilus.
Дифференциация видов рода Pectinatus; См. табл. 6.15.
Таблица 6.15. Дифференцирующие признаки видов рода Pectinatus1
Род Pelobacter
От редакции: Род Pelobacter не приведен в «Вегgey's Manual of Systematic Bacteriology». Этот род был описан в 1982 г. (Schink, Pfennig, Arch. Microbiol. 133: 195–201; Int. J. Syst. Bacteriol. 33: 896, 1983) и первоначально включал единственный вид jP. acidigallici. Второй вид, P. venetianus, добавлен в 1983 г. (Schink, Stieb, Appl Environ. Microbiol. 45: 1905-1913; Int. J. Syst. Bacteriol. 34: 91, 1984). Еще два вида, P. carbinolicus и P. propionics, добавлены в 1984 г, (Schink, Arch. Microbiol. 137: 33–41; Int. J. Syst. Bacteriol. 34: 356, 1984) и пятый вид, P. acetylenicus, – в 1985 г. (Schink, Arch. Microbiol. 142: 295–301; Int. J. Syst. Bacteriol. 36: 355, 1986).
Прямые или слегка изогнутые палочки, 0,5–0,8 х 1,2–6,0 мкм, с округлыми или слегка заостренными концами, одиночные, в парах или коротких цепочках. Неспорообразующие. У некоторых видов клетки подвижные. Оптимальная температура 33–35°С. Используют ограниченное число субстратов, среди них пирогаллол, 2,3-бутандиол, этиленгликоль, полнэтиленгликоль, ацетилен, ацетонн, пируват и лактат. Обнаружены в бескислородных морских и пресноводных илах.
Типовой вид: Pelobacter acidigallici.
Дифференциация видов рода Pelobacter: См, табл. 6.16.
Таблица 6.16. Отличительные признаки видов рода Pelobacter 1,2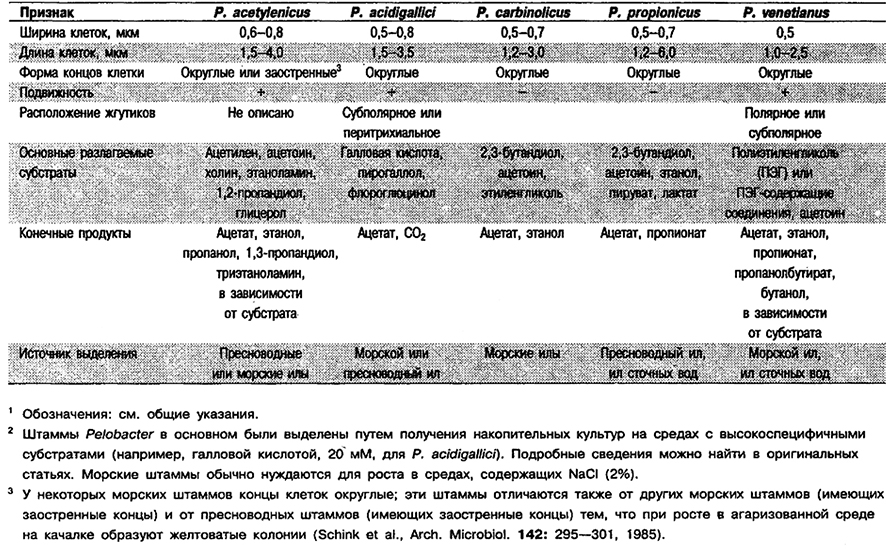
Род Porphyromonas
От редакции: Род Porphyromonas не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1988 г. (Shah, Collins, Int J. Syst. Bacteriol. 38: 128–131) и включает три вида: P. asaccharolytica (прежнее название – Bacteroides asaccharolyticus), P. gingivalis (прежнее название – Bacteroides gingivalis) и P. endodontalis (прежнее название – Bacteroides endodontalis).
Короткие палочки, 0,5–0,8 x 1,0–3,0 мкм. Неспорообразующие; неподвижные. Как правило, на кровяном агаре колонии окрашены в цвета от коричневого до черного вследствие образования протогема. Несахаролитические: присутствие углеводов не влияет существенно на рост, тогда как гидролизаты белков, такие как пептон или дрожжевой экстракт, сильно стимулируют рост. Основные продукты брожения – н-масляная н уксусная кислоты; в небольших количествах образуются пропионовая, изомасляная, изовалериановая и иногда фенилуксусная кислоты. Негидроксилиро- ванные клеточные жирные кислоты в основном метилразветвленные (изо-С15). Выделены при инфекциях из ротовой полости н корневых каналов зубов.
Типовой вид: Porphyromonas asaccharolyticus.
Дифференциация видов рода Porphyromonas: См. табл. 6.17.
Таблица 6.17. Дифференцирующие признаки видов рода Porphyromonas1
Род Prevotella
От редакции: Род Prevotella не приведен в «Вегgey's Manual of Systematic Bacteriology». Этот род описан в 1990 г, (Shah, Collins, Int. J. Syst. Bacteriol, 40: 205–208) с типовым видом P. melaninogenica (прежнее название – Bacteroides melaninogenicus). Из рода Bacteroides в Prevotella перенесено еще 15 видов.
Плеоморфные палочки, неподвижные, неспорообразующие. Облигатные анаэробы. Хемоорганотрофы. Умеренные сахаролитики; основные продукты брожения – ацетат и сукцинат, в небольших количествах образуются изобугират, изовале- рат и лактат. Рост ингибируется желчью в концентрации 20% (масса/объем).
Дифференциация видов рода Prevotella: См, табл. 6.7, (Виды Bacteroides, включенные теперь в Prevotella, отмечены значком 2).
Род Propionigenium
От редакции: Род Propionigenium не приведен в «Beigey's Manual of Systematic Bacteriology». Этот род описан в 1982 г. (Schink, Pfennig, Arch. Microbiol. 133: 209–216; Int. J. Syst. Bacteriol. 33: 896, 1983) и включает единственный вид P. modestum.
Короткие палочки, 0,5–0,6 х 0,5–2,0 мкм, с округлыми концами, одиночные, в парах или коротких цепочках, Неспорообразующие, неподвижные. Оптимальная температура 33°С. Используют для роста органические кислоты с короткой цепью и образуют пропионат как основной конечный продукт. Выделены из морских и пресноводных илов, а также из слюны человека.
Типовой (и единственный) вид: Propionigenium modestum.
Признаки вида: Сбраживают сукцинат, фумарат, малат, аспартат, оксалоацетат и пируват, образуя пропионат, ацетат и СО2. Для роста чистой культуры необходимо, чтобы среда содержала NaCl (1%),
Как грамотрицательные анаэробы, образующие пропионат, представители Propionigenium могут быть ошибочно приняты за Propionispira. Однако Propionigenium – это короткие неподвижные палочковидные клетки, сбраживающие только ограниченный набор огранических кислот и не содержащие цитохромов, тогда как Propionispira – длинные, изогнутые, подвижные клетки, сбраживающие углеводы и содержащие цитохромы.
Род Propionispira
От редакции: Род Propionispira не приведен в- «Beigey's Manual of Systematic Bacteriology». Этот род описан в 1982 г. (Shink et al., J. Gen. Microbiol. 128: 2771-2779; Int. J. Syst, Bacteriol. 33: 673, 1983) и включает единственный вид P. arboris.
Палочки, от изогнутых до спиральных, 1,0 х 7,0 мкм. Могут образовывать длинные спиральные клеточные нити. Неспорообразующие. Подвижные за счет пернтрнхиальных жгутиков. Оптимальная температура 30–33°С. Сбраживают разнообразные углеводы, образуя в основном пропионат, ацетат и СО2. Могут расти за счет использования N2 как единственного источника азота и восстанавливать ацетилен до этилена. Содержат цитохром b. Выделены нз щелочной влажной древесины тополя.
Типовой (и единственный) вид: Propionispira arboris.
Признаки вида: Как грамотрицательные анаэробы, образующие пропионат, организмы рода Propionispira могут быть ошибочно приняты за Propionigeniurrt. Однако Propionispira – это изогнутые подвижные клетки, сбраживающие разнообразные сахара и содержащие цитохромы, тогда как Propionigenium – короткие неподвижные палочковидные клетки, не содержащие цитохромов и сбраживающие только ограниченный набор органических кислот.
Род Rikenella
От редакции: Род Rikenella не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1985 г. (Collins et al,, Syst. Appl. Microbiol. 6: 79–81; Int. J. Syst, Bacteriol, 35: 375, 1985) и включает единственный вид R. microfusus (прежнее название – Bacteroides mic- rofusus).
Мелкие палочки с заостренными концами. Как правило, 0,15–0,3 x 0,3–1,5 мкм, но могут быть до 5 мкм длиной. Неспорообразующие. Неподвижные. Оптимальная температура 37°С. Сбраживают глюкозу и немногие другие сахара, образуя в основном пропионовую и янтарную кислоты. Жирные кислоты в основном метилразветвленные; присутствуют менахиноны. Выделены из фекалий или содержимого слепой кишки телят, кур и японской куропатки.
Типовой (и единственный) вид: Rikenella microfusus.
Признаки вида. Сбраживают глюкозу, маннозу, лактозу, мелибиозу и иногда галактозу. Углеводы стимулируют рост. Штаммы обладают β-гемолитической активностью и способностью разжижать желатину; хорошо растут в присутствии желчи (20%). Продукты брожения – пропионо- вая и янтарная кислоты, но не масляная или молочная.
Род Roseburia
От редакции: Род Roseburia не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1983 г. (Stanton, Savage, Int. J. Syst. Bacteriol. 33: 618–627) и включает единственный вид R. cecicola.
Слегка изогнутые палочки, 0,5 х 2,0–5,0 мкм. Неспорообразующие. Активно подвижные за счет пучка жгутиков, расположенного субтерминально на вогнутой стороне клетки или иногда на одном ее конце. Сбраживают несколько углеводов с образованием бутирата в качестве основного или единственного продукта метаболизма. Выделены из слепой кишки мыши.
Типовой (и единственный) вид: Roseburia cecicola.
Признаки вида: Клетки имеют 20–35 жгутиков в пучке, который при фазово-контрастной микроскопии может выглядеть как один субтерминальный пучок. Хорошо растут на среде, содержащей летучие жирные кислоты, дрожжевой экстракт, триптиказу, минеральные соли, гемин и глюкозу. Кроме бутирата конечные продукты сбраживания глюкозы – СО2, Н2 и в следовых количествах этанол.
Единственный другой род выделяемых из кишечника подвижных грамотрицательных анаэробных организмов в виде изогнутых палочек, образующих большие количества масляной кислоты, – это Butyrivibrio. Однако в отличие от Roseburia клетки Butyrivibrio имеют преимущественно один полярный или субполярный жгутик и грамположительный тип клеточной стенки (хотя клетки окрашиваются по Граму отрицательно). Roseburia можно также принять за Fusobacterium, у которого основной конечный продукт – бутират, но эти организмы различаются по форме клеток, и кроме того, представители Roseburia подвижные, a Fusobacterium нет.
Род Ruminobacter
От редакции: Род Ruminobacter не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1986 г. (Stackebrandt, Hippe, Syst. Appl. Microbiol. 8: 204–207; Int. J. Syst. Bacteriol. 37: 175, 1987) и включает единсгаенный вид R. amylophilus (прежнее название – Bacteroides amylophilus).
Плеоморфные клетки, по форме от овальных до коротких палочек, 0,9–1,2 х 1,0–3,0 мкм, с округлыми или суженными концами, иногда раздутые или неправильной формы. Неспорообразующие. Неподвижные. Оптимальная температура 37–39°С. Метаболизируют углеводы с образованием ацетата, формиата и сукцината. Сфинголипиды и менахиноны отсутствуют; основные клеточные жирные кислоты – неразветвленные, насыщенные и ненасыщенные. Обнаружены в рубце крупного рогатого скота и овец.
Типовой (и единственный) вид: Ruminobacter amylophilus.
Признаки вида. Для роста нуждаются в СО2, NH3 и сбраживаемом углеводе. Образуют кислоту из декстрина, гликогена, крахмала и мальтозы; целлюлозу и целлобиозу не сбраживают.
Род Sebaldella
От редакции: Род Sebaldella не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1986 г. (Collins, Sheh, Int. J. Syst. Bacteriol. 36: 349–350) и включает единственный вид S. termitidis (прежнее название – Bacteroides termitidis).
Палочковцдные клетки, 0,3–0,5 х 2,0–12,0 мкм, со вздутиями в центре, одиночные, в парах и нитях. Неспорообразующие. Неподвижные. Сбраживают сахара с образованием уксусной и молочной кислот и иногда также муравьиной кислоты. Менахиноны отсутствуют; основные клеточные жирные кислоты неразветвленные, насыщенные и ненасыщенные. Обнаружены в содержимом заднего отдела кишечника термитов.
Типовой (и единственный) вид: Sebaldella termitidis.
Признаки вида: Сбраживают глюкозу, фруктозу, мальтозу, маннитол, маннозу, рамнозу, сахарозу, трегалозу и ксилозу. Разлагают мочевую кислоту до СО2, ацетата и аммония. Большинство штаммов образуют H2S. Другие свойства – как для рода.
Род Selenomonas
Изогнутые палочкн, обычно серповидные, 0,9–1,1 х 3,0–6,0 мкм, часто с суженными концами, одиночные, в парах или коротких цепочках. Капсулы отсутствуют, Неспорообразующие. Обладают активной подвижностью кувыркательного типа за счет жгутиков (до 16), расположенных в виде пучка или короткого шнура около центра на вогнутой стороне клетки. Метаболизм бродильного типа. Сбраживают глюкозу с образованием в основном уксусной и пропионовой кислот, СО2 и/или лактата. Встречаются главным образом в ротовой полости человека, в рубце жвачных, слепой кишке свиней н некоторых грызунов; однако один вид выделен из промышленного метантенка, а другой из образца пивных дрожжей.
Типовой вид: Selenomonas sputigena.
Дифференциация видов рода Selenomonas: См. табл. 6.18, Кроме того, для различения видов Selenomonas, выделенных из ротовой полости, полезен следующий ключ.
Таблица 6.18. Дифференцирующие признаки видов рода Selenomonas1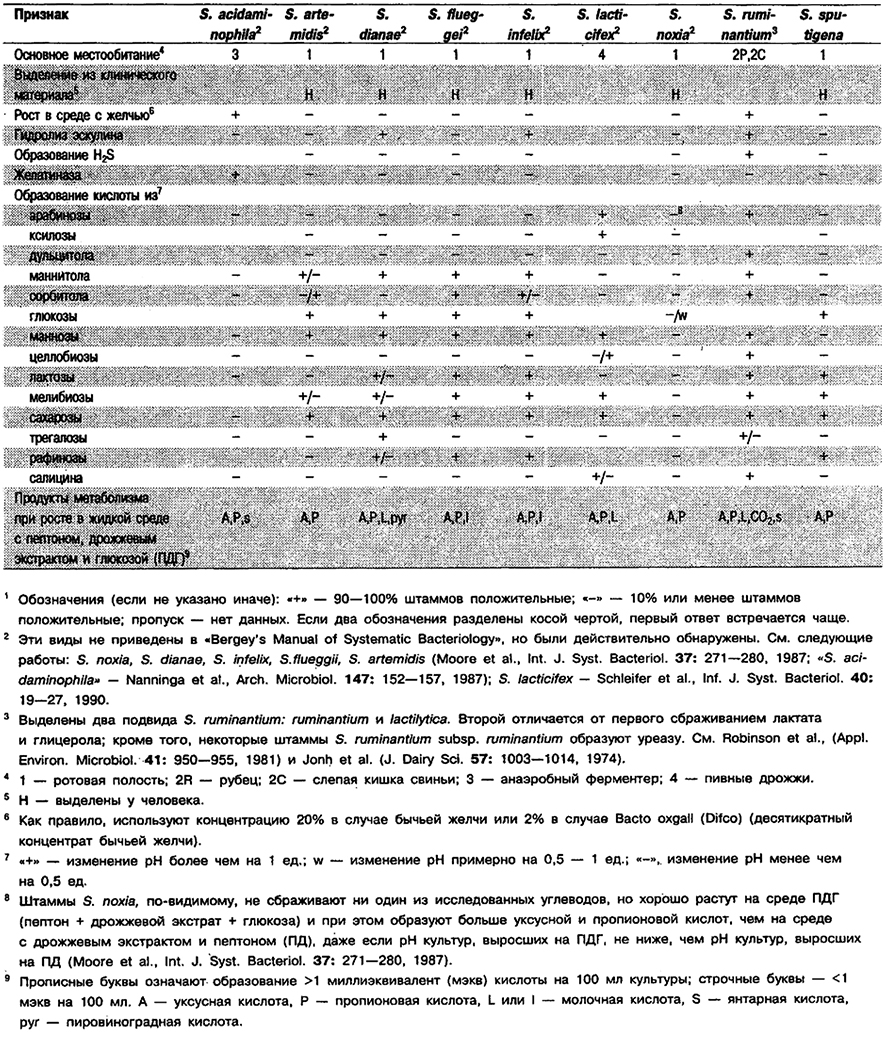
Ключ для определения видов Selenomoitas, выделенных из ротовой полости
I. ОБРАЗУЮТ КИСЛОТУ ИЗ САХАРОЗЫ.
А. Гидролизуют эскулин.
1. Образуют кислоту из трегалозы.
S. dianae
2. Не образуют кислоту из трегалозы.
S. infelix
Б. Не гидролизуют эскулин.
1. Образуют кислоту из лактозы.
а. Образуют кислоту из маннитола,
S. flueggei
б. Не образуют кислоту из маннитола.
S. sputigena
2. Не образуют кислоту из лактозы.
S. artemidis
II. НЕ ОБРАЗУЮТ КИСЛОТУ ИЗ САХАРОЗЫ.
S. noxia
Род Sporomusa
От редакции: Род Sporomusa не приведен в «Вегgey's Manual of Systematic Bacteriology». Этот род описан в 1984 г. (Moller et ah, Arch. Microbiol. 139: 388–396; Int. 3. Syst. Bacterid 35: 224, 1985) в составе двух видов – S. sphaeroides и S. ovata. Последовательно в него добавлены четыре других вида: S. acidovorans (Otliver et al., Arch. Microbiol. 142: 307-310, 1985; Int. J. Syst. Bacteriol. 40: 105, 1990); S. paucivorans (Hermann et al., Int. J. Syst. Bacteriol. 37: 93–101, 1987), S. termitida (Breznak et ai.t Arch. Microbiol. 150: 282–288; Int. J. Syst. Bacteriol. 40: 212, 1990) и S. matonica (Dehning et al., Arch. Microbiol. 151: 421-426, 1989; Int. J. Syst. Bacteriol, 40: 321, 1990).
Изогнутые палочки, 0,5–1,0 x 1–8 мкм, с суженными концами. Большинство штаммов образует округлые или овальные, терминальные или субтерминальные, устойчивые к нагреванию (при 80°С по крайней мере 10 мин) споры. Подвижные за счет жгутиков (до 15), расположенных на вогнутой стороне клетки. Оптимальная температура 30–39°С. Способны использовать в качестве источника энергии некоторые N-метильные соединения» аминокислоты, спирты, гидр оксидированные жирные кислоты или другие органические кислоты, но лишь немногие сахара. Все виды могут использовать Н2 плюс СО2 как субстраты для роста. Основной конечный продукт брожения – ацетат; из некоторых субстратов может дополнительно образовываться бутират или пропионат. Сульфат не восстанавливают. Выделены из речного ила, промышленных отходов и кишечника термитов.
Типовой вид: Sporomusa sphaeroides.
Дополнительное примечание: Морфологически род Sporomusa сходен с родом Selenomonas; оба рода представлены изогнутыми, подвижными, грамот- рицательными палочками с латеральными жгутиками на вогнутой стороне клетки. Однако, кроме того что клетки Selenomonas не образуют споры, они не могут расти с использованием Н2 и СО2 и сильно отличаются по набору субстратов (см. табл. 6.19). По тем же признакам Sporomusa можно отличить от изогнутых, подвижных, грамотри- цательных организмов других родов, таких как Roseburia, Centipeda, Pectinatus и Acetivibrio.
Дифференциация видов рода Sporomusa: См. табл. 6.19.
Таблица 6.19. Дифференцирующие признаки видов рода Sporomusa1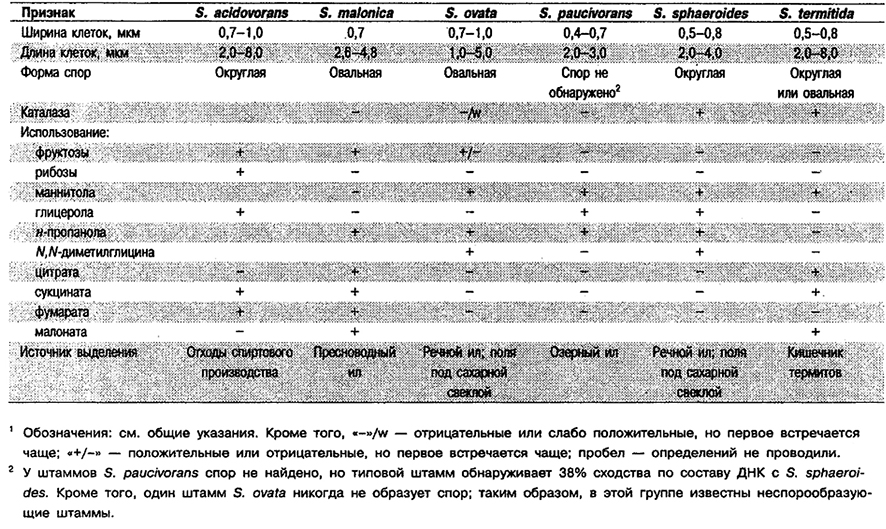
Род Succinimonas
Короткие прямые палочки, 1,0–1,5 х 1,2–3,0 мкм, с округлыми концами. Неспорообразующие, Подвижные за счет единственного полярного жгутика. Оптимальная температура 30–37°С. Сбраживают глюкозу, мальтозу, декстрин или крахмал, но не другие сахара. Образуют большое количество янтарной н меньшее количество уксусной кислот. Обнаружены в рубце крупного рогатого скота, содержавшегося на сено-зерновом рационе.
Типовой (и единственный) вид: Succinimonas аmуlolytica.
Признаки вида: Клетки S. amylolytica – это короткие прямые палочки, иногда почти кокки, в отличие от других грамотрицательных подвижных палочковидных бактерий из рубца, которые имеют изогнутую или спиральную форму. Особенность вида состоит, кроме того, в образовании большого количества янтарной кислоты. Штаммы хорошо растут на синтетической среде, содержащей глюкозу, минеральные соли, аммиак, витамины группы В и буфер СО2/НСО3. Ацетат стимулирует рост; при сбраживании глюкозы происходит потребление СО2.
Род Succinivibrio
Изогнутые или спиральные палочки, 0,4–0,6 х 1,0–7,0 мкм, с заостренными концами. Неспорообразующие. Подвижные за счет единственного полярного жгутика. Сбраживают различные сахара с образованием уксусной н янтарной кислот в качестве основных конечных продуктов и в меньших количествах формиата и лактата. Рост может сопровождаться значительным поглощением СО2. Выделены нз рубца крупного рогатого скота и овец, изредка обнаруживаются при инфекциях у человека.
Типовой (и единственный) вид: Succinivibrio dextrinosolvens.
Признаки вида. Местообитанием служит рубец крупного рогатого скота и овец, чаще при содержании животных на рационах с высоким количеством зерна. У многих штаммов отмечена уреазная активность. Имеются сообщения об обнаружении вида при сепсисе у человека (Porschen, Chan, J. Ciin. Microbiol. 5: 444-447, 1977; Southern, Am. J. Pathol. 64: 540–543, 1975). По многим характеристикам S. dexirinosolvens сходен с Anaerobiospirillum succiniciproducens, однако клетки последнего имеют по пучку жгутиков на каждом полюсе клетки (см. табл. 6.4).
Род Syntrophobacter
От редакции: Род Syntrophobacter не приведен в «Beigey's Manual of Systematic Bacteriology». Этот род описан в 1980 г. (Boone, Bryant, Appl. Environ. Microbiol. 40: 626–632; Int. J. Syst. Bacteriol. 34: 356, 1984) и включает единственный вид S. wolinii.
Палочки н длинные нити, 0,6–1,0 x 1,0–35,0 мкм. Неспорообразующие. Неподвижные. Оптимальная температура 35°С. Растут только в совместной культуре со штаммами Desulfovibrio. В отсутствие организмов, использующих Н2, роста нет. Окисляют пропионат, но не другие жирные кислоты, до ацетата, Н2 и С02- Выделены нз установки для анаэробной очистки сточных вод.
Типовой (и единственный) вид: Syntrophobacter wolinii.
Признаки вида. Свойства те же, что описаны для рода. Отличия от Syntrophomonas spp. см. в табл. 6.20.
Таблица 6.20. Дифференцирующие признаки видов родов Syntrophomonas и Syntrophobacter1
Род Syntrophomonas
От редакции: Род Syntrophomonas не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род был описан в 1981 г. (Mclnemey et al., Appl. Environ. Microbiol. 41: 1024–1039; Int. J. Syst, Bacteriol. 32: 267, 1982) и первоначально включал один вид S. wotfei. Второй вид, S, sapovorans, добавлен в 1986 г. (Roy et al., Arch. Microbiol, 145: 142-147; Int. J. Syst. Bacteriol. 37: 179, 1987).
Слегка изогнутые спиральные палочки, 0,5–1,0 х 2–7 мкм. Неспорообразующие. Подвижные за счет 2–8 жгутиков, расположенных в ряд на вогнутой стороне клетки. В большинстве случаев (при разных условиях) клетки лишь слабо подвижны. Могут содержать гранулы поли-β-гидроксибутирата. Оптимальная температура 30–37°С. Получают энергию за счет β-окисления жирных кислот в совместной культуре с организмами, использующими Н2, такими как Desulfovibrio spp. или Methanospirillum hungatei. Большинство штаммов способно адаптироваться к росту в чистой культуре за счет использования кротоната, хотя растут при этом медленно. Жирные кислоты разлагают преимущественно до ацетата и Н2, но возможно образование пропионата или изовалерата в зависимости от расщепляемой жирной кислоты. Из кротоната образуют ацетат и бутират. Выделены нз бескислородных илов, установок для анаэробной очистки сточных вод и из бычьего рубца.
Типовой вид: Syntrophomonas wolfei.
Дифференциация видов рода Syntrophomonas: См. табл. 6.20.
Род Thermobacteroides
От редакции: Род Thermobacteroides не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1981 г, (Ben-Bassat, Zeikus, Arch, Microbiol. 128: 365–370; Int. J. Syst. Bacterid. 33: 673, 1983); в 1985 г. описание рода исправлено (Ollivier et al., Int. J. Syst. Bacterid. 35: 425–428), Первоначально этот род включал единственный вид Т acetoethylicus, но впоследствии в него были добавлены два других вида – Т. рroteоlyticus (Ollivier et al., Int. J. Syst. Bacteriol. 35: 425–428, 1985), и Т. leptospartum (Toda et al, Agr. Biol. Chem. 52: 1339–1344, 1988; Int. J. Syst. Bacteriol. 39: 93, 1989).
Прямые палочки, 0,5–0,6 x 1,0–6,0 мкм, одиночные или в парах» У некоторых штаммов описаны нитевидные формы. Неспорообразующие. Подвижные за счет перитрихиальных жгутиков или неподвижные. Термофилы; оптимальная температура 60–65°С; диапазон – 35–80°С. Метаболизируют углеводы или пептоны; среди конечных продуктов метаболизма могут быть ацетат, бутират, изобутират, изовалерат, пропиоиат, этанол, СО2 и Н2. Обнаружены в высокотемпературных установках для анаэробной биологической очистки, в компосте из отходов от содержания крупного рогатого скота и в природных термальных местообитаниях, где происходит разложение органических веществ.
Типовой вид: Thermobacteroides acetoethylicus.
Дифференциация видов рода Thermobacteroides: См. табл. 6.21.
Таблица 6.21. Дифференцирующие признаки видов рода Thermobacteroides1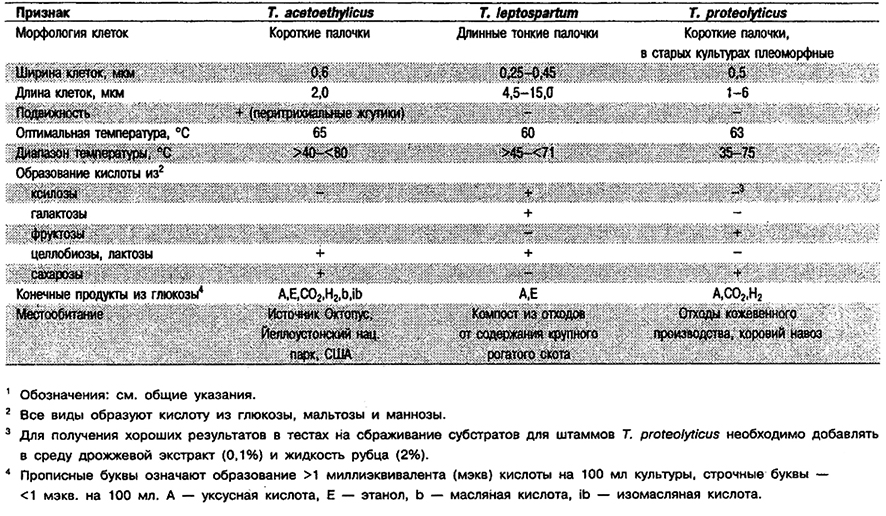
Род Thermosipho
От редакции: Род Thermosipho не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1989 г. (Huber et aJ., Syst. Appl. Microbiol. 12: 32–37; Int. J. Syst. Bacteriol, 39: 496, 1989) и включает единственный вид Т. ajricanus.
Палочки, 0,5 х 3,0–4,0 мкм, окруженные подобной чехлу структурой, за концами клеток раздутой; в одном чехле может быть заключено до 12 клеток. Неподвижные. Термофильные; оптимальная температура 75°С, но могут расти и при температуре 35°С. Не разлагают углеводы; для роста нуждаются в сложных органических субстратах, таких , как дрожжевой экстракт, пептон или триптон. Водород ингибирует рост; добавление Серы снимает ингибирующий эффект; при этом образуется H2S. Растут при концентрации NaCl до 3,6%. Выделены из геотермальных источников в приливно-отливной зоне, Джибути, Африка.
Типовой (и единственный) вид: Thermosipho africanus.
Признаки вида: Признаки, отличающие Т. africanus от других термофильных грамотрицательных, образующих чехлы бактерий, приведены в табл. 6.22.
Таблица 6.22. Дифференцирующие признаки видов рода Thermotoga и Thermosipho africanus 1
Род Thermotoga
От редакции: Род Thermotoga не приведен в «Bergey's Manual of Systematic Bacteriology», Этот род был описан в 1986 г, (Huber et al., Arch. Microbiol 144: 324–333; Int; J, Syst. Bacteriol. 36: 575, 1986) и первоначально включал единственный вид Т maritima. Впоследствии в него добавлены два других вида – Т. neapolitana (Jannasch et al., Arch. Microbiol. 150: 103–104, 1986; Int. J. Syst. Bacteriol. 39: 93, 1989) и Т. thermarum (Windberger et al., Arch. Microbiol. 151: 506–512, 1989; Int. J. Syst. Bacteriol. 42: 327, 1992).
Палочки, 0,6 x 1,5–11,0 мкм, окруженные подобной чехлу структурой, которая выходит в виде вздутий за концы клетки. Неспорообразующие. Могут быть подвижными за счет полярных, латеральных или перитрихиальных жгутиков или неподвижные. Термофильные; оптимальная температура 70–80°С, растут при температуре до 90°С.
Метаболизируют углеводы, триптон или дрожжевой экстракт. Основные продукты брожения – лактат, ацетат, СО2, и Н2. Водород ингибирует рост; у большинства штаммов добавление серы снимает ингибирующий эффект, при этом образуется H2S. Морские штаммы растут при концентрации NaCi до 3,75%. Выделены из геотермально нагретых морских осадков или горячих источников.
Типовой вид: Thermotoga maritima.
Дифференциация видов рода Thermotoga: См. табл. 6.22.
Род Tissierella
От редакции: Род Tissierella не приведен в «Bergey's Manual of Systematic Bacteriology». Этот род описан в 1986 г. (Collins, Shah, Int. J. Syst. Bacteriol. 36: 461–463) и включает единственный вид Т. praeacuta (прежнее название – Bacteroides praeacutus).
Палочки, 0,6–0,9 х 2,0–20,0 мкм, с округлыми или заостренными концами, иногда нитевидные, Неспорообразующие. Подвижные за счет перитри- хнальных жгутиков. Оптимальная температура 37°С; диапазон 25–45°С. Способностью к сбраживанию субстратов не обладают или обладают в слабой степени; конечные продукты при росте в жидкой среде с глюкозой – уксусная, масляная и нзовалернановая кислоты. Менахиноны отсутствуют; жирные кислоты в составе клеток преимущественно метилраэветвленные. Малатдегидрогеназа и глугаматдегидрогеназа отсутствуют, Вьшелены из фекалий взрослых и детей и в некоторых случаях из клинического материала.
Типовой (и единственный) вид: Tissierella praeacuta.
Признаки вида: Как для рода.
Род Wolinella
Спиральные, изогнутые или прямые палочки, 0,5–1,0 х 2–6 мкм, с округлыми или заостренными концами. Неспорообразующие. Подвижные за счет одного полярного жгутика. Хотя виды Wolinella описаны в «Bergey's Manual of Systematic Bacteriology», т. 1, как анаэробы, на самом деле это микроаэрофилы, нуждающиеся в Н2 или формиате и способные к кислородному дыханию. В отсутствие альтернативного конечного акцептора электронов, такого как фумарат, рост происходит с использованием кислорода при его низких концентрациях, а в анаэробных или аэробных условиях отсутствует (относительно W. succinogenes см. Wolin et al., J. Bacteriol. 81: 911–917, 1961; относительно JV. recta см. Ohta, Gottschal, FEMS Microbiol. Ecol. S3: 79-86, 1988 и FEMS Microbiol. Lett. 50: 163–168, 1988; относительно W. curve см, Han et al., Int. J. Syst. Bacteriol. 41: 218–222, 1991). См. также разд. «Группа 2» и «Группа 4» в этом определителе. Оксидазоположителъные; каталазоотрицательные. Содержат цитохромы. В анаэробных условиях рост зависит от присутствия Н2 и фумарата или формиата и фумарата: формиат или Н2 окисляются до СО2, а фумарат восстанавливается до сукдината. Углеводы не используют. Выделены из бычьего рубца, а также у человека из десне вой борозды» корневых каналов зубов при инфекции и клинического материала.
Типовой вид: Wolinella succinogenes.
Дополнительное примечание: Хотя волинеллы не анаэробы, для их выделения пригодны методы, используемые для строгих анаэробов, и из практических соображений они включены в Группу 6.
Дифференциация видов рода Wolinella: См. табл. 6.23.
Таблица 6.23. Дифференцирующие признаки видов рода Wolinella 1,2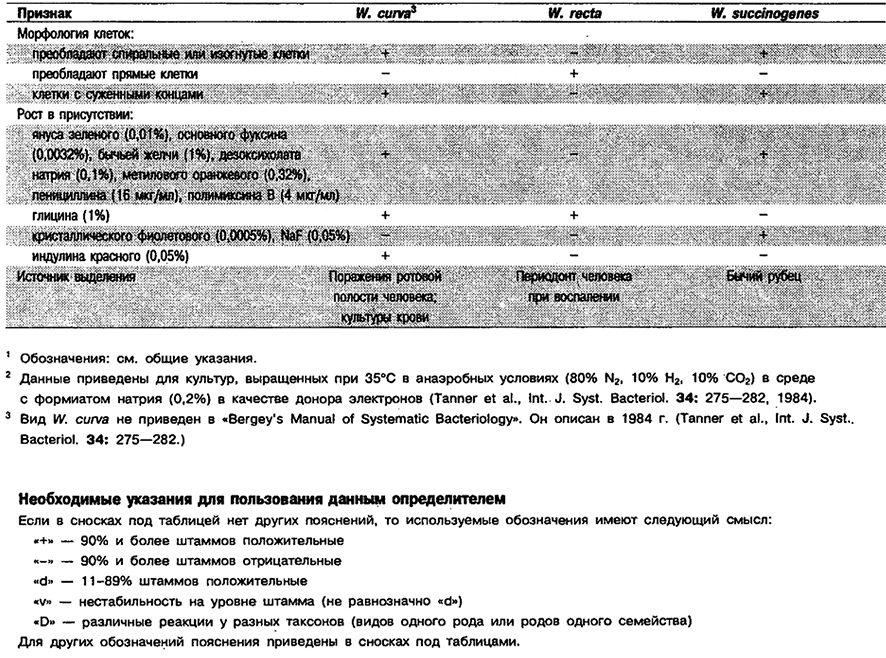
Род Zymophilus
От редакции: Род Zymophilus не приведен в «Bergey's Manual of Systematic Bacteriology». Он описан в 1990 г, (ScWeifer et al., Int. J. Syst. Bacteriol. 40: 19–27) и включает два вида: Z. raffinosivorans и Z. paucivorans.
Палочки, от прямых до слабо изогнутых или спиральных, 0,7–1,0 х 3–30 мкм, одиночные, в парах или коротких цепочках. Подвижные (но подвижность может теряться после нескольких пересевов). Расположение жгутиков не описано. Метаболизм бродильного типа. Глюкозу сбраживают с образованием уксусной и пропноновой кислот и иногда молочной кислоты в следовом количестве. Выделены из задаточных дрожжей и отходов пивоварения.
Типовой вид: Zymophilus raffinosivorans.
Дифференциация видов рода Zymophilus: Z raffinosivorans можно отличить от Z. paucivorans по его способности использовать ксилозу, рафинозу, ино- зитол и мелибиозу. Кроме того, форма клеток у Z. raffinosivorans – от прямой до слегка изогнутой, а у Z. paucivorans – от изогнутой до спиральной.
